| CATEGORII DOCUMENTE |
| Alimentatie nutritie | Asistenta sociala | Cosmetica frumusete | Logopedie | Retete culinare | Sport |
CARACTERIZAREA GENERALÃ A ANTIGENELOR
Conventional, antigenele se definesc ca substante straine, care, consecutiv introducerii in organismul uman sau animal pe o cale parenterala (alta decat cea digestiva), declanseaza sinteza anticorpilor cu care se combina specific. Definitia este incompleta din cateva motive.
1. Calea digestiva de administrare a antigenelor nu exclude totdeauna posibilitatea declansarii raspunsului imun. Pentru agentii infectiosi care se multiplica in tractul digestiv, administrarea orala asigura o buna imunizare (de exemplu, vaccinul polio se administreaza oral, desi calea parenterala este mai eficienta).
2. Unele substante nonself sunt in mod eronat considerate ca neantigenice, deoarece, desi in vivo stimuleaza reactivitatea imunitara si induc sinteza unei cantitati mici de anticorpi, in vitro nu produc reactii vizibile antigen-anticorp.
3. Fata de unele antigene, organismele nu declanseaza raspunsul imun, ci manifesta o stare de toleranta.
4. Unele molecule in stare nativa nu induc un raspuns imun, ci numai dupa cuplarea covalenta cu o molecula purtator. Molecula nativa isi pastreaza proprietatea de a se combina specific cu anticorpii sintetizati. Astfel de molecule se numesc haptene.
J. F. Bach (1976) defineste antigenele ca fiind molecule care, consecutiv introducerii in organism pe o cale adecvata, induc un raspuns imun materializat prin proliferarea celulelor limfoide si sinteza moleculelor de recunoastere (anticorpi si receptori celulari), cu care se combina in vivo si in vitro.
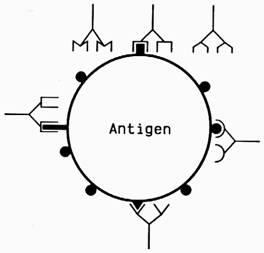
O molecula antigenica este alcatuita din doua componente;
- componenta purtator ("carrier"), care corespunde celei mai mari parti a moleculei:
- gruparile determinante de specificitate sau epitopi, localizate pe suprafata componentei purtator si formate din secvente specifice de monomeri. Epitopii, prin secventa proprie a monomerilor si prin configuratia spatiala specifica, confera individualitate chimica si specificitate antigenica moleculei nonself. Gruparile determinante de specificitate sunt echivalentii moleculari si functionali ai haptenei.
Gruparile determinante de specificitate se gasesc in numar variabil pe suprafata purtatorului si pot fi identice atat in ceea ce priveste compozitia chimica, cat si configuratia spatiala (ca in cazul antigenelor polizaharidice cu epitopi repetitivi) sau sunt diferite, atat ca secventa a monomerilor cat si in privinta configuratiei spatiale.
 Proprietatile definitorii ale antigenelor
Proprietatile definitorii ale antigenelor
In studiile experimentale asupra imunogenitatii unor molecule sintetice, M. Sela (1969) a descris doua proprietati esentiale ale antigenelor:
1. Imunogenitatea sau antigenitatea este proprietatea unui antigen complet, format din gruparea carrier si epitopi, de a declansa un raspuns imun, humoral sau celular, ori de cate ori patrunde in organism pe o cale adecvata. Proprietatea de imunogenitate este asociata cu gruparea carrier a moleculei de antigen, grupare care intr-o oarecare masura influenteaza si specificitatea anticorpilor.
2. Specificitatea defineste capacitatea antigenului intreg sau numai a epitopilor sai de a se combina specific cu anticorpii sau cu receptorii celulari a caror sinteza a fost indusa. Proprietatea de specificitate este dependenta, in primul rand de epitopi, dar este influentata intr-o masura mai mare sau mai mica si de gruparea carrier.
Notiunea de imunogen, uneori, este distincta de aceea de antigen. Notiunea de imunogen este mai restrictiva si semnifica proprietatea unei substante, in stare nativa, de a stimula raspunsul imun, fara sa necesite conjugarea cu o alta molecula.
Notiunea de antigen este mai larga, deoarece desemneaza molecule nonself care sunt imunogene in stare nativa sau devin imunogene dupa conjugarea cu o molecula purtator. Antigenul poate fi uneori inacapabil, in forma sa nativa, sa stimuleze raspunsul imun.
CLASIFICAREA SI IMUNOGENITATEA ANTIGENELOR
Dupa originea lor, antigenele sunt exogene si endogene.
Antigenele exogene sunt cele mai numeroase si pot fi impartite in trei categorii: 1) naturale; 2) artificiale; 3) sintetice.
Antigenele naturale formeaza categoria cea mai cuprinzatoare. Aici sunt incluse toate macromoleculele naturale din virusuri, microorganisme, fungi, plante si animale.
Dupa dimensiuni se disting antigene moleculare ("solubile") si antigene corpusculare.
Antigenele moleculare (solubile) constituie gruparea cea mai numeroasa, care include toate tipurile de macromolecule: proteine, polizaharide, lipide, acizi nucleici.
