| CATEGORII DOCUMENTE |
| Alimentatie nutritie | Asistenta sociala | Cosmetica frumusete | Logopedie | Retete culinare | Sport |
ORGANIZAREA INFORMATIONALA A ADN: GENELE
1. STRUCTURA SI LOCALIZAREA GENELOR
In prezent, gena este definita ca un complex de secvente nucleotidice necesare pentru a produce o proteina sau o molecula de ARN functional.
Gena ocupa in cromozom o pozitie fixa, intotdeauna aceeasi, numita locus. Locus-ul unei gene poate fi situat pe cromozomi somatici (autozomi) sau pe cromozomii sexuali; fiecare tip de gena ocupa acelasi locus pe cromozomii omologi.
La indivizii unei specii, o gena se poate gasi in forma denumita "salbatica". Gena "salbatica" poate suferi o mutatie, generand o forma alternativa a genei. Aceasta varianta alternativa a genei, care ocupa acelasi locus si influenteaza acelasi caracter se numeste gena alela. Exista situatii in care o gena sufera mai multe mutatii diferite, producand variante alelice numite alele multiple; efectele fenotipice ale alelelor multiple sunt diferite, dar se limiteaza la acelasi caracter. Exemplul clasic il reprezinta alelele multiple A1, A2, B si O pentru locusul care determina grupul sanguin ABO(H).
Gene alele care ocupa loci omologi pot fi identice, definind starea de homozigot, sau diferite - heterozigot. La heterozigoti, alelele diferite se pot manifesta fenotipic diferit. Uneori, una dintre alele este mai puternica si se manifesta fenotipic; gena si caracterul care se manifesta la heterozigoti se numesc dominante. Gena care nu se manifesta fenotipic la heterozigoti, ci se exprima numai in stare homozigota se numeste recesiva.
Termenul de hemizigot se foloseste pentru a defini situatia, diferita de homozigot si heterozigot, in care la barbatii XY o gena de pe cromozomul X nu are echivalent pe cromozomul Y, existand deci o singura gena pentru un caracter.
STRUCTURA GENELOR CARE CODIFICA PROTEINE
Majoritatea genelor umane au o structura discontinua, fiind alcatuite din secvente codante si secvente necodante. Genele care codifica proteine sunt alcatuite dintr-un cadru de citire si din secvente de reglare a genei (fig. 1).
1. Cadrul de citire
Cadrul de citire, aflat in regiunea centrala a genei, reprezinta regiunea transcrisa, sub actiunea ARN-polimerazei, in ARN mesager precursor (ARN premesager). El este alcatuit din secvente codante numite exoni si secvente necodante numite introni.
Exonii sunt secventele codante ale genei, care codifica proteine. Regiunea centrala a genelor incepe si se termina cu exoni, ceea ce inseamna ca orice gena are n exoni si n-1 introni. Numarul exonilor variaza de la o gena la alta, fiind cuprins intre 2 si cateva zeci
Intronii sunt secventele care, in general, nu codifica nici un produs specific si care sunt transcrise initial in ARNm precursor, dar nu se regasesc in ARNm matur. Numarul intronilor variaza de la 2 la 79 la genele distrofinei. Lungimea intronilor este variabila, de obicei mult mai mare ca a exonilor.
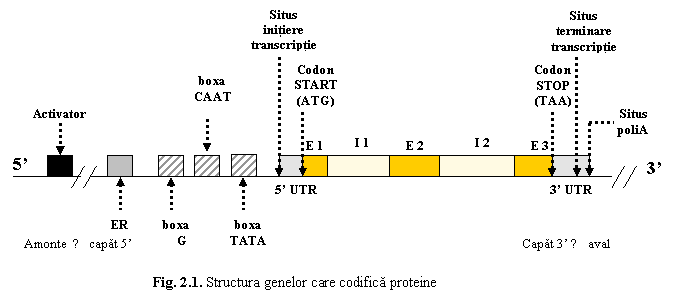 Cadrul
de citire al genei incepe cu situsul de
initiere (start) al transcrierii. Situsul de initiere este urmat de o
regiune netradusa 5'UTR (untranslated region), care contine codonul
start ATG (locul de initiere a translatiei, corespunzator primului aminoacid
din lantul polipeptidic) ; urmeaza primul exon.
Cadrul
de citire al genei incepe cu situsul de
initiere (start) al transcrierii. Situsul de initiere este urmat de o
regiune netradusa 5'UTR (untranslated region), care contine codonul
start ATG (locul de initiere a translatiei, corespunzator primului aminoacid
din lantul polipeptidic) ; urmeaza primul exon.
Dupa ultimul exon din cadrul de citire exista o secventa netradusa 3'UTR, care incepe cu unul dintre codonii stop (TGA, TAG, TAA); codonii stop reprezinta semnalul de oprire a translatiei. Regiunea 3'UTR se continua cu situsul de terminare al transcriptiei. La circa 20 de nucleotide in aval de situsul terminator se afla situsul de poliadenilare, denumit astfel, deoarece la acest nivel la ARNm se adauga circa 200 de nucleotide cu adenozina.
Secventele de reglare a genei
Secventele de reglare a genelor sunt secvente netranscrise, dispuse de o parte si de alta a cadrului de citire, majoritatea la capatul 5' al genei.
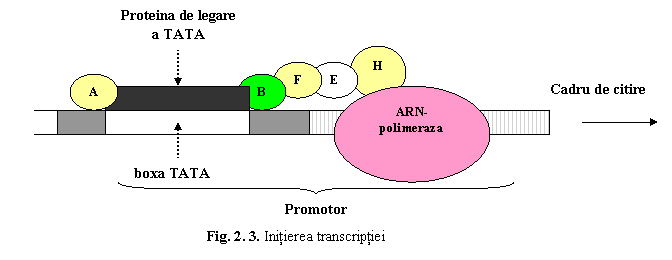
a) Promotorul reprezinta o succesiune de secvente situata in amonte de cadrul de citire. Promotorul contine mai multe secvente nucleotidice scurte, cea mai importanta fiind boxa TATA este situata la 25-35 pb in amonte fata de situsul de initiere a transcrieii si este prezenta la majoritatea genelor. Boxa TATA reprezinta locul la care se fixeaza ARN-polimeraza prin intermediul unor proteine numite factori de transcriptie. Aceasta secventa este necesara pentru initierea corecta a procesului de transcriptie (in punctul de start) (deoarece ARN-polimeraza nu poate recunoaste direct promotorul si nu poate initia singura transcriptia)
b) Intensificatorii (activatorii) (enhancers) sunt secvente ADN care amplifica activitatea promotorului, crescand nivelul transcriptiei. Activatorii sunt situati in amonte fata de promotor.
c) Atenuatorii (silencers) sunt secvente ADN situate, de asemenea, in amonte de promotor si care reduc sau uneori chiar reprima (blocheaza) transcriptia unor gene.
FUNCTIA GENELOR (EXPRESIA INFORMATIEI EREDITARE)
Genele care codifica proteine sunt numite si gene structurale. Genele structurale contin informatia codificata necesara pentru asamblarea intr-o anumita ordine a aminoacizilor in proteina. Relatia "o gena → un polipeptid" este mult mai complexa (de exemplu, exista situatii in care o gena poate produce mai multe polipeptide asemanatoare sau diferite).
MECANISMELE MOLECULARE ALE EXPRESIEI GENICE
 Expresia
informatiei genetice se realizeaza in toate celulele pe baza dogmei centrale a
geneticii:
Expresia
informatiei genetice se realizeaza in toate celulele pe baza dogmei centrale a
geneticii:
Prima etapa a expresiei informatiei genetice este sinteza ARN, cu ajutorul ARN-polimerazei, prin copierea informatiei din ADN. Ea are loc in nucleu si se numeste transcriptie.
A doua etapa este sinteza unui polipeptid, prin decodificarea (traducerea) informatiei din ARNm intr-o secventa specifica de aminoacizi. Procesul are loc in citoplasma, pe ribozomi, si se numeste translatie.
1. TRANSCRIPTIA
Transcriptia este procesul prin care informatia genetica a unei gene este copiata intr-o molecula de ARN complementara si antiparalela, numita ARN mesager (ARNm). Acest proces are loc in nucleu.
Acidul ribonucleic (ARN) prezinta o serie de particularitati structurale.
ARN este alcatuit din ribonucleotide. Fiecare ribonucleotid este format din riboza (o pentoza care prezinta cate o grupare OH in pozitiile 2' si 3'), o baza azotata (adenina - A, guanina - G, citozina - C sau uracil - U) si un grup fosfat. Prin polimerizarea in sensul 5'→3' a ribonucleotidelor rezulta o monocatena, de obicei scurta, de ARN.
Se deosebesc mai multe tipuri de ARN, cu functii diferite:
ARNm sau mesager este codant, deoarece prin translatie determina sinteza unui polipeptid;
ARNt sau de transfer are rolul de a transporta aminoacizii la ribozomi;
ARNsn sau ARN mic nuclear intervine in procesarea intronilor.
In cele ce urmeaza, este prezentata sinteza ARNm catalizata de ARN-polimeraza II.
(1) Ribonucleotide activate. Prin ruperea legaturilor fosfat macroergice dintre fosfatii α si β ai ribonucleozid-trifosfatilor (adenozintrifosfat - ATP, guanozintrifosfat - GTP, citidintrifosfat - CTP si uridintrifosfat - UTP) se elibereaza ribonucleotidele (ribonucleozid-monofosfatii) si energia necesara polimerizarii acestora.
(2) Matrita este reprezentata de o singura catena de ADN. Numai una dintre cele doua catene ale ADN (3'→5') este transcrisa si serveste ca matrita pentru sinteza moleculei complementare si antiparalele de ARNm (5'→3'); catena transcrisa este denumita si catena antisens. Catena ADN netranscrisa, ale carei secvente nucleotidice sunt identice cu cele ale ARNm, este numita si catena sens (fig. 2). Cele doua catene ale ADN se vor separa temporar si pe lungimi scurte, pentru ca una din ele sa serveasca drept matrita pentru ribonucleotidele complementare.
5'.ATGTTACGACGT.3' catena ADN netranscrisa (catena sens)
![]() 3'.TACAATGCTGCA.5' catena
ADN transcrisa (catena antisens)
3'.TACAATGCTGCA.5' catena
ADN transcrisa (catena antisens)
Transcriptie
5'.AUGUUACGACGU.3' ARNm
Fig. 2.2. Catenele ADN
(3) Enzima care catalizeaza tanscrierea ADN in ARNm este ARN-polimeraza II.
Procesul transcriptiei se desfasoara in doua mari etape:
formarea ARN mesager precursor;
procesarea (maturarea) ARN mesager precursor.
a. Formarea ARNm precursor
 ARNm precursor se
formeaza in trei etape: initiere, elongare si terminare.
ARNm precursor se
formeaza in trei etape: initiere, elongare si terminare.
1) Initierea transcriptiei. Deoarece ARN-polimeraza nu poate recunoaste direct promotorul si nu poate initia singura transcriptia, sunt necesari o serie de factori de transcriptie, care se fixeaza in regiunea promotor (fig. 3), dirijand si activand astfel enzima. Transcriptia incepe la nivelul situsului de initiere a transcriptiei.
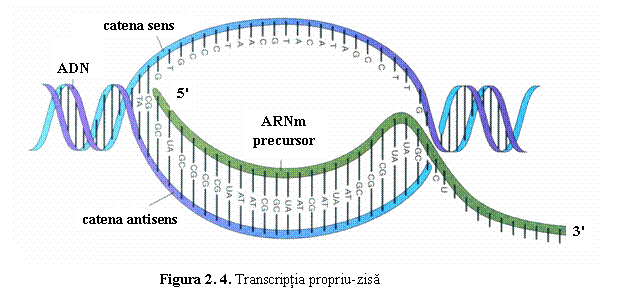 (2)
Transcriptia propriu-zisa (elongarea). Dupa fixarea ARN-polimerazei pe regiunea promotor, cele doua catene ale
ADN sunt separate, eliberandu-se catena antisens; catena antisens serveste ca
matrita pentru atasarea complementara, in
sensul 5'- 3', a ribonucleotidelor activate (fig. 4).
(2)
Transcriptia propriu-zisa (elongarea). Dupa fixarea ARN-polimerazei pe regiunea promotor, cele doua catene ale
ADN sunt separate, eliberandu-se catena antisens; catena antisens serveste ca
matrita pentru atasarea complementara, in
sensul 5'- 3', a ribonucleotidelor activate (fig. 4).
(3) Terminarea transcriptiei se produce atunci cand ARN-polimeraza intalneste situsul de terminare a transcriptiei. ARN-polimeraza si factorii de transcriptie se detaseaza de matrita ADN si se elibereaza molecula de ARNm precursor care contine atat exonii, cat si intronii genei copiate.
O gena poate fi transcrisa simultan de mai multe molecule de ARN-polimeraza, formandu-se astfel mai multe copii de ARNm ce contin aceeasi informatie genetica.
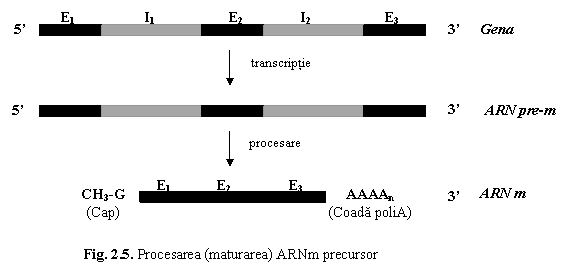
b. Procesarea (maturarea) ARNm precursor
Maturarea ARNm precursor (fig. 5) are loc la nivel nuclear si se realizeaza prin doua procese: (1) insertia de secvente nucleotidice la extremitatile moleculei si (2) matisarea ARN precursor.
(1) Insertia de secvente nucleotidice. La primul nucleotid al capatului 5' al ARN premesager se adauga o molecula de metil-guanozina, formand o structura denumita boneta (cap). La capatul 3' se adauga intre 50-200 nucleotide cu adenina, formand o coada poliadenilica.
(2) Matisarea ARN precursor (splicing) consta in decuparea si eliminarea intronilor, urmata de reunirea (legarea "cap la cap") a exonilor, cu formarea ARNm matur.
Procesul de matisare este mediat de un complex format din ARN nuclear mic (ARNsn) si proteine.
In cadrul procesului de matisare exonii sunt sudati, de cele mai multe ori, in ordinea in care acestia sunt dispusi in interiorul genelor - matisare constitutiva (constitutive splicing). La numeroase gene umane, din ARNm precursor sunt eliminati atat intronii, cat si o parte din exoni; moleculele de ARN matur contin doar anumiti exoni care pastreaza ordinea in care sunt dispusi in gena; prin acest proces de matisare alternativa (alternative splicing), dintr-o molecula de ARNm precursor se formeaza molecule diferite de ARNm matur, o gena putand determina, astfel, sinteza mai multor proteine diferite: de exemplu, gena calcitoninei produce in celulele C din glanda tiroida calcitonina (hormon cu rol in reglarea metabolismului fosfo-calcic), iar in hipotalamus un peptid inrudit cu calcitonina (calcitonin-related peptide) (cu functii neuromodulatoare si trofice).
TRANSLATIA
Translatia (traducerea) reprezinta a doua etapa a expresiei genice si consta in decodificarea informatiei genetice din ARNm care permite aranjarea specifica a aminoacizilor si polimerizarea lor intr-un lant polipeptidic
Procesul de translatie necesita un "dictionar" reprezentat de codul genetic si un aparat de translatie, care cuprinde un "traducator", reprezentat de moleculele de ARN de transfer (ARNt), ribozomi, enzime si diferiti cofactori.
|
Primul nucleotid (capatul 5') ↓ |
Al 2-lea nucleotid |
Al 3-lea nucleotid (capatul 3') ↓ |
|||
|
U |
C |
A |
G |
||
|
U |
Phe |
Ser |
Tyr |
Cys |
U |
|
Phe |
Ser |
Tyr |
Cys |
C |
|
|
Leu |
Ser |
STOP |
STOP |
A |
|
|
Leu |
Ser |
STOP |
Trp |
G |
|
|
C |
Leu |
Pro |
His |
Arg |
U |
|
Leu |
Pro |
His |
Arg |
C |
|
|
Leu |
Pro |
Gln |
Arg |
A |
|
|
Leu |
Pro |
Gln |
Arg |
G |
|
|
A |
Ile |
Thr |
Asn |
Ser |
U |
|
Ile |
Thr |
Asn |
Ser |
C |
|
|
Ile |
Thr |
|
Arg |
A |
|
|
Met |
Thr |
|
Arg |
G |
|
|
G |
Val |
|
Asp |
Gly |
U |
|
Val |
|
Asp |
Gly |
C |
|
|
Val |
|
Glu |
Gly |
A |
|
|
Val |
|
Glu |
Gly |
G |
|
1.) Codul genetic
Conform dogmei centrale a geneticii moleculare informatia din structura unei gene, stocata ("scrisa") sub forma unei anumite secvente a celor patru tipuri de nucleotide (A, T, G, C), este copiata (transcrisa) complementar (U, A, C, G) in ARNm si apoi tradusa intr-o anumita secventa de aminoacizi, care vor forma un lant polipeptidic. Translatia este efectuata cu ajutorul codului genetic.
Codul genetic reprezinta relatia de corespondenta intre o anumita secventa de trei nucleotide numita codon si un anumit aminoacid. Cele patru litere (nucleotide) pot forma cuvinte alcatuie din trei litere, iar gena este un "mesaj coerent" format dintr-o insiruire de codoni (tabel 1).
Tabel 2. 1. Codul genetic
Caracteristicile codului genetic.
(1) "Cuvintele" codului sunt reprezentate de grupuri de cate trei nucleotide, numite codoni. Codul genetic este triplet. Combinatiile celor patru baze luate cate trei (43) realizeaza 64 de codoni, dintre care 61 codifica un aminoacid (codoni sens).
(2) Codul genetic are un codon start (AUG) si trei codoni stop (UAA, UAG, UGA) (codoni nonsens).
(3) Deoarece exista 61 de codoni sens si numai 20 de aminoacizi, inseamna ca unii aminoacizi pot fi specificati de mai multi codoni; codonii diferiti ce codifica acelasi aminoacid se numesc codoni sinonimi (diferiti, de obicei, prin a treia baza), iar codul genetic este degenerat.
Caracterul degenerat al codului genetic constituie un avantaj pentru celula, oferind protectie impotriva efectelor mutatiilor (mutatiile genice care produc codoni sinonimi nu afecteaza structura proteinei codificate).
(4) Codul genetic este nesuperpozabil: codonii vecini nu au nici un nucleotid comun.
(5) Codul genetic prezinta continuitate (fara pauze): intre codonii adiacenti nu exista nucleotide cu rol de "virgula".
(6) Codul genetic este lipsit de ambiguitate: un codon specifica intotdeauna un aminoacid, totdeauna acelasi.
(7) Codul genetic este universal, acelasi la toate organismele (bacterii, plante, animale si om). Cu toate acestea, codul genetic mitocondrial are cateva mici diferente fata de cel nuclear.
) Aparatul de translatie
Aparatul de translatie este alcatuit din ARNm, ARNt, ribozomi, aminoacizi, enzime, etc.
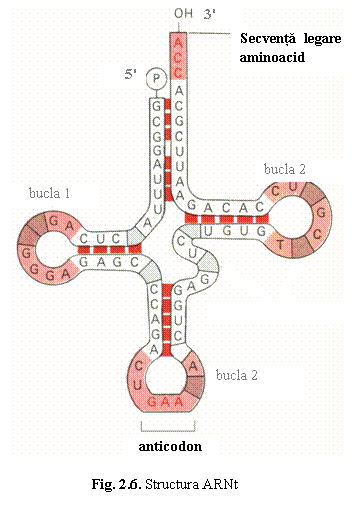 a) ARN de transfer (ARNt) transporta aminoacizii din citoplasma la ribozomi,
sediul sintezei proteice, unde ii pozitioneaza in ordinea dictata de codonii
din ARNm; ARNt are deci rolul de "translator".
a) ARN de transfer (ARNt) transporta aminoacizii din citoplasma la ribozomi,
sediul sintezei proteice, unde ii pozitioneaza in ordinea dictata de codonii
din ARNm; ARNt are deci rolul de "translator".
ARNt este un poliribonucleotid cu structura "in trifoi": prezinta trei regiuni scurte, dublu catenare numite tulpini, care se termina cu bucle monocatenare (fig. 6), structura care este identica la toate tipurile de ARNt. ARNt contine doua secvente importante: (1) secventa CCA de la capatul 3'OH de care se leaga aminoacidul cu ajutorul enzimei aminoacil-ARNt-sintetaza; (2) anticodonul, o secventa de trei nucleotide situata in bucla opusa capatului 3'; anticodonul recunoaste, prin complementaritate, codonul din ARNm corespunzator aminoacidului.
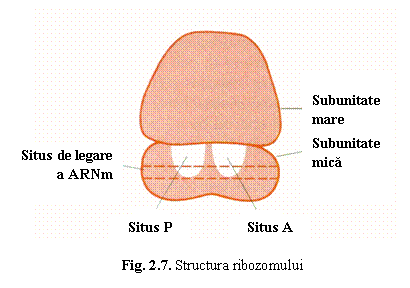 (b) Ribozomii sunt organite
citoplasmatice in care are loc asamblarea aminoacizilor in proteine pe baza
informatiei din ARNm. Sunt alcatuiti din doua subunitati deosebite prin
constanta de sedimentare (S). In perioada in care nu sunt implicate in sinteza
proteica, cele doua subunitati sunt disociate si libere in citoplasma; ele se
asociaza la inceputul translatiei formand ribozomul activ. Fiecare subunitate
este alcatuita din ARN ribozomal (ARNr) si proteine. Subunitatea mica (40S) prezinta un situs de legare a ARNm. Subunitatea mare (60S) prezinta doua
situsuri invecinate: situsul A
(aminoacil) si situsul P (peptidil)
unde se fixeaza moleculele de ARNt cuplate cu aminoacizi (fig. 7).
(b) Ribozomii sunt organite
citoplasmatice in care are loc asamblarea aminoacizilor in proteine pe baza
informatiei din ARNm. Sunt alcatuiti din doua subunitati deosebite prin
constanta de sedimentare (S). In perioada in care nu sunt implicate in sinteza
proteica, cele doua subunitati sunt disociate si libere in citoplasma; ele se
asociaza la inceputul translatiei formand ribozomul activ. Fiecare subunitate
este alcatuita din ARN ribozomal (ARNr) si proteine. Subunitatea mica (40S) prezinta un situs de legare a ARNm. Subunitatea mare (60S) prezinta doua
situsuri invecinate: situsul A
(aminoacil) si situsul P (peptidil)
unde se fixeaza moleculele de ARNt cuplate cu aminoacizi (fig. 7).
(c) Enzimele implicate in procesul translatiei sunt: aminoacil-ARNt-sintetaza de care se leaga aminoacidul si ARNt corespunzator; peptidil-transferaza catalizeaza formarea de legaturi peptidice intre aminoacizi.
Translatia se desfasoara in trei faze succesive: initierea, elongarea si terminarea translatiei.
(1) Initierea translatiei este etapa in care primul aminoacid al proteinei este pozitionat in dreptul codonului start AUG din ARNm (ce corespunde metioninei) si sunt asamblate cele doua subunitati.
Initial, ARNm se fixeaza cu "boneta" (cap) pe subunitatea mica (40S) a ribozomului;
apoi ARNm si primul ARNt "incarcat" cu primul aminoacid metionina formeaza complexul
de initiere; anticodonul ARNt-1 este in contact cu codonul AUG al ARNm.
Urmeaza fixarea subunitatii mari (60S), iar ribozomul devine activ. ARNt-1
ocupa situsul P al subunitatii 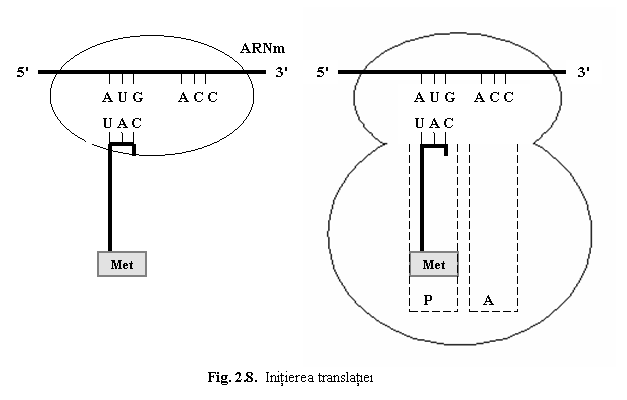
 mari,
situsul A fiind liber (fig. 8).
mari,
situsul A fiind liber (fig. 8).
(2) Elongarea este etapa in cursul careia se formeaza legaturi peptidice intre aminoacizii aranjati pe baza ordinii codonilor din ARNm.
Dupa pozionarea ARNt initiator in situsul P al ribozomului, pe situsul A liber se plaseaza al doilea ARNt incarcat cu aminoacidul codificat de al doilea codon din ARNm. Sub actiunea peptidil-transferazei, se formeaza o legatura peptidica intre primii doi aminoacizi; rezulta astfel un dipeptid care este legat la cel de-al doilea ARNt. Apoi, ribozomul se deplaseaza cu trei nucleotide in lungul ARNm, in directia 5'→3'. Astfel, ARNt-1 paraseste situsul P, care va fi ocupat de cel de-al doilea ARNt ce contine dipeptidul. In situsul A eliberat se fixeaza a treilea ARNt cu aminoacidul corespunzator, iar acest al treilea aminoacid se leaga de dipeptid (fig. 9).
Cele trei faze - atasarea aminoacid-ARNt la ribozom, formarea legaturii peptidice si translocarea ribozomului - se repeta ciclic, determinand cresterea lantului polipeptidic.
Pe masura ce un ribozom avanseaza in lungul moleculei de ARNm, si alti ribozomi pot incepe citirea moleculei, formandu-se astfel sute sau mii de polipeptide identice.
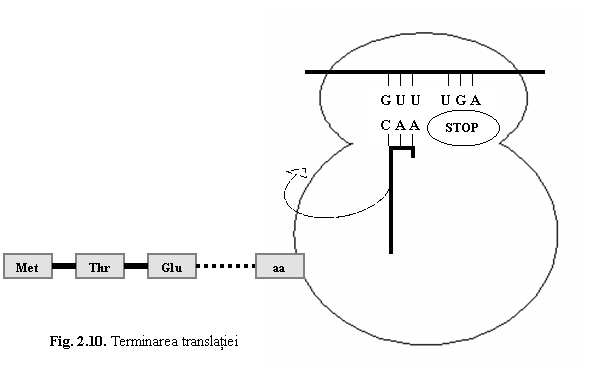
(3) Terminarea traducerii are loc in momentul cand in situsul A ajunge un codon stop (UAA, UAG sau UGA) (fig. 10). Deoarece nu exista un ARNt corespunzator codonilor stop, se leaga o molecula de apa la nivelul peptidului. Polipeptidul eliberat astfel din ribozom va suferi o serie de modificari si va dobandi forma tridimensionala specifica.
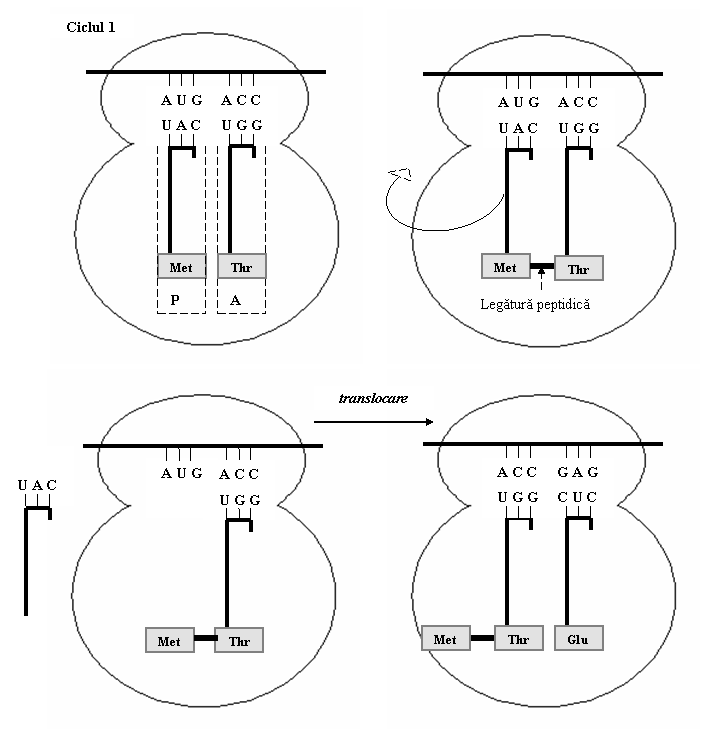
Fig. 2.9. Translatie - elongare
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 4743
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved