| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Plasmidele
Plasmidele sunt structuri genetice separate fizic de cromosomul bacterian, capabile de replicare independenta de cromosom, adica entitati cu capacitati de replicare autonoma sau repliconi. Ele contin informatie genetica neesentiala pentru cresterea si diviziunea celulei, care poate fi castigata sau pierduta fara ca existenta celulei sa fie afectata. Autonomia lor fata de cromosom este relativa: sunt separate fizic de cromosom, dar in ceea ce priveste replicarea, sunt permanent controlate de gene cromosomale.
In categoria plasmidelor tipice intra
factorii genetici F,
Clasificarea plasmidelor
Plasmidele se denumesc si se clasifica dupa efectul lor cel mai evident asupra celulei (conferirea proprietatii de donor de material genetic, sinteza de colicine, rezistenta la antibiotice etc.). Acest criteriu de clasificare nu este satisfacator, deoarece, uneori o plasmida confera mai multe proprietati celulei in care se gaseste: de exemplu, plasmida R ii confera rezistenta la antibiotice, dar in acelasi timp celula poate dobandi si calitatea de donor de material genetic. Din acest motiv se folosesc si alte criterii de clasificare.
Plasmidele se grupeaza dupa criteriul "incompatibilitatii" lor, adica al capacitatii de a exclude din celula alte plasmide cu aceiasi secventa de baze, sau una foarte apropiata. Altfel spus, plasmidele cu acelasi tip de informatie genetica sunt incompatibile, adica nu pot coexista intr-o celula. Clasificarea bazata pe compararea secventei de baze este mult mai practica. In acelasi scop se poate folosi comparatia secventei de aminoacizi a proteinelor Rep, deoarece sunt codificate de majoritatea plasmidelor si au functii comune.
In functie de capacitatea lor de a media transferul de material genetic prin conjugare, plasmidele se impart in doua categorii.
l. Plasmidele conjugative (transmisibile sau infectioase), denumite si factori de sex, sunt acelea care confera celulei purtatoare de plasmida, proprietatea de donor de material genetic (celula "mascul"), in raport cu o celula care nu poseda o astfel de plasmida si care se comporta ca receptor (celula "femela"). Plasmidele conjugative se mai numesc conjugoni, factori de fertilitate sau transferoni. Ele au in structura genetica, pe langa genele de replicare autonoma, determinantii genetici de transfer (tra). Din aceasta categorie fac parte plasmidele de sex (F, F', Hfr), plasmidele Ent, unele plasmide Col si unele plasmide R. Plasmidele conjugative au cel putin 30 kbp.
2. Plasmidele neconjugative nu sunt autotransmisibile deoarece nu confera celulei purtatoare proprietatea de donor de material genetic, neavand determinantii genetici "tra". Din aceasta categorie fac parte celelalte plasmide Col si R. Ele se pot transfera de la o celula la alta, fie prin intermediul unui fag transductor, fie prin procesul de conjugare initiat de o plasmida conjugativa, coexistenta in aceiasi celula.
Dupa criteriul capacitatii lor de a se integra in cromosomul celulei, se disting doua categorii de plasmide.
l. Plasmidele episomale (episom = corp adaugat) sau integrative pot exista in celula, atat in stare autonoma (fizic independenta) de cromosom, fie in stare integrata in cromosomul bacterian. Din aceasta categorie fac parte plasmidele F si Col E1. Cele doua stari sunt reversibile si alternative: dupa integrare in structura cromosomului, ele pot trece din nou in stare autonoma si redevin fizic independente, printr-o excizie corecta, sau printr-o excizie incorecta, cand plasmida se desprinde din insertia cromosomala impreuna cu cateva gene ale acestuia.
2. Plasmidele neintegrative nu se integreaza in cromosomul bacterian, ci persista indefinit numai in stare autonoma.
Structura moleculara a plasmidelor
Plasmidele sunt cromosomi miniaturali (minicromosomi) alcatuiti din molecule de ADN dublu catenare, circulare, inchise covalent, care reprezinta l-2% din marimea cromosomului. O plasmida poate sa contina una sau cateva sute de gene. ADN plasmidial se gaseste in celulele bacteriene in trei forme fizic diferite (fig. 114).
l. Forma circulara inchisa covalent (denumita C C C)(Covalently Closed Circular), care prezinta in plus 1-2 torsiuni ce dau moleculei aspectul de suprahelice.