Antigenele corpusculare ("insolubile") sunt reprezentate de virusuri si de celule (procariote si eucariote).
Cele mai studiate antigene sunt proteinele si polizaharidele, la care se adauga conjugatele: glicoproteine, nucleoproteine, lipoproteine, peptidoglicani, glicolipide.
Proteinele sunt cele mai numeroase si mai importante antigene moleculare. Diversitatea lor chimica, generata de variatia secventei de aminoacizi este uriasa. Practic, fiecare tip de molecula proteica nonself din lumea vie este un antigen pentru organismul animal si uman, deoarece are o secventa unica de aminoacizi, care determina o structura secundara si tridimensionala proprie si implicit, existenta unor epitopi proprii ca secventa a aminoacizilor si conformatie spatiala.
Imunogenitatea este o proprietate generala a proteinelor, a celor cu rol structural (colagenul, cheratina, elastina, fibroina viermelui de matase, proteinele capsidei virale), a celor cu rol functional (miozina, actina, albumina, hemoglobina, mioglobina, enzime, hormoni, imunoglobuline), a celor cu rol de depozit de aminoacizi(ovalbumina, cazeina, gliadina - din semintele de grau). Toate proteinele si polipeptidele cu o greutate moleculara mai mare de 1000 D sunt imunogene, intr-o masura mai mare sau mai mica.
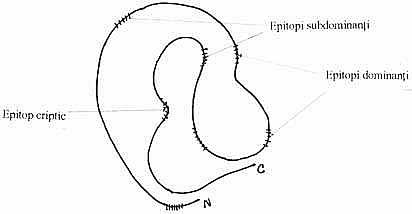
De cele mai multe ori, pentru antigenele proteice, nu se face distinctia dintre epitopii inductori ai raspunsului imun si gruparea carrier, deoarece proteinele poseda un spectru continuum de determinanti antigenici, ce corespund unor secvente discrete ale suprafetei moleculare localizate in zonele cele mai expuse contactului cu receptorii sistemului imunitar. Antigenitatea moleculelor globulare este determinata adeseori, de configuratia lor spatiala, rezultata din plierea tridimensionala. Pentru cele mai multe proteine globulare (mioglobina, hemoglobina, lizozimul, ribonucleaza etc.), aproape toti determinantii antigenici sunt conformationali, adica sunt rezultatul plierii spatiale a moleculei, iar altii sunt secventiali, adica sunt reprezentati de o secventa particulara de aminoacizi. Moleculele proteice fibrilare (cheratina, colagenul, fibroina) au configuratii mai simple decat cele globulare, catenele lor fiind aranjate sau rasucite pe o singura dimensiune. Determinantii antigenici ai acestor proteine sunt secventiali, formati din 3-6 aminoacizi.
Sistemul imunitar al unui organism recunoaste un numar limitat de determinanti antigenici ai unei molecule proteice. Epitopii, conformationali sau secventiali, care stimuleaza raspunsul imun in vivo, iar in vitro induc proliferarea limfocitelor, se numesc epitopi dominanti. O parte a determinantilor antigenici ai unei molecule native, cel mai adesea, sunt neimunogeni (imunosilentiosi), nefiind accesibili sistemului imunitar al organismului, dar se pot exprima intr-un anumit set de conditii de imunizare (gazda, adjuvant etc.). Acestia sunt epitopi interni ai proteinelor globulare. Un determinant intern poate fi silentios in molecula nativa, dar devine imunostimulator dupa clivarea enzimatica a moleculei, in vivo sau in vitro. De aceea, M. Sela a recomandat utilizarea termenului de "grupare imunodominanta", pentru epitopul sau epitopii care se exprima in anumite conditii(gazda, adjuvant, cale de administrare) si determina specificitatea raspunsului imun.
 Imunogenitatea antigenelor proteice se modifica in diferite
conditii.
Imunogenitatea antigenelor proteice se modifica in diferite
conditii.
Denaturarea moleculelor native sub actiunea agentilor chimici si a caldurii, modificarea configuratiei moleculei sub actiunea agentilor reducatori sau hidroliza enzimatica, modifica imunogenitatea. Anticorpii specifici fata de proteina nativa precipita slab sau de loc proteina denaturata termic sau chimic.
Formaldehida si glutaraldehida sunt agenti de legare incrucisata a moleculelor proteice, constituind retele multimoleculare stabile. Acesti agenti produc denaturarea proteinelor si modifica functiile celor cu activitate biologica (toxine, enzime). Astfel, exotoxinele tratate cu formaldehida, isi pierd proprietatile toxice, dar raman imunogene. Formaldehida si glutaraldehida se folosesc pentru conservarea antigenelor cu greutate moleculara mica (peptide), dar sunt mai putin utilizate pentru conservarea proprietatilor antigenice ale moleculelor mari. Agentii chimici de legare incrucisata modifica imunogenitatea moleculelor proteice prin schimbarea conformatiei moleculei si mascarea epitopilor sau prin modificarea chimica a aminoacizilor epitopului.
