| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Deletiile 1p. Studii recente au sugerat existenta in tumorile colorectale a uneia sau mai multor gene tumorosupresoare in zona 1p. Un studiu al LOH al acestei zone prin PCR (Matsuzaki, M., Nagase, S., Abe, T. si colab., 1998) a identificat trei regiuni de deletie: 1p36.3, 1p35.1-p36.3, 1p34.2-p35.
Deletiile 8p. Exista dovezi ale prezentei a cel putin 2 gene tumorosupresoare pe bratul p al cromozomului 8 in tumorile colorectale. Fujiwara si colab. (1993) au identificat doua astfel de deletii la 44 % din pacientii cu cancer colorectal, una localizata in zona 8p23.2-8p22, iar cealalta la 8p21.3-8p11.22. Pierderea heterozigotiei 8p a fost corelata cu stadiul clinicopatologic, sugerand un posibil rol al genelor tumorosupresoare 8p in progresia maligna. Un alt grup (Lerebours si colab., 1999) a identificat o deletie in zona 8p21 ce implica genele LPL (lipoprotein lipaza) si NEFL (gena lantului usor al neurofilamentelor). Studiul a permis identificarea unui interval flancat de locusul D8S1771 si gena NEFL, unde ar putea fi localizata o gena tumorosupresoare.
Deletiile 17p. LOH 17p a fost corelata cu microinvazia limfatica si/sau vasculara din stadiile avansate ale carcinoamelor colorectale (Takanishi si colab., 1995), cu stadiul D Dukes si prezenta metastazelor la distanta (Khine, K., Smith, DR, Goth, DS, 1994). Un studiu al corelatiei deletiilor 17p cu progresia neoplazica (Risio si colab., 2003) a identificat deletii centromerice in displazia de mucoasa (40 %), cancerul incipient (50 %) si avansat (10 %) (Risio si colab., 2003). Deletiile subtelomerice au fost observate in 60 % din cazurile de cancer avansat si in 15 % din cancerele incipiente.
Deletiile 18q. Rezultatele studiilor privind corelatia dintre deletiile 18q si cancerul de colon nu sunt semnificative. Se inregistreaza o variabilitate considerabila in functie de metoda utilizata, markerul folosit si pragul dezechilibrului alelic (AI) ales (Popat, S., Houlston, R.S., 2005). Cancerele cu deletii 18q par a avea un prognostic mai prost.
Formarea unei tumori solide este initiata de o serie de modificari somatice mostenite - mutatii sau evenimente epigenetice transmisibile (epimutatii) cum ar fi metilarea insulelor CpG (Baylin, 1997). Unele dintre aceste modificari produc un avantaj de crestere fata de celulele inconjuratoare. Acest proces poate fi accelerat de instabilitatea genetica sau fenotipul mutant (Jackson si Loeb, 1998; Tomlinson si Bodmer, 1999).
Daca intr-o populatie normala de celule in diviziune o celula capata un avantaj de crestere printr-o mutatie sau epimutatie, va avea tendinta sa se multiplice clonal in detrimentul celulelor inconjuratoare, luand nastere astfel ozona de tesut anormal (patch). In interiorul acestei zone poate avea loc o a doua modificare de acest tip, astfel incat o celula sa capete un avantaj de crestere fata de celelalte celule, formandu-se astfel o a doua zona in interiorul primei. Procesul se poate repeta de cateva ori, pana la aparitia unei celule maligne care se multiplica clonal in cancer (figura 1). Deci in general tumorile se asociaza si sunt precedate de o anomalie ce reflecta succesiunea de evenimente premaligne.
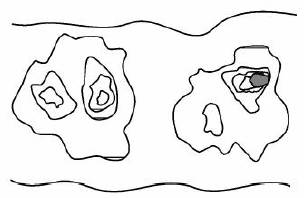
Figura 1. Zone de epiteliu colonic anormal ce se extind pornind de la defecte punctiforme, si progreseaza spre adenocarcinom. Zona neagra reprezinta un adenocarcinom.
Mutatiile somatice sau epimutatiile ce contribuie la progresia spre adenocarcinom pot apare prin interactiunea celulelor cu mediul (de exemplu componente ale fumului de tigara, carcinogeni alimentari) sau prin producerea excesiva de agenti endogeni ce produc stress (de exemplu secretia crescuta de acizi biliari asociata cu o dieta bogata in grasimi). Mutatiile liniei germinale ale unor gene cum este APC pot predispune individul la cancer. Pe acest fond genetic, mutatiile sau epimutatiile ulterioare vor genera un defect punctiform, asa cum se observa in figura 1.
In general, un avantaj de crestere se poate obtine fie prin scaderea capacitatii de intrare in apoptoza fie prin cresterea numarului de diviziuni celulare. Ambele tipuri de modificari vor conduce nu numai la o expansiune clonala, dar vor creste si probabilitatea aparitiei de mutatii. Se observa frecvent o scadere a apoptozei in cancerele de colon (Bedi et al., 1995). Celulele normale care au suferit leziuni ADN si care sunt incapabile sa le repare, intra in mod normal in apoptoza. Acest proces de "sinucidere" celulara impiedica replicarea ADN defectuos, cu mare probabilitate de a genera o mutatie in ADN din celulele fiice. Deci o celula cu un defect in initierea apoptozei ca raspuns la leziunile ADN nereparate nu numai ca va avea un avantaj de crestere, dar va avea o probabilitate crescuta de a dezvolta mutatii
Datorita acestor considerente, este foarte probabil ca o zona de celule defective sa contina mutatii si/sau alterari epigenetice care sa induca rezistenta la apoptoza si/sau rata de diviziune crescuta.
Acestea sunt gene care functioneaza normal in sensul suprimarii transformarii maligne. Spre deosebire de oncogene, inactivarea sau pierderea acestor gene este cea care duce la transformarea maligna. Dovezile in ce priveste rolul genelor tumorosupresoare in carcinogeneza colorectala se bazeaza pe constatarea ca in majoritatea tumorilor a fost observata deletia in anumite zone cromozomiale. Aceasta constatare promoveaza ipoteza ca aceste zone cuprind gene ale caror produse regleaza in mod normal cresterea intr-un sens negativ. Primele dovezi clare ca inactivarea genelor tumorosupresoare este un eveniment comun in dezvoltarea tumorilor colorectale au aparut in studiul genei Polipozei Adenomatoase Colonice (APC).
Proto-oncogena src codifica o protein-tirozin kinaza citoplasmatica nereceptoare. Nivelele protein kinazei src au fost raportate crescute in in cancerul de colon (Bolen si colab., 1987; Cartwright si colab., 1989).
Gena p53 se gaseste in cromozomul 17p. Produsul genei p53 este o proteina de 53 kD, de unde si numele. Proteina p53 are rolul de a opri ciclul celular daca ADN-ul este lezat sau celula a suferit alt tip de leziune, prin legarea de un factor de transcriptie numit E2F. Legarea proteine p53 la E2F impiedicand legarea acestuia la promotorii proto-oncogenelor c-myc si c-fos. Transcriptia acestor gene este necesara pentru initierea mitozei, astfel ca proteina p53 blocheaza ciclul celular. Daca leziunile sunt minore, oprirea ciclului celular ii da celulei timp de reparatii, daca sunt mai grave p53 semnalizeaza intrarea celulei in apoptoza.
P53 sufera o mutatie in 75-83 % din carcinoamele colorectale, dar nu foarte frecvent in adenoame (Baker, Fearon, Nigro si colab., 1989). Deci mutatia genei p53 poate fi asociata evolutiei de la adenom la carcinom. Rolul genei p53 se crede ca este acela de a regla cresterea normala prin controlul expresiei unui numar de gene celulare. Pierderea functiei normale a p53 poate genera instabilitate in celule. Mutatiile p53 sunt asociate cu cresterea duratei de viata a proteinei p53, ducand la acumularea acesteia. Aceasta acumulare poate fi detectata cu anticorpi specifici, permitand astfel identificarea celulelor cu mutatii p53.
Metilarea - de cautat
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 1629
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2024 . All rights reserved