2. Forma circulara deschisa, avand o catena deschisa si una inchisa.
3. Plasmidele concatemere sau oligomere concatenate, sunt complexe supramoleculare formate din cateva plasmide si sunt rezultatul unor erori de replicare a monomerilor circulari sau a proceselor de recombinare interplasmidiala. Daca ambele catene sunt inchise, componentele individuale ale complexului sunt suprahelicale, iar daca una dintre catene este deschisa, forma este circulara. Configuratia circulara este o conditie a existentei lor in celula, cea care le confera rezistenta la actiunea nucleazelor celulare. Totusi, s-au identificat plasmide cu configuratie lineara (la Streptomyces, Rhodococcus).
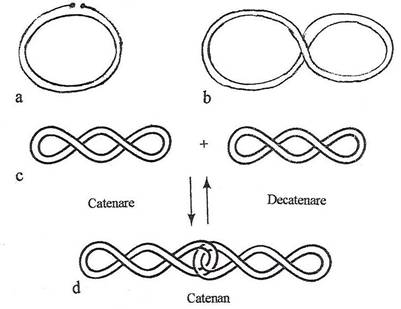
Fig. 108. Configuratii diferite ale ADN plasmidial. a. Plasmida circulara cu o catena incizata. b, c. Plasmide covalent inchise superhelicale. d. Plasmide concatenate.
Structura genetica si functiile plasmidelor
Plasmidele contin urmatoarele categorii de determinanti genetici:
l. Gene esentiale pentru existenta lor ca repliconi fizic independenti, adica genele implicate in replicarea acestor structuri genetice.
2. Gene de specificitate pentru incompatibilitate. Incompatibilitatea corespunde situatiei in care, doua plasmide omologe nu pot fi mentinute stabil in aceiasi celula, deoarece una o exclude pe cealalta. Tehnica hibridarii ADN-ADN evidentiaza o omologie neta intre plasmidele din acelasi grup de incompatibilitate si o foarte mica asemanare intre plasmidele compatibile. Pe baza incapacitatii de a coexista in aceiasi celula, plasmidele au fost grupate in grupe de incompatibilitate.
3. Gene structurale, in numar variabil, care confera celulei proprietati noi.
4. Suprafata de legare de mezosomi, care asigura corelarea replicarii plasmidelor cu ciclul celular, precum si repartizarea lor echilibrata in cele doua celule fiice.
5. Elemente genetice transpozabile (secvente de insertie si transpozoni).
6. Plasmidele conjugative au determinanti genetici de transfer (tra).
Plasmidele au dimensiuni variate: l - 400 kb (echivalentul a l0% din cromosomul de E. coli). Cele mai mici au ADN echivalent pentru 2-3 gene. Genele plasmidiale nu confera un fenotip detectabil celulei purtatoare si de aceea se numesc gene criptice.
De cele mai multe ori, prezenta plasmidelor confera celulelor purtatoare, proprietati noi, concretizate intr-o gama larga de functii, dintre care, cele mai importante sunt urmatoarele:
rezistenta la una sau la mai multe grupe de antibiotice (ampicilina, streptomicina, tetraciclina, kanamicina etc.) si la sulfamide;
rezistenta la cationii metalelor grele (la ionii de Hg2+ si combinatiile sale orgnomercurice, la ionii de Ni, Co, Pb, Cd, Zn, Ag, At);
rezistenta la anioni: arseniat, arsenit, telurit, borat, cromat;
rezistenta la compusii de intercalare(acridina, etidiu) si la radiatiile UV;
proprietati noi de biosinteza: sinteza antibioticelor si bacteriocinelor;
proprietati metabolice noi: metabolismul unor glucide simple (lactoza, sucroza, rafinoza), al compusilor complecsi (octan, toluen, camfor, 2,4-diclor-toluen), al proteinelor (caseina, gelatina);
producerea de toxine (enterotoxina la E. coli, toxina exfoliativa la S. aureus, neurotoxina la C. tetani) si prin ele, proprietati noi de virulenta;
sinteza antigenelor de colonizare la E. coli (antigenele fimbriale K88 si K99);
sinteza materialului capsular la B. anthracis;
inducerea tumorilor de colet la plante (plasmida Ti la A. tumefaciens) si sinteza de catre celulele tumorii, a derivatilor azotati din aminoacizi, denumiti opine (octopina, derivat al argininei si nopalina);
infectia si nodularea plantelor leguminoase (plasmida Sym la Rhizobium);
proprietati conjugative: plasmide ce codifica sinteza pilinei si asamblarea pililor (si implicit dobandirea sensibilitatii la fagii ARN masculi, care se leaga specific de pili);
alte proprietati: formarea vacuolelor cu gaz la Halobacterium, variatia translucid/opac a coloniilor de Mycobacterium, producerea H2S la Enterobacteriaceae.
Genele localizate pe plasmide inhiba exprimarea genelor cromosomale omologe.
Controlul numarului de copii plasmidiale
Intr-o celula bacteriana se gaseste un numar diferit de copii plasmidiale: una - cateva/celula, sau un numar de l0-l00 copii. Numarul difera in functie de tipul de control pe care il exercita cromosomul asupra replicarii plasmidelor, de starea fiziologica a celulei, de dimensiunile plasmidelor, dar, in esenta, numarul de copii se regleaza prin controlul ratei de initiere a sintezei ADN. Daca o celula este transformata cu o plasmida ce se afla intr-un numar mic de copii in celula de origine, ea se va replica odata sau de doua ori inainte de diviziunea celulei. Daca transformarea se face cu o plasmida, care in celula de origine se gaseste intr-un numar mare de copii, ea se va replica in mod repetat pana este atins numarul caracteristic de copii.
Explicatia acceptata pentru reglarea numarului de copii este urmatoarea: plasmida codifica un inhibitor, cu rol reglator negativ (inhibitor) asupra initierii replicarii. Activitatea inhibitorului este dependenta de concentratie. Pe masura ce celula creste, concentratia de inhibitor scade si replicarea nu mai este inhibata. Dupa replicare, numarul moleculelor de ADN plasmidial se dubleaza. In acelasi timp, se dubleaza numarul de gene codificatoare ale sintezei inhibitorului si prin sinteza proteica, concentratia inhibitorului. Consecinta este stoparea replicarii.
Secventa de evenimente este aceiasi pentru cazul in care transformarea celulei s-a facut cu o plasmida ce realizeaza un numar mare de copii/celula. Probabil ca, pentru plasmidele care se gasesc in numar mare de copii/celula, efectul inhibtor necesita o concentratie mai mare a factorului inhibitor, decat in cazul plasmidelor cu un numar mic de copii.
Amplificarea numarului de copii plasmidiale/celula, semnifica cresterea majora a numarului de copii, in raport cu situatia obisnuita. Amplificarea este consecinta tratamentelor chimice sau a manipularilor genetice ale celulei bacteriene. Faptul este extrem de util in domeniul ingineriei genetice celulare cu scopuri biotehnologice. Se creeaza posibilitatea ca o gena utila, transferata artificial sau existenta in mod natural in structura plasmidei, sa realizeze un numar mare de copii/celula si sinteza unui anumit produs, cu o rata corespunzator mai inalta. De exemplu, sinteza catalazei poate fi amplificata de pana la 25 de ori.
Incompatibilitatea plasmidelor
Cuplurile de plasmide strans inrudite nu pot fi mentinute stabil in descendenta unei celule, deoarce sunt incompatibile. Faptul este explicabil prin modelul inhibitorului, a carui concentratie este esentiala pentru initierea replicarii. De exemplu, o celula care contine doua plasmide, F si Col E1 sintetizeaza inhibitori diferiti. Replicarea fiecarui tip de plasmida se va desfasura independent, deoarece inhibitorul unei plasmide nu regleaza replicarea celeilalte. Astfel, plasmidele F si Col E1 sunt compatibile si apartin unor grupe diferite de incompatibilitate.
Invers, doua variante ale aceleiasi plasmide nu pot fi mentinute stabil in aceiasi celula, adica sunt incompatibile. Explicatia consta in faptul ca, fiecare plasmida codifica sinteza propriului inhibitor de replicare, care este activ nu numai asupra repliconului codifcator ci si asupra celuilalt replicon. Astfel, numarul moleculelor plasmidiale in celula, este mentinut la un nivel inferior sumei copiilor potentiale ale celor doua plasmide.
Plasmidele cu sisteme neinrudite de control al replicarii sunt compatibile. Ele apartin unor grupe diferite de incompatibilitate, iar cele incompatibile apartin aceluiasi grup.
Eliminarea plasmidelor
Eliminarea plasmidelor din celula sau procesul de "vindecare" se poate realiza spontan sau cu o frecventa superioara, prin tratarea celulelor cu substante care interfera selectiv cu replicarea lor, fara sa modifice dinamica replicarii cromosomului. Pierderea plasmidelor nu afecteaza viabilitatea celulei, datorita caracterului neesential al informatiei lor genetice. Eliminarea plasmidelor este realizata cu oarecare eficienta de acriflavina, rifampicina, bromura de etidiu, ionii de cobalt. Unii agenti chimici au actiune selectiva, fiind foarte eficienti pentru eliminarea unor plasmide (acridina elimina plasmida F), fara sa afecteze pe altele. Din punct de vedere practic, actiunea selectiva este importanta, in primul rand pentru eliminarea plasmidelor de rezistenta la antibiotice. Selectivitatea actiunii unor agenti chimici se explica prin compozitia diferita in baze, a plasmidelor fata de cromosom. Quinolonele sunt recunoscute pentru eficacitatea lor ca agenti de eliminare a plasmidelor, dar nu exista nici un agent chimic de "vindecare" a tuturor plasmidelor.
Replicarea plasmidelor
Studiul replicarii plasmidelor a dus la descoperirea ARN antisens. Plasmidele au o regiune esentiala ce contine genele implicate in replicare si controlul ei, care teoretic, sunt singurele gene obligatorii ale unei plasmide. In regiunea esentiala a unei plasmide sunt concentrate cateva gene si secvente:
originea replicarii (ori), caracteristica fiecarui replicon. La nivelul regiunii ori se gasesc secvente specifice cu care interactioneaza proteina Rep(proteina de initiere a replicarii). Aici, cele doua catene se pot separa pentru initierea replicarii si incepe sinteza catenei leading. In multe cazuri, originea replicarii contine secvente repetate direct denumite iteroni. Ele sunt situsurile de legare ale proteinei Rep, se gasesc si in afara originii replicarii, avand si rol in controlul replicarii ;
gena codificatoare a proteinei Rep, existenta la multe plasmide;
secventele cu rol in controlul replicarii.
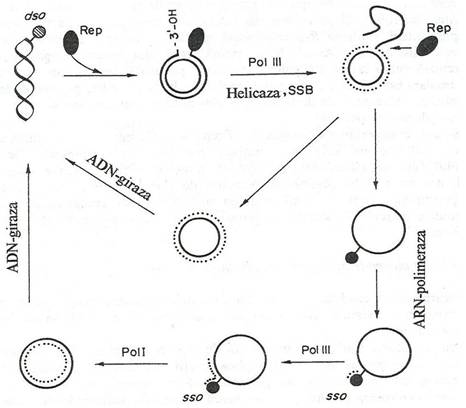
Fig. 109. Replicarea plasmidelor dupa modelul "cercului rotativ". Proteina Rep codificata de plasmida recunoaste originea dublu catenara (dso) a ADN superhelical si produce o clivare situs specifica, generand un capat 3'OH. Capatul 3'OH este alungit de proteinele de replicare, iar catena parentala este dislocata. Cand bifurcatia de replicare ajunge la situsul dso, proteina Rep catalizeaza o reactie de transfer a catenei, eliberand un intermediar monocatenar de ADN si o molecula dublu catenara, cu o catena parentala si una nou sintetizata (cercul punctat). Pe molecula circulara monocatenara este initiata sinteza unei catene lagging, la situsul monocatenar de origine (sso), catalizata de ARN-polimeraza. Enzima va sintetiza un primer scurt de ARN, iar sinteza catenei lagging este catalizata de ADN-polimeraza. Produsele sintezei sunt doua molecule de ADN superhelicale (dupa Bennett, 1998).
Plasmidele se replica fizic autonom fata de cromosomul bacterian, dar functional, replicarea este total sau partial dependenta de proteinele codificate de genele cromosomale. Majoritatea plasmidelor sunt circulare. Plasmidele lineare s-au gasit la bacteriile Gram pozitive si Gram negative.
Initierea replicarii necesita asamblarea replisomului, format din ADN polimeraza III, o ADN helicaza si primaza (proteina Rep).
S-au propus 3 mecanisme generale de replicare ale plasmidelor circulare: replicarea dupa modelul theta, replicarea prin deplasarea catenei si replicarea dupa mecanismul cercului rotativ.
Plasmidele circulare se replica semiconservativ si isi pastreaza forma circulara pe toata durata ciclului replicativ.
Replicarea dupa modelul theta (al cercului simplu) este foarte frecventa la plasmidele bacteriilor Gram negative si incepe la un punct denumit originea replicarii (ori), prin incizia ambelor catene. Evenimentele timpurii sunt:
deschiderea celor doua catene la secvente specifice ("ori"), catalizata de proteina Rep;
sinteza primerilor ARN.
De cele mai multe ori, initierea replicarii necesita o proteina initiatoare, codificata de plasmida (proteina Rep). Proteina Rep recunoaste specific si se asociaza cu secventa de origine a replicarii. Regiunea ori contine secvente de baze repetate in ordine directa, denumite iteroni. Iteronii sunt esentiali nu numai pentru replicare, dar si pentru controlul replicarii. Cele doua catene, dupa incizie au rolul de matrite pentru sinteza catenelor noi. Bifurcatia de replicare se deplaseaza uni- sau bidirectional. Ambele catene se replica simultan.
Replicarea prin deplasarea catenei necesita 3 proteine codificate de plasmida, pentru initierea replicarii ADN. Replicarea este initiata la "origine" si progreseaza in oricare din cele doua directii prin mecanismul deplasarii catenei. Cele 3 proteine codificate de plasmida (Rep A, Rep B, Rep C) au rol de helicaza, primaza si respectiv de initiere.
Replicarea incepe de la doua origini simetrice si adiacente monocatenare (ssi A si ssiB). Replicarea incepe cand aceste origini sunt accesibile ca regiuni monocatenare. Despiralizarea este dependenta de doua proteine de replicare, Rep C si Rep A si este usurata de o secventa bogata in A-T care precede regiunile ssi A si ssiB. Rep C recunoaste secventele repetate ale regiunii adiacente secventei bogate in A-T, iar Rep A este o helicaza. Rep B are rol de primaza si este specifica plasmidei. Sinteza fiecarei catene este continua si se face cu deplasarea catenei complementare. Replicarea catenei deplasate se initiaza la originea ssi.
Replicarea dupa modelul "cercului rotativ" este unidirectionala, deoarece sinteza celor doua catene este decalata in timp. Proteina Rep creeaza o bresa la secventa dso (double strand origin) si genereaza gruparea 3'OH, cu rol de primer terminal. Capatul 3'OH al catenei de polaritate negativa are rol de primer pentru sinteza catenei "leading", fiind alungit prin polimerizare pe catena circulara de polaritate opusa, cu rol de matrita. Catena "negativa" este indepartata, pe masura ce replicarea progreseaza. Alungirea capatului 3'OH al catenei "negative" continua pana cand complexul enzimatic de replicare (replisomul) a parcurs intregul cerc al matritei "pozitive". Dupa ce intreaga catena circulara "pozitiva" a fost copiata intr-o catena complementara, proteina Rep catalizeaza o reactie de transfer si catena "negativa" este circularizata. Astfel, intr-o prima etapa, din procesul replicarii, rezulta o molecula circulara dublu catenara si catena "negativa" parentala, de asemenea circulara. Catena "negativa" circulara este ulterior convertita la molecula dublu catenara, pornind de la o origine proprie a replicarii. Mecanismele de replicare a plasmidelor sunt comune si pentru replicarea genomului unor dezoxiribovirusuri.
Plasmidele actinomicetelor
La actinomicetele miceliene s-a identificat o larga varietate de plasmide diferite, majoritatea conjugative. Ele nu contin gene de rezistenta sau pentru alte particularitati metabolice, ci contin numai gene de replicare si fertilitate. Unele contin genele codificatoare ale sintezei antibioticelor.
Unele sunt plasmide mari (cateva sute de kbp) si adeseori codifica pentru caile de biosinteza ale antibioticelor. Ele se replica de la punctul de origine, localizat central si poarta proteine legate de secventele repetitive de la ambele capete. Aceste plasmide par sa se recombine frecvent cu cromosomul linear, rezultatul fiind schimbul secventelor terminale ale celor doua structuri. Extremitatile cromosomului de Streptomyces nu contin gene esentiale si de aceea pierderea acestor gene nu interfera cu viabilitatea. Schimbul fragmentelor de ADN plasmidial si cromosomal poate fi o cale foarte eficienta de diseminare a genelor prin transfer pe orizontala.
Plasmidele integrative de la Streptococcus pot fi excizate din cromosom si devin autonome. In stare autonoma se replica dupa modelul cercului rotativ sau dupa modelul theta). Integrarea se face prin recombinare la situs specific, mediat de o integraza codificata de plasmida, la un situs ce corespunde unei gene cromosomale pentru sinteza ARNt. Diferitele plasmide se integreaza in diferite gene pentru ARNt. Deoarece genele pentru ARNt sunt bine conservate la bacterii, spectrul de gazda al plasmidelor integrative este mai larg decat al celor care se replica autonom.
Sinteza ADN linear necesita prezenta unui primer, care in mod obisnuit este generat de o ARN-polimeraza ce se asociaza cu ADN si sintetizeaza o molecula scurta de ARN. ARN-polimerazele nu copiaza secventa situsului la care ele se leaga. Cum sunt copiate capetele acestor structuri ? Pentru cromosomii eucariotelor (care contin ADN linear), impedimentul este depasit prin prezenta secventelor repetitive la capete (telomere). Copii ale acestei secvente pot fi adaugate dupa replicare, sub actiunea enzimelor specifice (telomeraze).
La Streptomyces, replicarea ADN linear al plasmidei (fig. 110) este initiata de o proteina cu rol de primer, atasata la capatul 5' al fiecarei catene (fig. 112). Capetele 3' sunt libere si sunt sensibile la degradarea cu exonucleaza 3'. La diferite tulpini de Streptomyces producatoare de antibiotice, s-au gasit plasmide gigante (180-590 kb), pe care sunt plasate genele codificatoare ale antibioticelor. Initierea replicarii se face prin asocierea acestei proteine cu o a II-a molecula a aceleiasi proteine, legata covalent cu o nucleotida. Nucleotidul formeaza legaturi de H cu capatul 3' al catenei complementare si ofera o grupare 3'OH, cu rol de primer pentru sinteza ADN.
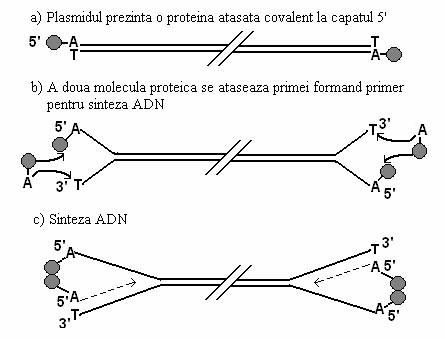
Fig. 110. Replicarea plasmidelor lineare la Streptomyces (dupa Dale, 1996).
Unele dovezi experimentale sugereaza ca replicarea plasmidelor este initiata de proteine de replicare, distincte de cele care initiaza replicarea cromosomului. De exemplu, cloramfenicolul (sau un alt inhibitor al sintezei proteice) inhiba initierea replicarii ADN cromosomal, dar nu a ADN plasmidial. Numarul de plasmide/celula creste la circa l000. Replicarea plasmidelor continua dupa inhibitia replicarii ADN cromosomal, deoarece ele utilizeaza proteine de replicare codificate de plasmide, stabile la actiunea agentilor chimici.
Distributia plasmidelor in procesul diviziunii
Replicarea este o conditie necesara, dar nu suficienta pentru mentinerea plasmidelor in celula bacteriana. Prin diviziune, fiecare dintre cele doua celule fiice trebuie sa dobandeasca cel putin o copie a plasmidei.
Pentru plasmidele mari, cu un numar mic de copii/celula (plasmida F si unele plasmide R), distributia este un proces activ, care implica functii codificate de plasmida. Mecanismul distributiei active este necunoscut. Se presupune ca o secventa de cateva sute de baze, echivalenta unui centromer primitiv, imperecheaza plasmidele inainte de diviziune. Aceasta secventa, impreuna cu una sau mai multe proteine, codificate de plasmida sau de cromosom formeaza sistemul de distributie sau de partitie. Sistemul poate orienta asocierea plasmidelor de membrana sau langa zona de formare a septului de diviziune. Incompatibilitatea functioneaza si la acest nivel: plasmidele asemanatoare au acelasi sistem de distributie si se repartizeaza dezechilibrat in cele doua celule fiice.
Plasmidele mici nu au sisteme de distributie activa. Distributia lor in celulele fiice este intamplatoare. Ele realizeaza un numar mare de copii/celula (30-40), astfel incat sansa ca o celula fiica, sa nu primeasca nici o copie plasmidiala, este foarte mica.
Recombinarea plasmidelor
Recombinarea este definita de proprietatea plasmidelor de a se asocia prin mecanisme genetice, cu alti repliconi: cu cromosomul bacterian sau cu alte plasmide. Recombinarea este o proprietate limitata numai la plasmidele care au capacitate integrativa. Ca urmare a acestei proprietati, ele trec reversibil de la starea fizic autonoma, la cea integrata.
Plasmidele cu functii episomale se pot integra reversibil, atat in cromosomul bacterian, cat si intr-o alta plasmida existenta in celula. Integrarea in structura cromosomului este precedata de sectionarea ambelor structuri genetice, sub actiunea unei nucleaze, urmata de reunirea lor prin extremitatile libere, intr-o singura molecula circulara. Consecutiv recombinarii, plasmida nu se mai replica autonom, ci numai sincron cu ceilalti determinanti genetici cromosomali.
Recombinarea genetica plasmida-plasmida se face prin acelasi mecanism molecular al integrarii plasmida-cromosom si este deosebit de importanta prin consecintele sale. Fenomenul sta la baza formarii plasmidelor mari, cu rol de conjugon, ca si la baza acumularii pe aceiasi plasmida, a unui numar mare de gene structurale, care confera, fiecare in parte, rezistenta la un antibiotic diferit.
Integrarea recombinatorie a plsmidelor este un proces reversibil. Printr-un proces de excizie (invers celui de recombinare), plasmidele se desprind din insertiile structurii genetice in care au fost integrate si revin la starea fizic autonoma. Excizia poate fi corecta, cu exactitate de la situsurile de integrare, astfel incat plasmida redevenita autonoma, este identica cu cea dinainte de integrare. Uneori, excizia este incorecta, caz in care, plasmida schimba cu cromosomul, prin procese de recombinare, o secventa de nucleotide. Plasmida recombinata este modificata: a pierdut o parte din determinantii genetici proprii, dar a castigat o secventa echivalenta, de origine cromosomala.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 3352
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2024 . All rights reserved