Denaturarea semnifica deplierea structurii rasucite a moleculelor proteice si are loc prin modificarea pH, prin incalzire, prin reducerea legaturilor S-S sub actiunea ureii si a beta-mercaptoetanolului sau a acidului performic. Prin denaturare, proteina isi pierde nu numai functia biologica, dar isi modifica specificitatea antigenica. De exemplu, cele 4 punti S-S ale RN-azei, intre resturile de cistina, sunt reduse de β-mercaptoetanol si transformate in 8 resturi de cisteina, cu pierderea totala a activitatii enzimatice. Anticorpii fata de RN-aza pancreatica bovina nativa nu precipita moleculele de RN-aza denaturata prin reducerea legaturilor S-S. Invers, anticorpii fata de RN-aza denaturata, nu precipita RN-aza nativa. Modificarea specificitatii anticorpilor sugereaza ca reducerea legaturilor S-S determina pierderea epitopilor conformationali.
Proteinele denaturate reverseaza greu la forma nativa, chiar prin restabilirea conditiilor de mediu.
Hidroliza enzimatica modifica configuratia spatiala a moleculelor proteice native si diminua imunogenitatea lor, cu atat mai mult cu cat fragmentele rezultate au dimensiuni mai mici. Prin clivare enzimatica se anuleaza imunogenitatea epitopilor conformationali si se releva epitopi care in molecula nativa au statutul de epitopi criptici.
O atentie speciala s-a acordat studiului imunogenitatii unor proteine ale caror proprietati biologic-active sunt usor de evaluat: enzime, inhibitori enzimatici, hormoni proteici, toxine, imunoglobuline (in calitatea lor de antigene), proteine ale capsidei sau ale invelisului viral.
Enzimele sunt antigenice, indiferent de originea lor. Reactia moleculelor de enzima cu anticorpii specifici a constituit o modalitate de determinare a pozitiei epitopilor. Anticorpii fata de diferiti epitopi ai moleculei de enzima modifica in grade foarte diferite activitatea ei catalitica. Daca anticorpii sunt specifici fata de epitopi localizati la nivelul situsului activ al enzimei, molecula isi pierde activitatea fata de substrat, deoarece legarea anticorpilor la situsul activ inhiba competitiv legarea moleculelor de substrat. Gradul de inhibitie a activitatii enzimatice este cu atat mai accentuat, cu cat molecula este mai mare. Efectul inhibitor al anticorpilor nu se produce daca enzima a legat deja substratul specific. Daca gruparile determinante de specificitate ale moleculei de enzima sunt situate in afara situsului catalitic, activitatea enzimei este partial inhibata, datorita modificarilor conformationale care survin dupa reactia antigen-anticorp, sau ramane intacta. Foarte rar, complexul enzima-anticorp are un efect catalitic superior, comparativ cu enzima nativa.
Hormonii sunt molecule slabimunogene, datorita uniformitatii relative a structurii lor chimice in regnul animal. Anticorpii specifici fata de majoritatea hormonilor proteici se obtin prin asocierea lor prealabila cu adjuvantul Freund. Imunogenitatea hormonilor este intr-o relatie directa cu gradul deosebirilor chimice existente intre hormonul exogen si hormonul produs de organismul receptor. Consecinta este sinteza anticorpilor antihormon.
Proprietatile
antigenice ale insulinei sunt bine cunoscute, datorita utilizarii
clinice a hormonului. Molecula de insulina este alcatuita
din doua catene polipeptidice, cu un numar total de 51 de aminoacizi:
21 ai catenei A si 30 ai catenei B. Cele doua catene sunt reunite prin
punti S-S. Structura moleculelor de insulina de la diferite specii este
foarte asemanatoare, 47 din cei 51 de aminoacizi fiind identici.
Deosebirile se gasesc in catena A pentru aminoacizii 8, 9 si 10 (la
bovine
Polizaharidele, desi au complexitate structurala relativ mare, conditionata de multitudinea posibilitatilor de legare a atomilor de carbon, sunt molecule slabimunogene in stare nativa, comparativ cu proteinele. Antigenitatea lor este conferita de succesiunea unitatilor componente, de configuratia spatiala a moleculei si de greutatea moleculara. Cele cu greutati mai mici de 50 kD nu sunt imunogene. Polizaharidele sunt antigene cu epitopi secventiali repetitivi si cel putin uneori, in functie de originea polizaharidului si de specia imunizata, sunt imunogeni.
Din punctul de vedere al structurii moleculare, se disting doua tipuri de polizaharide: a) cele care au o catena centrala pe care se insera ramificatiile laterale; b) polizaharide lipsite de o catena centrala, iar ramificatiile sunt dispuse aleatoriu, fara nici o simetrie. Rolul catenelor centrale in conferirea imunogenitatii este controversat, dar ramificatiile laterale au o importanta deosebita pentru determinarea specificitatii antigenice a polizaharidelor. Din punctul de vedere al compozitiei chimice pot fi homo- sau heteroplizaharide, iar in ceea ce priveste sarcina, pot fi neutre sau incarcate. Oligo- si polizaharidele pot dobandi o structura tertiara (globulara). Uneori, configuratia spatiala a polimerului glucidic este determinanta pentru specificitatea sa antigenica. Schimbarile conformationale ale polizaharidelor se produc mai usor decat ale proteinelor, pentru ca au bariere energetice scazute. Pentru polizaharide, denaturarea este practic necunoscuta, ceea ce le confera stabilitate. Daca imunogenitatea polizaharidelor native este slaba, adeseori ele se comporta ca haptene, adica devin antigenice dupa cuplarea cu un purtator proteic, rezultand lectine, cu o foarte larga distributie in lumea vie. In calitate de haptene, polizaharidele au proprietatea de specificitate, adica se combina cu anticorpii complementari fata de complexul glicoproteic.
Din motive de ordin practic, cele mai studiate polizaharide din punct de vedere antigenic sunt cele de origine bacteriana: dextranul si polizaharidele capsulare.
Dextranii sunt polimeri ramificati de glucoza, resturile glucozil fiind unite mai ales prin legaturi de tip α 1-6, dar in functie de specia producatoare, punctele de ramificatie ale catenelor polimere pot fi 1-2, 1-3 sau 1-4.
Dextranii sunt sintetizati in special de unele bacterii lactice, din zaharoza, dupa reactia:
|
Dextranaza |
|
N(zaharoza) ----- ----- ----- (Glucoza)n + (Fructoza)n |
Dextranii au greutati moleculare foarte diferite (pana la 106 D), in functie de gradul de polimerizare. Nu sunt imunogeni si de aceea se folosesc ca inlocuitori ai plasmei. Prin transfuzii repetate cu solutii de dextran la om si prin injectare repetata la soarece, s-au sintetizat anticorpi antidextran. Specificitatea anticorpilor antidextran este foarte inalta. In serul animalelor imunizate cu dextrani s-au detectat doua tipuri de anticorpi: unii specifici fata de resturile de glucozil legate 1-2 si altii specifici fata de resturile de glucozil legate 1-3, ce nu dau reactii incrucisate, desi deosebirea dintre cele doua categorii de molecule de dextran consta numai in modul diferit de legare a resturilor de glucozil intre ele.
Polizaharidele capsulare se pot gasi fie subforma moleculelor libere ("solubile"), fie subforma corpusculara (atasate celulelor bacteriene capsulate).
Variatiile biochimice ale polizaharidelor capsulare, determinate de compozitia glucidica a catenei, de secventa monomerilor sau de modul de legare a lor in catena, confera tulpinilor bacteriene, specificitate antigenica de tip. La Str. pneumoniae s-au identificat peste 80 de tipuri antigenice ale polizaharidelor capsulare. In compozitia lor intra hexoze, pentoze, derivatii lor aminati, metilati etc. Specificitatea antigenica a polizaharidelor capsulare depinde atat de compozitia chimica, cat si de succesiunea monomerilor in catena polizaharidica. Ca vaccinuri, polizaharidele induc starea de toleranta.
In stare purificata sunt molecule neimunogene, datorita uniformitatii lor structurale in lumea vie. Injectarea lor la animale nu induce sinteza anticorpilor. Acizii nucleici nativi sunt conjugate nucleoproteice, in care acizii nucleici au rolul de haptena. Majoritatea epitopilor conjugatului sunt conformationali. O fractie din anticorpii anti-conjugat se combina cu acizii nucleici. Anticorpii anti-acizi nucleici se combina cu acizii nucleici in stare pura, indiferent de provenienta. Proteinele asociate acizilor nucleici confera o noua specificitate antigenica si determina sinteza anticorpilor care se combina cu proteina putator.
Experimental, anticorpii anti-acizi nucleici se obtin pe una din urmatoarele cai:
1. Imunizarea cu bacteriofagi din seria T par (T2, T4, T6), supusi socului osmotic. ADN al acestor fagi se deosebeste de ADN din celulele eucariote, prin prezenta 5-hidroxi-metilcitozinei glicozilate, in locul citozinei. Anticorpii au specificitate fata de bazele glicozilate, ceea ce explica lipsa reactiilor incrucisate cu alti acizi nucleici.
2. Imunizarea cu ribosomi din celulele vegetale sau animale. Anticorpii sintetizati reactioneaza cu ARN de origine bacteriana, vegetala sau animala, precum si cu polinucleotidele sintetice(poli-A, poli-C, poli-U), dar nu reactioneaza cu ADN nativ si nici cu ADN denaturat.
3. Imunizarea cu conjugate haptena-proteina, in care haptena este reprezentata de baze azotate, ribonucleozide, dezoxiribonucleozide, nucleotide, dinucleotide si trinucleotide. Anticorpii sintetizati reactioneaza atat cu haptena cat si cu ADN nativ sau denaturat.
4. Imunizarea cu complexe formate din acizi nucleici si albumina metilata. Cele doua molecule formeaza un complex necovalent, datorita interactiei dintre gruparile negative ale acizilor nucleici si cele pozitive ale proteinei. Anticorpii sintetizati reactioneaza cu complexul molecular, cu proteina, cu acidul nucleic nativ si denaturat de diferite origini.
5. Anticorpii anti-acizi nucleici se gasesc in sangele pacientilor cu lupus eritematos diseminat (LED).
Anticorpii sintetizati fata de acizii nucleici cu rol de haptene in conjugatele cu proteine nu au specificitate, deoarece precipita ADN monocatenar, ARN de diferite origini, poliribonucleotide si acizii nucleici dublu catenari.
Lipidele sunt molecule neimunogene in stare nativa, dar se pot cupla cu proteinele si in conjugatul format au rolul de haptene. Din punct de vedere imunologic, cele mai importante lipide sunt fosfatidele (sfingomielina si cefalina) si glicosfingolipidele(galactocerebrozida).
O haptena lipidica cu o importanta practica deosebita este cardiolipina, din cordul mamiferelor. In sangele indivizilor infectati cu T. pallidum se gasesc anticorpi care reactioneaza cu cardiolipina inalt purificata (reactie incrucisata), extrasa din cordul bovin.
Un alt antigen lipidic este antigenul Forssman, inductor al sintezei anticorpilor hemaglutinanti si in prezenta complementului, hemolitici.
Studiul imunogenitatii lipidelor a fost ingreunat de insolubilitatea lor in apa. Problema reactivitatii anticorpilor cu antigenele lipidice a fost depasita partial, prin utilizarea lipidelor auxiliare (lecitina si colesterolul) in suspensia antigenica. Disponibilitatea liposomilor a permis studiul imunogenitatii lipidelor asociate cu membranele.
Haptenele(haptein, grec = a apuca) sunt substante chimice naturale sau de sinteza, cu molecula mica, a caror imunogenitate este conditionata de cuplarea cu o molecula purtator, dar isi pastreaza proprietatea de specificitate, adica reactioneaza cu anticorpii specifici a caror sinteza a fost indusa de haptena conjugata cu o molecula cu rol de purtator.
Denumirea de "haptena" a fost introdusa de Landsteiner pentru a caracteriza din punct de vedere functional un extract alcoolic de rinichi de cal, neimunogen ca atare pentru iepure, dar capabil sa se combine cu anticorpii sintetizati dupa imunizarea iepurelui cu extractul alcoolic de rinichi de cal, cuplat cu o molecula purtator. El a propus ca in categoria haptenelor sa fie cuprinsa orice substanta naturala sau sintetica, cu greutate moleculara mica sau mare, care in forma nativa nu poate sa induca un raspuns imun detectabil, dar dobandeste capacitatea imunogena, dupa cuplarea sa in vivo sau in vitro, cu o molecula purtator cu greutate moleculara mare. Conjugatul este imunogen nu numai in raport cu epitopii moleculei purtator, ci si cu epitopul haptenei.
Din punct de vedere functional, haptenele s-au numit "jumatati de antigene", deoarece au numai una din cele doua proprietati esentiale ale antigenelor: nu sunt imunogene, dar isi pastreaza proprietatea de specificitate. De aceea, termenul "antigenic" nu este sinonim cu cel de "imunogenic". Haptena este un antigen, dar in forma sa nativa, nu este imunogena.
In general, haptenele sunt molecule mici, desi uneori, macromoleculele pot functiona ca haptene. Extractul alcoolic de rinichi de cal este o haptena complexa. Haptenele simple sunt reprezentate de polinucleotide, alcooli, formaldehida, unele medicamente etc.
Utilizand haptenele simple, prin reactii de cuplare cu o molecula purtator s-au obtinut antigene artificiale. Haptena din complexul molecular are rolul gruparii determinante de specificitate. Studiul imunogenitatii conjugatelor haptena-molecula purtator a permis determinarea marimii gruparilor determinante de specificitate ale antigenelor si indirect, determinarea situsului de combinare a anticorpilor. Pe aceiasi cale s-a evaluat specificitatea, afinitatea si heterogenitatea anticorpilor. Haptenele au fost folosite pentru studiile de cristalografie cu raze X a unui complex antigen-anticorp.
Haptene autocuplante sunt molecule cu greutate moleculara mica, a caror particularitate consta in aceea ca, dupa injectare, in organism se combina spontan cu proteinele tisulare si formeaza conjugate haptena-proteina, in vivo. Conjugatele haptena-proteina induc sinteza anticorpilor si determina procese de hipersensibilitate sau initiaza maladii autoimune. Astfel se comporta derivatii dinitrofenolului substituiti cu clor sau fluor, unii produsi de degradare a penicilinei.
Antigenele corpusculare sunt, in esenta, antigene moleculare asociate virusurilor si celulelor. Proteinele capsidale si glicoproteinele invelisului viral sau proteinele prezentate pe suprafata celulelor infectate cu virusuri, sunt foarte imunogene si stimuleaza raspunsul imun al gazdei. De aceea, imunitatea consecutiva infectiei virale este, de obicei, de lunga durata.
Antigenele bacteriene sunt fie solubile (eliminate in mediul extracelular), fie corpusculare (legate de celula). Din prima categorie fac parte exotoxinele si polizaharidele capsulare libere, iar din cea de a II-a, antigenul somatic O (endotoxina bacteriilor Gram negative), antigenele polizaharidice din glicocalix, flagelina, pilina, acizii teichoici, mureina etc.
Antigenele eritrocitare sunt glicoproteine ale suprafetei eritrocitare, cu determinism biochimic cunoscut, in sistemul A, B, 0. Glicoproteinele eritrocitare de grup sanguin se gasesc si pe suprafata celulelor tisulare, dar si in secretiile exocrine (saliva, suc gastric etc.), la circa 75% dintre indivizi, denumiti "secretori". Glicoproteinele din secretii sunt hidrosolubile si studiul lor a fost mai usor decat al moleculelor eritrocitare.
Gruparile glucidice ale glicoproteinelor membranei eritrocitare au rol dominant in determinarea specificitatii de grup sanguin, asa cum au evidentiat studiile de digestie enzimatica controlata. Eritrocitele tuturor grupelor sanguine au un precursor antigenic comun - antigenul H (codificat de gena H), bine exprimat pe suprafata antigenelor de grup 0 si in cantitati progresiv descrescande pe hematiile de grup A, Bsi Ab.
Specificitatea antigenica de grup 0 este conferita de L-fucoza. Grupul A are o gena ce codifica sinteza glicozil-transferazei, enzima ce adauga N-acetil-D-galactozamina, la galactoza preterminala a moleculei H. De aceea, specificitatea antigenica a eritrocitelor de grup A este conferita de trizaharidul N-acetil-galactozamina, galactoza si L-fucoza:
N-acetil galactozamina (α 1-3) Gal - R (R = restul catenei polizaharidice)
![]() α
1-2
α
1-2
Fuc
Indivizii de grup Bau o gena ce adauga D-galactoza (in loc de N-acetil galactozamina) la galactoza preterminala a moleculei H, avand un determinant antigenic format din doua resturi terminale de D-galactoza si L-fucoza.
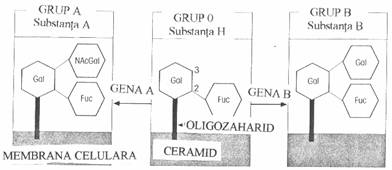 Prezenta restului de fucoza (adica
antigenul H) este esentiala pentru expresia epitopilor A si B. Gena H si
antigenul sau lipsesc la fenotipul
Prezenta restului de fucoza (adica
antigenul H) este esentiala pentru expresia epitopilor A si B. Gena H si
antigenul sau lipsesc la fenotipul
Exprimarea
antigenelor ABO pe hematii poate fi modificata prin tratamentul in vitro
cu glicozidaze: o α-glicozidaza (extrasa din bobul de cafea
verde) poate cliva restul de Gal de pe hematiile de grup B si le converteste in
hematii de grup 0, ce pot ramane functionale dupa transfuzia la
subiectii
de grup 0.
Antigenele de histocompatibilitate (descrise de J. Dausset, 1958) sunt molecule de suprafata ale majoritatii tesuturilor. Din punct de vedere biochimic, ele sunt strict specifice fiecarui organism uman si animal si confera individualitate biochimica proprie fiecarui organism. Se numesc si antigene de transplantare, deoarece, dupa grefarea unui tesut sau a unui organ, moleculele de histocompatibilitate se comporta ca antigene si declanseaza raspunsul imun al organismului receptor, care determina respingerea grefei.
Antigenele individuale de histocompatibilitate se evidentiaza prin reactia de respingere a grefei. In functie de raportul genetic dintre donor si receptor, antigenele de histocompatibilitate apartin urmatoarelor categorii:
1) autoantigenele includ antigenele proprii de histocompatibilitate, care, in conditii normale sunt tolerate de sistemul imunitar. Sub actiunea unor factori fizici, chimici sau biologici, antigenele de histocompatibilitate se modifica devenind autoantigene, generatoare ale conflictelor autoimune;
2) izoantigenele cuprind antigenele de transplantare comune orga-nismelor identice din punct de vedere genetic, care apartin unei linii genetic pure (inbred). Verificarea puritatii genetice a unei populatii de organisme se face prin transplantul de piele. Daca grefa este acceptata, organismele respective apartin aceleiasi linii inbred. Termenii "izoantigen" si "inbred" nu au corespondenta pentru populatia umana;
3) aloantigenele (alos = altul) includ molecule care, dupa injectare declanseaza raspunsul imun la organisme ale aceleiasi specii, dar diferite genetic, de organismul donor. Aloantigenele sunt inegal raspandite la indivizii unei specii si induc raspunsul imun la organismele care nu poseda antigenul respectiv. Aloantigenele se evidentiaza dupa imunizarea unui organism, cu o suspensie celulara provenita de la organisme ale aceleiasi specii, dar apartinand unui alotip diferit;
4) heteroantigenele (xenoantigene, xenos = strain) includ molecule care se gasesc in/pe celulele tuturor indivizilor unei specii si care se comporta ca antigene fata de organismele altei specii. Heteroantigenele se evidentiaza prin sinteza anticorpilor fata de antigenele celulelor provenite de la un organism al unei specii diferite.
Celulele unei specii diferite aduc in organismul receptor nu numai heteroantigene, ci si aloantigene si chiar autoantigene. De aceea injectarea unui heteroantigen este una dintre cele mai utilizate metode pentru a induce sinteza autoanticorpilor.
Antigenele de organ sunt molecule specifice care confera particularitatile biochimice si functionale ale celulelor unui organ. De exemplu, proteinele hepatice sau ale glandei mamare difera de proteinele tesutului renal al aceluiasi organism.
La origine, antigenele artificiale sunt antigene naturale, modificate chimic prin cuplarea, cel mai adesea covalenta, cu una sau mai multe molecule mici, care le confera o noua individualitate antigenica si o noua specificitate de combinare cu anticorpii, in raport cu molecula de origine.
Antigenele artificiale s-au obtinut, in principal, pornind de la moleculele proteice. Prin legarea moleculelor proteice cu diferite haptene s-au obtinut trei tipuri de antigene artificiale:
a) conjugate haptena-proteina, prin reactia de diazotare, iodurare si respectiv substitutie nucleofila;
b) conjugate proteina-proteina, prin intermediul unor agenti bifunc-tionali de legare (diizocianatii si carbodiimidele);
c) proteine legate de suporturi insolubile, prin reactia de diazotare sau prin intermediul carbodiimidelor.
a) Conjugatele haptena-proteina au fost utilizate de Landsteiner, in studiile cu privire la mecanismele raspunsului imun. In conjugate, haptenele indeplinesc rolul de epitopi (grupari determinate de specificitate), iar mole-culele proteice au rolul de carrier. Raspunsul imun nu este orientat strict fata de epitopii haptenici, ci si fata de determinanti antigenici ai gruparii carrier.
Cuplarea haptena-purtator necesita existenta unei grupari reactive a haptenei, care sa se lege covalent cu gruparile functionale ale purtatorului, cu conditia pastrarii integritatii functionale a celor doi reactanti. Haptenele se pot cupla cu purtatori foarte diversi, dar proteinele naturale (albumina, globulinele) furnizeaza conjugate foarte imunogene.
Landsteiner a cuplat amino-benzen-sulfonatul cu molecule proteice, prin reactia de diazotare si a obtinut azoproteine:
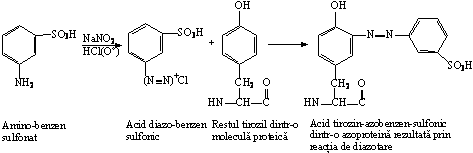
Dupa cuplarea haptenei cu Tir, His sau Lys din structura unei proteine, rezulta un antigen artificial care induce formarea a doua categorii de anticorpi cu specificitati diferite, dupa cum reactioneaza cu haptena, sau cu molecula purtator.
Conjugatele azoproteice au permis studiul influentei configuratiei spatiale a haptenei, asupra specificitatii antigenice. Gruparea sulfonat a fost legata in pozitia orto, meta sau para a haptenei aminobenzen. Antiserurile obtinute au specificitate fata de fiecare izomer. Izomerul meta al aminobenzen-sulfonatului, cuplat cu proteina, pastreaza capacitatea de a precipita cu anticorpii specifici fata de proteina nativa, in timp ce conjugatele cu izomerii orto- si para- dau reactie foarte slaba de precipitare. Concluzia este ca izomerii de pozitie induc modificari sterice (conformationale) ale haptenei.

Conjugatele haptena-proteina se pot obtine prin reactia de iodurare. Proteinele puternic iodurate isi modifica specificitatea antigenica. Ele induc sinteza anticorpilor care dau reactii incrucisate de precipitare cu proteinele iodurate heterologe. Semnificatia este ca prin iodurare, proteinele isi pierd specificitatea antigenica. Toate proteinele iodurate induc sinteza anticorpilor fata de o grupare iodurata, in special fata de tirozina iodurata, indiferent de specificitatea gruparii purtator.
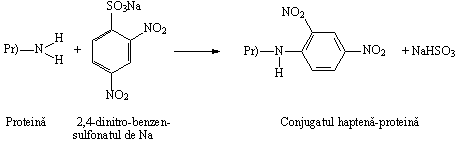
O alta reactie de obtinere a conjugatului haptena-proteina este cea de substitutie nucleofila. Cele mai folosite haptene sunt 2,4-dinitrofenolul(DNP) si 2,4,6-trinitrofenolul(TNP).
Mecanismul
molecular al cuplarii este urmatorul: un atom de H din gruparea
Reactia dintre o proteina si 2,4-dinitrobenzen-sulfonatul de sodiu ilustreaza mecanismul atacului nucleofilic, reactie in care proteina cedeaza electroni, iar nucleul benzenic ii accepta.
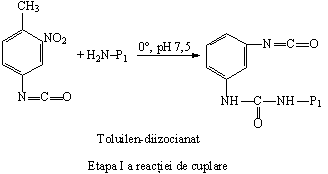
b) Conjugatele proteina-proteina se obtin prin intermediul agentilor bifunctionali de legare: diizocianatii si carbodiimidele. Deoarece gruparile ciano au reactivitate diferita, reactia de cuplare se realizeaza in trepte. De exemplu, gruparea din pozitia 4 a toluilen-diizocianatului este mai reactiva decat gruparea ciano din pozitia 2. Aceasta permite ca una dintre proteine sa se cupleze in pozitia 4, iar ulterior, intr-o noua etapa a reactiei, cea de a II-a proteina se va cupla la gruparea ciano din pozitia 2:
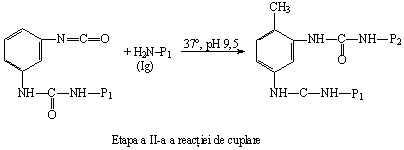
Carbodiimidele, utilizate ca agenti bifunctionali pentru cuplarea proteinelor, sunt considerate anhidride simetrice ale ureii:
|
H2N - C - NH2 -
HOH |
HN = C =NH |
H2N - C 4 N |
|
Uree |
Carbodiimida simetrica |
Cianamida asimetrica |
Carbodiimidele pot fi substituite simetric sau asimetric cu molecule proteice:
R1N = C = NR1 sau R1N = C = NR2.
Agentii bifunctionali de cuplare permit obtinerea conjugatelor proteice (conjugate anticorpi-feritina, insulina-albumina), dar se folosesc si ca mediatori ai legarii diferitelor molecule proteice pe suprafata eritrocitelor.
Marcajul cu feritina este deosebit de important din punct de vedere practic, deoarece se foloseste pentru evidentierea electrono-optica, la nivelul membranei, a diferitelor molecule proteice.
c) Conjugatele proteina-suport insolubil se obtin prin cuplarea proteinelor cu un suport insolubil, prin reactia de diazotare, prin intermediul carbodiimidelor sau al BrCN. Ca suporturi insolubile se folosesc derivati celulozici: sephadex, sepharoza, agaroza etc. Legarea proteinei de suport, prin reactia de diazotare, se face prin intermediul tirozinei, lizinei, histidinei, triptofanului sau argininei.
Conjugatele anticorpi-suport insolubil se numesc imunosorbenti si sunt folositi cu o eficienta deosebita pentru purificarea proteinelor dintr-un amestec, datorita specificitatii lor de combinare cu anticorpii corespunzatori, fixati intr-o coloana de material inert. Antigenul complementar specificitatii de legare a anticorpului fixat in coloana, se leaga necovalent de imunosorbent, dupa care poate fi eluat cu un agent chimic.
Anticorpii pot fi imobilizati pe un suport insolubil, prin tratamentul cu un agent de legare incrucisata (glutaraldehida), dar multe din situsurile reactive pt fi denaturate sau raman ascunse.
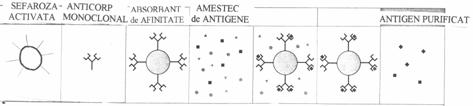 Cel mai bun suport de imobilizare este agaroza, un
polizaharid obtinut prin fractionarea agarului. Agaroza este rezistenta
la actiunea degradativa a enzimelor bacteriene si a agentilor chimici si
poate fi regenerata. Este disponibila subforma sferelor poroase
hidratate, cu diametrul de 40-300 μm si contine 2-8% agaroza in
solutie apoasa.
Cel mai bun suport de imobilizare este agaroza, un
polizaharid obtinut prin fractionarea agarului. Agaroza este rezistenta
la actiunea degradativa a enzimelor bacteriene si a agentilor chimici si
poate fi regenerata. Este disponibila subforma sferelor poroase
hidratate, cu diametrul de 40-300 μm si contine 2-8% agaroza in
solutie apoasa.
In coloana de imunosorbent se pot fixa nu numai anticorpii, ci si antigenele sau chiar celule intacte.
Imunosorbentii se folosesc in activitatea de cercetare si in clinica, pentru prepararea unor produse biologice si a medicamentelor.
Antigenele sintetice sunt polimeri de aminoacizi, cu secventa cunoscuta, obtinuti in vitro. Proprietatile imunogenice ale homopolimerilor (poli-Lys, poli-Glu, poli-Pro) si ale heteropolimerilor au fost studiate de M. Sela. Studiul imunogenitatii heteropolimerilor are avantajul ca ofera posibilitatea studiului influentei compozitiei chimice, a greutatii moleculare si a conformatiei moleculare, usurand studiul imunochimic al gruparilor determinante de specificitate antigenica.
Catenele polipeptidice sintetice pot fi lineare sau ramificate. Cele ramificate rezulta prin atasarea polimerilor lineari, la o catena polifunctionala. Ramificarea se obtine mai usor cu aminoacizi aromatici.
Homopolimerii nu sunt imunogeni, cu exceptia poli-L-Pro, poli-L-Glu, poli-L-Arg, poli-L-Lys. Copolimerii formati din doi aminoacizi nu sunt totdeauna imunogeni, dar cei rezultati prin polimerizarea a trei aminoacizi diferiti sunt totdeauna imunogeni. Cu cat compozitia lor este mai heterogena, imunogenitatea este mai accentuata. Prezenta aminoacizilor aromatici confera o anumita rigiditate a epitopilor si implicit, o imunogenitate superioara.
Pentru a fi imunogeni, copolimerii trebuie sa fie catabolizati de aparatul enzimatic al celulelor care prelucreaza si prezinta antigenul. Polipeptidele formate din D-aminoacizi sunt slab imunogene, datorita incapacitatii organismului de a cataboliza polimerul. Polipeptidele sintetice s-au dovedit a fi foarte utile in studiile de imunochimie, cu privire la:
- determinarea marimii gruparii determinante de specificitate si indirect, a situsului de combinare a anticorpilor specifici;
- rolul dimensiunilor moleculei asupra proprietatilor de imunogenitate;
- rolul configuratiei spatiale a moleculei in conferirea proprietatii de imunogenitate;
- identificarea epitopilor secventiali si conformationali.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 3404
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved