| CATEGORII DOCUMENTE |
| Alimentatie nutritie | Asistenta sociala | Cosmetica frumusete | Logopedie | Retete culinare | Sport |
Tesutul muscular
Tesutul muscular este adaptat functiei de contractie. Celula (fibra musculara) prezinta unul sau mai multi nuclei (dupa tipul de tesut muscular), o membrana celulara, numita sarcolema, si o citoplasma, numita sarcoplasma, in interiorul careia se gasesc organitele celulare comune si organitele specifice (contractile) - miofibrilele - aparute in urma diferentierii si adaptarii celulei la functia de contractie.
Muschii reprezinta 30-40% din greutatea corporala, adica aproximativ 30 kg pentru un om de 70 kg.
Dupa particularitatile miofibrilelor, tesuturile musculare se impart in trei tipuri: tesut muscular striat, cu miofibrile heterogene, de aspect striat, care se contracta voluntar, tesut muscular cardiac, in care miofibrilele sunt striate, dar tesutul se contracta involuntar; tesut muscular neted, in care miofibrilele sunt omogene si se contracta involuntar.
1. Tesutul muscular striat scheletic
Tesutul muscular striat este alcatuit din fibre care intra in constitutia muschilor scheletici (40% din greutatea corpului), iar, la nivelul viscerelor, le intalnim atat in musculatura limbii, faringelui, a portiunii superioare a esofagului si in cea a unor sfinctere (anal extern si cel extern al uretrei), cat si in muschii extrinseci ai globului ocular si diafragma. Fibra musculara striata are o forma cilindrica sau prismatica, cu extremitatile rotunjite sau ramificate (muschii fetei si ai limbii).

Tesutul muscular striat scheletic 1. striatie; 2. nucleu
Fibrele sunt elemente multinucleate, plasmoidale, cu zeci sau sute de nuclei de forma ovoida situati periferic, imediat sub sarcolema. Lungimea fibrei este cuprinsa intre 3 si 12 cm, iar grosimea este de 20-100 m.
Sarcolema, examinata la microscopul electronic, prezinta doua portiuni distincte: sarcolema propriu-zisa si membrana bazala. Sarcolema propriu-zisa are rolul de a propaga excitatia de-a lungul fibrei musculare si are structura generala a plasmalemei. Membrana bazala, formata din proteine colagenice, are rolul de a mentine forma fibrei musculare in limite normale.
Sarcoplasma contine organite comune, diferite incluziuni si numeroase miofibrile. Reticulul sarco-plasmatic este foarte dezvoltat, avand un rol important in contractie; datorita prezentei unei mari cantitati de ioni de calciu, la acest nivel se realizeaza cuplarea excitatiei cu contractia.
Miofibrilele sunt constituite din numeroase fibrile elementare, numite miofilamente, care constituie unitatea ultrastructurala si functionala a miofibrilei. Miofilamentele sunt de doua tipuri: groase, formate din miozina, si subtiri, formate din actina, tropomiozina si troponina. Cele doua tipuri de miofilamente au o asezare precisa, fiind dispuse intr-un aranjament hexagonal, cu un miofilament gros in centru, inconjurat de sase miofilamente subtiri.
Fibrele musculare striate se grupeaza in fascicule de 20-30 de fibre, numite fascicule primare, inconjurate de o teaca conjunctiva numita endomisium. Fasciculele primare (4-5) se grupeaza in fascicule secundare, delimitate de tesut conjunctiv cu fibre elastice, numit perimisium. Acestea se grupeaza in fascicule tertiare, cuaternare, care formeaza, de fapt, muschiul, iar acesta este invelit de epimisium.
Vascularizatia este foarte bogata, asigurand procesele metabolice intense din timpul contractiei.
Muschii striati au o inervatie motorie (placa motorie) si senzitiva (fibrele aferente plecate de la proprioceptori musculari).
2 Tesutul striat de tip cardiac (miocardul)
Miocardul este constituit din fibre musculare cu o structura asemanatoare fibrelor musculare striate (miofibrilele prezinta alternanta de benzi clare si intunecate) si fibrelor musculare netede, prin pozitia centrala a nucleului. Celulele musculare individualizate, alungite si ramificate, vin in contact unele cu altele la nivelul unor benzi numite discuri intercalare (striile scalariforme), care reprezinta jonctiuni intercelulare specializate.

Tesutul striat de tip cardiac 1. striatie; 2. nucleu; 3. disc intercalar
Fibra cardiaca are un diametru si o lungime mai mici decat fibra striata, iar capetele sunt de obicei ramificate. Mitocondriile sunt mai numeroase decat in fibrele striate si netede, fiind asezate intre miofibrile sub forma unor coloane (datorita activitatii contractile permanente). Miofibrilele prezinta aceleasi caractere morfologice si structurale ca la fibra striata de tip scheletic, fiind insa mai groase.
Pe langa acest miocard de tip contractil exista si tesutul nodal (miocardul specific), care determina contractia ritmica si automata a inimii, stabilind o legatura anatomica si functionala intre atrii si ventricule (nodului sinoatrial, atrioventricular, fasciculul Hiss si reteaua Purkinje).
3. Tesutul muscular neted
Fibra musculara neteda este unitatea morfo-functionala a tesutului muscular neted. Ea intra in constitutia tunicii musculare a viscerelor, a vaselor, in anexele unor organe de simt (piele), in capsulele unor organe (splina, suprarenala). Fibrele sunt asezate in straturi, benzi sau raspandite izolat in tesutul conjunctiv. Fibrele sunt paralele intre ele, iar portiunea ingrosata a unei fibre vine in raport cu extremitatile efilate ale fibrelor invecinate.
Fibra musculara neteda, de aspect fusiform, are o lungime cuprinsa intre 10-100 u- si cu 0 de 2-4 u. Este formata din sarcolema, sarcoplasma, care contine organitele comune, incluziuni celulare si organitele specifice - miofibrilele -, si un nucleu central, de forma alungita.
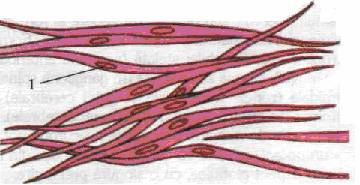
Tesut muscular neted 1. nucleu
Miofibrilele au o forma alungita si se intind de la un capat la altul al fibrei. Totodata, prezinta o structura complexa, fiind alcatuite din miofilamente de 10-150 Å, sunt omogene (fara striatii transversale), iar, din punct de vedere biochimic, sunt formate din proteine contractile (actina, miozina) si reglatoare (tropomiozina si troponina).
Inervatia este asigurata de sistemul nervos vegetativ simpatic si parasimpatic.
Pentru a sustine o tensiune de contractie egala cu a muschiului scheletic, muschiul neted necesita numai 1/10 pana la 1/300 din energia consumata de cel striat.
Dupa ce a fost stimulat, un muschi neted tipic incepe sa se contracte la 50-100 ms si atinge maximum de contractie o jumatate de secunda mai tarziu, dupa care, in una pana la doua secunde, incepe declinul fortei de contractie. Timpul total de contractie este de 1-3 secunde, de 30 de ori mai lung decat durata medie a contractiei unice a muschiului scheletic.
Forta maxima de contractie a muschiului neted este mai mare decat cea a muschiului scheletic.
In timp ce muschiul scheletic este activat exclusiv de catre sistemul nervos, muschiul neted poate fi stimulat de multiple categorii de semnale nervoase, hormonale si factori tisulari locali: a. lipsa oxigenului intr-un teritoriu tisular determina relaxarea muschiului neted vascular si vasodilatatie; b. excesul de CO2 determina vasodilatatie; c. scaderea pH-ului determina vasodilatatie; d. alti factori care determina vasodilatatie locala, precum acidul lactic, cresterea concentratiei ionilor de potasiu, scaderea concentratiei ionilor de calciu, cresterea temperaturii corpului.
O mare parte din activitatea contractila a muschiului neted este initiata fara potentiale de actiune, sub influenta factorilor enumerati.
Tesutul muscular neted visceral este constituit din fibre musculare netede, omogene, cu contractie involuntara, lenta si prelungita.
Formeaza musculatura tubului digestiv (incepand cu a doua treime a esofagului), a cailor aeriene respiratorii, a tractului genital, muschii erectori ai firelor de par, muschii corpului ciliar, stratul muscular din peretii vaselor sangvine. Fibrele sale sunt asezate in straturi si benzi sau sunt raspandite in tesutul conjunctiv. Fibrele musculare netede au aspect fusiform, 10-100 m lungime si sunt formate din sarcolema, sarcoplasma omogena si un nucleu situat central. in sarcoplasma sunt organite comune si organite specifice - miofibrile fara striatii transversale, neorganizate in sarcomere. Aceasta structura apare omogena ("neteda') la microscopul electronic.
Tesutul muscular neted multiunitar este format din fibre musculare separate si se gaseste in irisul globului ocular. Are contractii fine si limitate.
|
CARACTE-RISTICI |
MUSCHI STRIAT SCHELETIC |
MUSCHI STRIAT CARDIAC |
MUSCHI NETED VISCERAL |
MUSCHI NETED MULTIUNITAR |
|
Localizare |
atasati oaselor |
inima (miocard) |
in peretii viscerelor, vaselor de sange; erectorii firelor de par |
muschii irisului |
|
Structura fibrelor |
striate, multinucleate, neramificate |
striate, uninucleate, ramificate, cu discuri intercalare |
netede, uninucleate, fusiforme |
netede, uninucleate, fusiforme |
|
Control nervos |
voluntar |
involuntar |
involuntar |
involuntar |
|
Sarcomere |
prezente |
prezente |
absente |
absente |
|
Sistem tubular transversal |
prezent |
prezent |
absent |
absent |
|
Jonctiuni intre fibre |
prezente |
prezente |
prezente |
absente |
|
Dimensiunea fibrelor |
mare |
mare |
mica |
mica |
|
Sursa de calciu necesar contractiei |
reticulul sarcoplasmic |
reticulul sarcoplasmic; lichidul extracelular |
reticulul sarcoplasmic; lichidul extracelular |
reticulul sarcoplas-mic; lichidul extracelular |
|
Viteza de contractie |
rapida |
medie |
lenta |
lenta, progresiva |
Structura miofibrilelor
Miofibrilele, organite specifice fibrelor musculare, le confera acestora proprietati contractile.
Sunt formate din miofilamente de miozina (groase - de cea 100 Å) care formeaza discuri intunecate (impreuna cu capetele libere ale miofilamentelor de actina) si de actina (subtiri - de cea 50 Å) care formeaza discuri clare.

O miofibrila este alcatuita din aproximativ 1500 miofilamente de miozina si 3000 miofilamente de actina dispuse astfel: fiecare miofilament de miozina este inconjurat de 6 miofilamente de actina si fiecare miofilament de actina este inconjurat de 3 miofilamente de miozina.
Discurile intunecate (DI) si cele clare (DC) alterneaza si sunt asezate la acelasi nivel in toate fibrele musculare, conferindu-le acestora aspectul striat
In centrul discului intunecat se afla o portiune mai luminoasa - banda H (Hensen), iar in centrul discului clar, o portiune mai intunecata -membrana Z. Doua membrane Z succesive delimiteaza un sarcomer-unitatea morfofunctionala a fibrei musculare.
Tesutul nervos
Tesutul nervos este constituit din celule specializate (neuroni) si din celule de sustinere (celule nevroglice).
Neuronul reprezinta unitatea morfo-functionala a sistemului nervos. Din punctul de vedere al formei si al dimensiunilor, neuronii sunt foarte diferiti. Forma neuronilor este variabila: stelata (coarnele anterioare ale maduvei), sferica sau ovalara (in ganglionii spinali), piramidala (zonele motorii ale scoartei cerebrale) si fusiforma (in stratul profund al scoartei cerebrale).
In functie de numarul prelungirilor, neuronii pot fi:
. unipolari (celulele cu conuri si bastonase din retina); au aspect globulos, cu o singura prelungire;
. pseudounipolari; se afla in ganglionul spinal si au o prelungire care se divide in "T"; dendrita se distribuie la periferie, iar axonul patrunde in sistemul nervos central (SNC);
. bipolari, de forma rotunda, ovala sau fusiforma, cele doua prelungiri pornind de la polii opusi ai celulei (neuronii din ganglionii spiral Corti si vestibular Scarpa, din retina si din mucoasa olfactiva);
. multipolari; au o forma stelata, piramidala sau piriforma si prezinta numeroase prelungiri dendritice si un axon (scoarta cerebrala, cerebeloasa, coarnele anterioare din maduva spinarii).
|
Clasificarea neuronilor: |
||
|
a. Dupa forma corpului celular: |
b. Dupa numarul prelungirilor: |
c. Dupa functie: |
|
- stelati |
- unipolari |
- senzitivi |
|
- ovali |
- pseudounipolari |
- motori |
|
- piriformi |
- bipolari |
- de asociatie |
|
- piramidali |
- multipolari |
- secretori |
|
- rotunzi |
|
|
Dupa functie, neuronii pot fi: receptori, care, prin dendritele lor, receptioneaza stimulii din mediul exterior sau din interiorul organismului (somatosenzitivi si viscerosenzitivi), motori, ai caror axoni sunt in legatura cu organele efectoare (somatomotori sau visceromotori), si intercalari (de asociatie), care fac legatura intre neuronii senzitivi si motori.
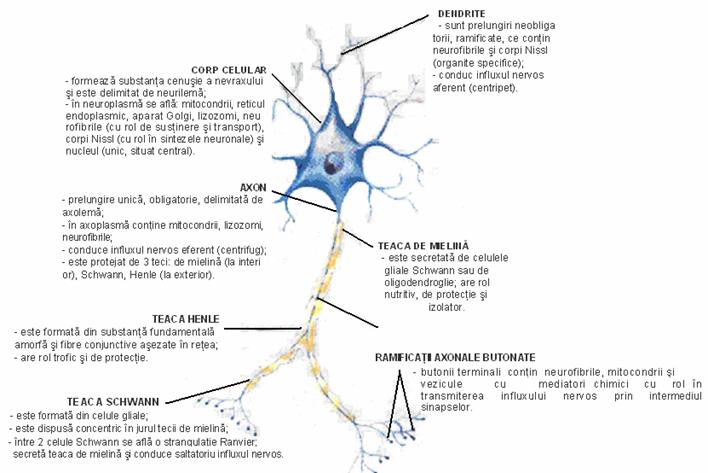
Neuronul este format din corpul celular (pericarionul) si una sau mai multe prelungiri, care sunt de doua tipuri: dendritele, prelungiri celulipete (majoritatea neuronilor au mai multe dendrite), si axonul, care, functional, este celulifug, prelungire unica a neuronului.
Corpul neuronului este format din neurilema (membrana plasmatica), neuroplasma (citoplasma) si nucleu.
Neurilema celulei nervoase este subtire, delimiteaza neuronul si are o structura lipoproteica.
Neuroplasma contine organite celulare comune (mitocondrii, ribozomi, reticul endoplasmatic, cu exceptia centrozomului, deoarece neuronul nu se divide), incluziuni pigmentare si organite specifice: corpii tigroizi (Nissl) din corpul celular si de la baza dendritelor, cu rol in metabolismul neuronal, si neurofibrilele, care se gasesc atat in neuroplasma (corp), cat si in prelungiri (dendrite si axon), avand rol mecanic, de sustinere si in conducerea impulsului nervos.
Nucleul. Celulele nervoase motorii, senzitive si de asociatie au un nucleu unic, cu 1-2 nucleoli. Celulele vegetative centrale sau periferice prezinta deseori un nucleu excentric. Aceste celule pot avea nuclei dubli sau multipli.
Dendritele, in portiunea lor initiala, sunt mai groase, apoi se subtiaza. In ele se gasesc neurofibrile. Acestea receptioneaza impulsul nervos si il conduc spre corpul neuronului.
Axonul este o prelungire unica, lunga (uneori de 1 m) si mai groasa. Este format dintr-o citoplasma specializata, numita axoplasma, in care se gasesc: mitocondrii, vezicule ale reticulului endoplasmatic si neurofibrile. Membrana care acopera axoplasma se numeste axolema si are un rol important in propagarea impulsului nervos. De-a lungul traseului sau, axonul emite colaterale perpendiculare pe directia sa, iar in portiunea terminala se ramifica; ultimele ramificatii - butonii terminali - contin mici vezicule pline cu mediatori chimici care inlesnesc transmiterea influxului nervos la nivelul sinapselor. Butonul mai contine neurofibrile si mitocondrii.
Nevroglia. La mamiferele superioare, numarul nevrogliilor depaseste de 10 ori numarul neuronilor. Forma si dimensiunile corpului celular pot fi diferite, iar prelungirile, variabile ca numar. Se descriu mai multe tipuri de nevroglii: celula Schwann, astrocitul, oligodendroglia, microglia, celulele ependimare si celulele satelite. Nevrogliile sunt celule care se divid intens (sunt singurele elemente ale tesutului nervos care dau nastere tumorilor din SNC), nu contin neurofibrile si nici corpi Nissl. Au rol de suport pentru neuroni, de protectie, trofic, rol fagocitar (microglia), in sinteza tecii de mielina si in sinteza de ARN si a altor substante pe care le cedeaza neuronului.
Inconjurand axonul, se deosebesc, in functie de localizare - sistemul nervos periferic (SNP) sau SNC - si de diametrul axonului, urmatoarele structuri:
|
Structura
|
Axonul neuronilor SNP
|
Axonul neuronilor SNC |
|
Teaca de mielina - axonii cu diametrul mai mic de 2 m si fibrele postganglionare nu au teaca de mielina. Rolul mielinei este de izolator electric, care accelereaza conducerea impulsului nervos. De asemenea protejeaza si hraneste axonul. |
- produsa de celulele Schwann (o celula produce pentru un singur axon); - prezinta discontinuitati numite noduri Ranvier, care reprezinta spatiul dintre doua celule Schwann.
|
. produsa de oligodendrocite (o celula produce pentru mai multi axoni).
|
|
Teaca Schwann are rol trofic si de secretie a mielinei si asigura conducerea saltatorie a influxului nervos.
|
. se dispune concentric in jurul tecii de mielina, fiind formata de celule Schwann; . fiecarui segment internodal de mielina |
Nu prezinta.
|
|
Teaca Henle Este un invelis continuu in jurul tecii Schwann, fiind formata din fibre conjunctive care au un rol trofic si de protectie. |
. separa membrana plasmatica a celulei . are rol in permeabilitate si rezistenta. |
Nu prezinta.
|
Fiziologia neuronului si a sinapsei
Proprietatile neuronilor:
- generarea influxului nervos (excitabilitatea)
- conducerea influxului nervos (conductibilitatea)
EXCITABILITATEA reprezinta capacitatea materiei vii de a raspunde prin manifestari specifice la actiunea stimulilor. in conditii experimentale, poate fi determinata cantitativ la animale si la om. Se caracterizeaza prin urmatorii parametri:
a), intensitatea prag a stimulilor (reobaza) reprezinta intensitatea necesara unui stimul pentru a produce un influx nervos. Stimulii cu intensitate inferioara pragului se numesc subliminari si nu produc influx nervos. Stimulii cu intensitate superioara pragului (supraliminari) au acelasi efect ca si cei cu intensitatea prag (legea "tot sau nimic')
b). timpul util reprezinta timpul minim necesar unui stimul cu intensitatea prag pentru a produce un influx nervos.
c). cronaxia este timpul minim necesar unui stimul (curent electric), avand o intensitate dubla fata de reobaza, pentru a produce un influx nervos. Are valori de 10-30 de ori mai mici decat timpul util si este cu atat mai scurta cu cat excitabilitatea este mai mare. Are valori apropiate pentru neuronii senzitivi, motori si efectori.
d). labilitatea este capacitatea neuronului de a raspunde la un anumit numar de stimuli pe unitatea de timp.
e). perioada refractara este proprietatea neuronului de a nu raspunde la un stimul nou, in timpul unui raspuns la un stimul anterior.
f). bruschetea reprezinta rapiditatea cu care actioneaza stimulul.
CONDUCTIBILITATEA reprezinta capacitatea de autopropagare a influxului nervos prin axon spre alt neuron sau spre efector.
Suportul fizico-chimic al excitabilitatii si al conductibilitatii este potentialul electric membranar.
Conform teoriei ionice a lui Julius Bemstein, acest potential electric apare ca o consecinta a repartitiei inegale a ionilor (Na+, K+, Ca2+, Cl-, etc) de o parte si de alta a membranei celulare neuronale care prezinta permeabilitate selectiva. Datorita miscarii ionice impuse de gradientul de concentratie, apar diferente de potential electric intre cele doua fete ale membranei.
In conditii de repaus (cand nu actioneaza nici un stimul), neurilema este pozitiva pe fata externa si negativa pe cea interna. intre cele doua fete exista o diferenta de potential de 70 mV numita potential de repaus (se noteaza in mod conventional -70 mV). in timpul potentialului de repaus, membrana este permeabila pentru K+ si impermeabila pentru Na+. Aplicarea unui stimul cu intensitatea prag determina depolarizarea membranei datorita cresterii permeabilitatii ei pentru Na+. Membrana devine negativa la exterior si pozitiva la interior. Diferenta de potential devine 35 mV si se numeste potential de actiune. Acesta se autopropaga. (in mod conventional, se noteaza +35 mV.) Transmiterea depolarizarii de-a lungul fibrei nervoase constituie influxul nervos.
Depolarizarea se propaga pas cu pas de-a lungul axonului, prin intermediul circuitelor locale (fluxuri circulare sau curentii lui Hermann). In axonii cu teaca de mielina, influxul nervos se propaga saltato-riu de la o strangulatie Ranvier la alta. in axonii fara teaca de mielina, influxul nervos se propaga pas cu pas. Viteza de propagare a influxului nervos in axonii mielinizati este de 50 ori mai mare decat in cei amielinici.
Ulterior, are loc repolarizarea membranei neuronale (scaderea permeabilitatii pentu Na+) si se stabileste echilibrul ionic initial. Transportul ionic se realizeaza pasiv, prin intermediul canalelor ionice (polipeptide sau lipopeptide cu greutate moleculara redusa) sau activ, prin intermediul pompelor ionice (agregat de proteine transportoare si enzime). Transmiterea unidirectioala a influxului nervos de la un neuron la altul, de la receptor la neuron si de la neuron le efector se realizeaza prin intermediul sinapselor.
Conducerea la nivelul axonilor amielinici. in acest caz, potentialul de actiune poate sa apara in orice zona a membranei. Proprietatile electrice ale membranei permit depolarizarea regiunilor adiacente, iar potentialul de actiune este condus intr-o singura directie, deoarece in directia opusa, unde s-a produs potentialul de actiune anterior, membrana este in stare refractara absoluta. De fapt, termenul de conducere este impropriu, deoarece orice nou potential de actiune este un eveniment complet nou, care se repeta, se regenereaza de-a lungul axonului. Toate potentialele de actiune au aceeasi amplitudine, de aceea se spune despre aceasta conducere ca este fara decrement (diminuare). Viteza de conducere este mai mare, daca axonul are diametrul mai mare, dar este mai mica decat in cazul axonilor mielinizati.
Conducerea la nivelul axonilor mielinizati In acest caz, datorita proprietatilor izolatoare ale mielinei, potentialul de actiune apare la nivelul nodurilor Ranvier si "sare" de la un nod la altul intr-un tip de conducere numita "saltatorie". Acest tip de conducere permite viteze mult mai mari (100-120 m/s prin fibrele mielinice groase, fata de 5-l0 m/s in fibrele mielinice subtiri si de 0,5 m/s in fibrele amielinice). Aceasta explica aparitia mai rapida a unor reflexe decat altele.
mielina
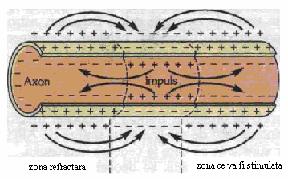
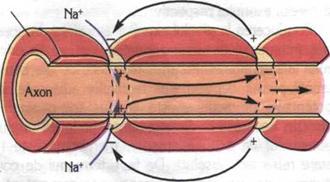
Conducerea impulsului nervos (potential de Conducerea impulsului nervos intr-o fibra mielinizata
actiune) intr-o fibra amielinica
Stimul
|
Potential de actiune ce se'autopropaga |
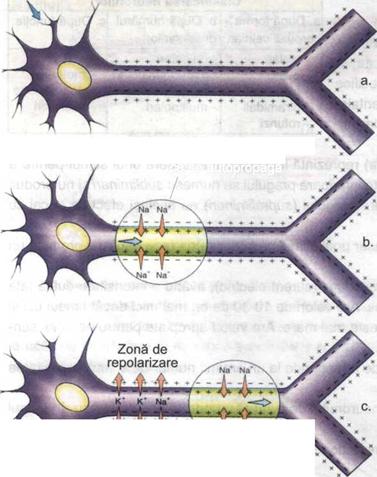
Transmiterea influxului nervos de-a lungul neuronului
a. potential de repaus
b. potential de actiune
c. repolarizarea membranei
a. potential de repaus b. potential de actiune c. repolarizarea membranei
Functionarea sinapselor
Influxul nervos, care se manifesta ca o unda de depolarizare, odata ajuns la nivelul butonilor terminali, determina fuzionarea veziculelor cu membrana presinaptica, spargerea lor si eliberarea mediatorilor chimici; acestia difuzeaza prin fanta sinaptica si ajung in contact cu fata externa a membranei postsinaptice, la nivelul receptorilor specifici. Drept urmare, membrana postsinaptica este depolarizata si permite propagarea influxului nervos. Ulterior, neurotransmitatorii sunt inactivati rapid de enzimele din fanta sinaptica. Apoi, neurotransmitatorii sunt reinglobati in vezicule sau trec in circulatia sangvina. Sinteza de noi cantitati de mediatori chimici se face pe seama ATP din mitocondriile de la nivelul butonilor terminali ai axonului. Datorita succesiunii acestor procese apare intarzierea sinaptica de 0,5-1 ms.
La nivelul sinapselor dintre receptori si neuroni, datorita fantei sinaptice foarte inguste, influxul nervos se autopropaga prin mecanismul circuitelor locale.
La nivelul placii motorii, acetilcolina se fixeaza prin receptori specifici de sarcolema depola-rizand-o si determinand un potential local terminal de placa. Acesta va genera potentiale de actiune ce se vor propaga de la placa motorie in toate directiile.
|
Acetilcolin-esteraza |
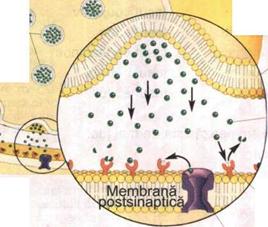 Structura
unei sinapse
Structura
unei sinapse
|
Vezicule sinaptice cu mediator |
Componenta presinaptica (buton
axonal)
contine vezicule cu mediator chimic
(neurotransmitator).
|
Membrana presinaptica |
Fanta (spatiul) sinaptica
|
Mediator chimic |
reprezinta spatiul dintre membrana plasmatica a butonilor axonali si cea a componentei postsinaptice. Are 200 - 300 Å
Componenta postsinaptica
este reprezentata de corpul celular, dendritele sau portiunea initiala a axonului unui neuron, respectiv de sarcolema fibrei musculare striate.
Receptor
Clasificarea sinapselor:
![]()
![]()
![]() a.Cu transmitere chimica
a.Cu transmitere chimica
|
1. Din punct de vedere functional: |
- adrenergice (mediatorul
chimic este adrenalina sau
noradrenalina)
-colinergice (mediatorul
chimic este acetilcolina)
b.Cu transmitere electrica
|
2. Din punct de vedere al efectului: |
![]() a.Excitatorii
a.Excitatorii
b.lnhibitorii
![]()
a.lnterneuronale
|
3. Dupa structurile implicate: |
- axo-somatice
- axo-dendritice
- axo-axonale
b.Neuron-receptor
c. Neuron-efector(de exemplu,
placa motorie)
![]()
Sinapsele neuro-neuronale pot fi axosomatice sau axodendritice, axoaxonice sau dendrodentritice.
Din punct de vedere al mecanismului prin care se face transmiterea, sinapsele pot fi chimice sau electrice.
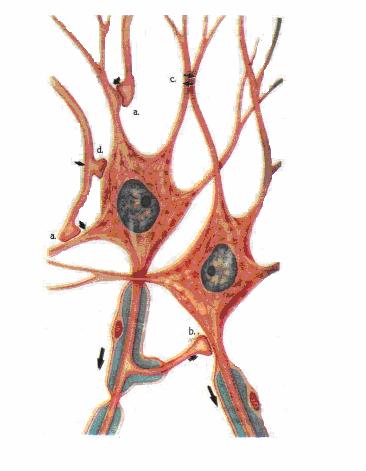
Tipuri de sinapse: a. axodendritica; b. axoaxonica;
c. dendrodendritica (electrica); d. axosomatica.
|
|
Sinapse chimice |
Sinapse electrice |
|
Alcatuire
|
- terminatia presinaptica - contine vezicule cu mediator chimic (se cunosc peste 40 de mediatori chimici, cel mai raspandit fiind acetilcolina); - fanta sinaptica; - celula postsinaptica - prezinta receptori pentru mediatorul chimic.
|
- doua celule de aceleasi dimensiuni, care sunt alipite in zonele lor de rezistenta electrica minima.
|
|
Mod de functionare
|
. sub actiunea impulsului nervos, se elibereaza cuante de mediator chimic in fanta sinaptica; . mediatorul chimic interactioneaza cu receptorii specifici de pe membrana postsinaptica, determinand modificari ale potentialului membranei postsinaptice. Conducerea este unidirectionala dinspre terminatia presinaptica spre cea postsinaptica. |
. trecerea ionilor si a moleculelor prin aceste locuri de jonctiune. Conducerea este, se pare, bidirectionala.
|
|
Exemple
|
. aproape toate sinapsele SNC; . placa motorie; . SN vegetativ. |
. miocard; . muschi neted; . in anumite regiuni din creier. |
In urma interactiunii dintre mediatorul chimic eliberat in fanta sinaptica si receptorii de pe membrana postsinaptica, apare depolarizarea membranei postsinaptice, numita potential postsinaptic excitator, daca este vorba de un neuron postsinaptic, sau potential terminal de placa daca este vorba despre o fibra musculara scheletica. Acest potential, care nu trebuie confundat cu potentialul de actiune, are doua proprietati speciale: sumatia temporala si sumatia spatiala. In primul caz, doua asemenea potentiale produse prin descarcarea de mediator din aceeasi fibra presinaptica se pot suma, rezultand un potential mai mare, iar in cel de-al doilea caz, potentialele postsinaptice excitatorii, produse de doua terminatii presinaptice vecine pe aceeasi membrana postsinaptica, se pot cumula. Alaturi de sinapsele excitatorii, exista si sinapse inhibitorii, in functie de tipul de mediator chimic continut de terminatia presinaptica. Mediatorul chimic inhibitor determina, prin legarea de receptorii sai de pe membrana postsinaptica, declansarea unor mecanisme care au drept consecinta hiperpolarizarea membranei postsinaptice, cu instalarea potentialului postsinaptic inhibitor si realizarea inhibitiei postsinaptice.
In afara de inhibitia postsinaptica, se poate produce si un alt tip de inhibitie, inainte ca semnalul sa ajunga la nivelul sinapsei. Aceasta, denumita inhibitie presinaptica se datoreaza unor sinapse "presinaptice", aflate pe terminatia presinaptica, inainte de contactul sau cu neuronul urmator.
Oboseala
transmiterii sinaptice. Stimularea
repetata si rapida a sinapselor excitatorii este urmata de
descarcari foarte numeroase ale neuronului postsinaptic, pentru ca, in
urmatoarele milisecunde, numarul acestora sa scada accentuat. in acest caz,
avem de-a face cu un mecanism de protectie impotriva
supraexcitarii, care se realizeaza prin epuizarea depozitelor de mediator
chimic (neurotransmitator) de la nivelul terminatiei presinaptice.
Efectele drogurilor asupra transmiterii sinaptice. Unele droguri cresc excitabilitatea sinapselor (cofeina), altele o scad (unele anestezice). Anumiti factori pot influenta transmiterea influxului nervos prin sinapse. Astfel, potasiul stimuleaza transmisia influxului nervos la nivelul sinapsei, iar anumite otravuri (curara sau matraguna) pot bloca transmiterea excitatiei; in astfel de cazuri se produc paralizii musculare sau stop respirator. Paraliziile pot surveni si in urma toxiinfectiilor alimentare, cand bacteriile secreta toxine care inhiba secretia de mediatori chimici. Nicotina si alcoolul au actiune inhibitoare, stricnina - excitatoare, iar drogurile si substantele halucinogene actioneaza direct asupra sinapselor din creier, afectand coordonarea nervoasa si comportamentul individului. Unele substante (exemplu, otrava E 605 folosita impotriva insectelor) inhiba redescompunerea mediatorului chimic; musculatura ramane excitata timp mai indelungat, iar intregul organism este cuprins de spasme musculare, si in final, intervine decesul.
In maladia Alzheimer, caracterizata prin pierderea memoriei si alterarea intelectului, se constata o concentrare a acetilcolinei in scoarta cerebrala, in zona implicata in mecanismul memoriei.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 15400
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved