| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
ACIZI NUCLEICI - STRUCTURA SI FUNCTII
Acizii nucleici reprezinta moleculele care pastreaza, transporta si manipuleaza informatia in orice celula vie. Codul de scriere a acestei informatii este universal valabil iar mecanismele biochimice implicate sunt pe de o parte complexe si pe de alta parte foarte precise.
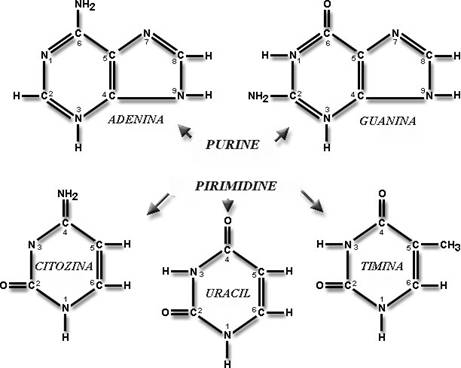
Unitatile de baza din compozitia acizilor nucleici sunt reprezentate de nucleotide - structuri care contin pentoze ( riboza sau deoxiriboza ), acid fosforic si baze azotate cu nucleu purinic sau pirimidinic:
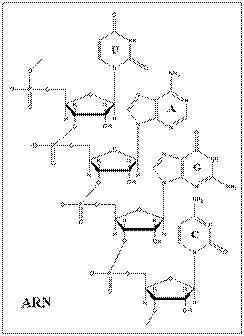
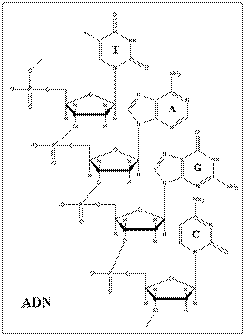
Acizii nucleici pot fi impartiti in 2 mari clase, intre care exista diferente structurale si functionale: acizii ribonucleici (ARN sau RNA) si acidul dezoxiribonucleic (ADN sau DNA).
Ambele macromolecule prezinta din punct de vedere chimic o structura polinucleotidica, construita cu ajutorul legaturilor fosfodiesterice si avand in componenta pentoze facand din ADN si ARN molecule inrudite. Exista insa diferente nete la nivelul structurii:
pentoza din ARN este reprezentata de riboza iar in ADN de deoxiriboza, o riboza in care gruparea hidroxil de la carbonul 2 lipseste fiind, inlocuita cu hidrogen.
ARN are o structura predominant monocatenara iar ADN are o structura bicatenara.
Timina din ADN este inlocuita cu uracilul in molecula de ARN.
|
|
|
ADN-ul, structura polinucleotidica dublu catenara, formeaza genomul unui organism, cu alte cuvinte, totalitatea genelor care poarta informatia acelui sistem biologic. ADN-ul este prezent in nucleul celulelor (si in mitocondrii) sub forma de cromatina sau cromozomi pentru animalele eucariote.Pentru procariote exista un cromozom circular in citoplasma, compactat -prin formarea unor superhelixuri cu ajutorul unei enzime numita ADN giraza. Plasmidele sunt unitati mici de ADN specifice bacteriilor (exemplu: plasmidele care codifica rezistenta la antibiotice).
La eucariote cromatina apare in nucleul celulelor in perioadele de repaus. In momentul inceperii mitozei cromatina se organizeaza in cromozomi, complexe ADN-proteine cu numar diferit de la specie la specie (la om 44xy sau 44xx).
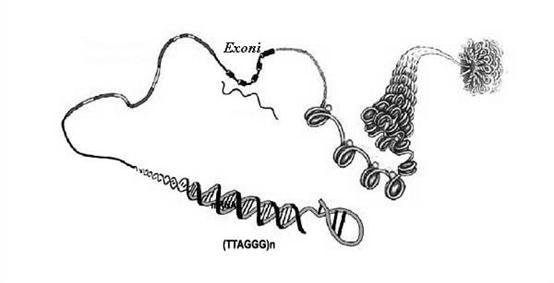
Lungimea de aproximativ 2m a ADN-ului eucariotelor face necesara o supercompactare a acestei molecule care trebuie sa incapa intr-un nucleu cu dimensiuni de ordinul micronilor. Aceasta superhelicare se realizeaza cu ajutorul unor proteine speciale numite histone. Exista 5 clase de histone (H1, H2a, H2b, H3, H4) care realizeaza un miez proteic pe care ADN-ul se infasoara de aproximativ doua ori. Histona H1 nu intra in structura miezului dar are un rol in stabilizarea structurii. Celelalte histone formeaza un octamer(2H2a-2H2b-2H3-2H4) care impreuna cu 2 spire de ADN formeaza un nucleozom. Deci cu ajutorul histonelor ADN-ul eucariot este impachetat de mai multe ori formandu-se o structura super spiralata.
Exista si proteine nehistonice in nucleu, mai mult de 1000 de tipuri, cu functii diverse: diferiti factori de transcriptie, polimeraze, receptori hormonali, etc. Hertonele, de exemplu, au rol in modificarea structurii hipercompactate a ADN-ului in vederea citirii genelor existente la acest nivel.
Nu toate nucleotidele din lantul ADN (numarul lor total fiind de aproximativ 3,5 miliarde de perechi) codifica informatia necesara sintezei de proteine. Exista 3 tipuri de portiuni in aceasta molecula:
Zone inalt repetitive construite din 6-10 perechi de baze ce se repeta de sute de mii de ori.
Zone moderat repetitive
Zone non-repetitive, portiuni corespunzatoare genelor, detinatoare de informatii ce sunt stocate prin insiruirea intr-o anumita ordine a celor 4 litere ale alfabetului genetic: A, T, C, G. La eucariote, o gena apare doar o singura data in tot genomul, exceptie facand doar cateva gene care se pot repeta, de exemplu genele care codifica histonele sau ARN-ul ribozomal.
Tot in cazul eucariotelor, s-a observat in ADN prezenta unor portiuni nepurtatoare de informatie chiar in interiorul genelor. Acestea poarta numele de introni si, in procesul de biosinteza proteica vor fi copiate pe ARN mesager dar vor fi apoi excizate. Doar genele pentru histone si genele procariotelor nu prezinta introni.
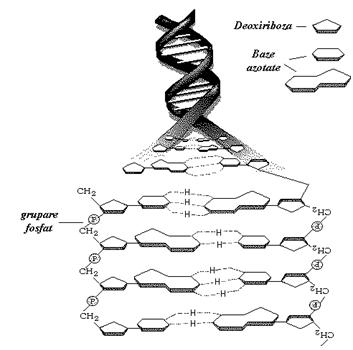
Lantul unei catene de ADN este format prin insiruirea celor 4 tipuri de nucleotide care se unesc intre ele cu ajutorul acidului fosforic. Astfel, se formeaza legaturi fosfodiesterice intre deoxiriboza unui nucleotid si o deoxiriboza a urmatorului, in pozitiile 5' si 3'. Scheletul va fi deci construit dintr-o succesiune deoxiriboza-acid fosforic, de fiecare pentoza fiind legata cate o baza azotata.
Legaturile 5'-3' vor conferi un sens pentru fiecare dintre cele 2 catene care se infasoara una in jurul celeilalte deci se poate vorbi de o catena cu sens 5'-3' si de o catena cu sens invers, 3'-5' numita si catena antisens (catene antiparalele).
S-a demonstrat ca ADN-ul adopta aceasta conformatie dublu helicoidala din ratiuni de stabilitate energetica. Structura este stabilizata astfel prin interactiunea dintre bazele azotate care se plaseaza in interiorul dublului helix. Aici apar legaturi de hidrogen intre bazele de pe o catena si bazele de pe cealalta catena conform unui principiu al complementaritatii. Acest principiu postuleaza ca intotdauna adenina se leaga de timina prin 2 legaturi de hidrogen si guanina se leaga de citozina prin 3 legaturi de hidrogen.
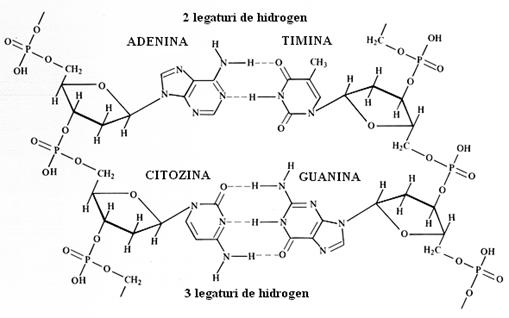
Cele 2 catene pot fi despartite in anumite conditii (de exemplu temperatura si pH) prin desfacerea legaturilor de hidrogen. Acest fenomen se numeste denaturare si este reversibil, prin revenirea la conditiile initiale, procesul numindu-se renaturare. S-a observat ca zonele cu o cantitate mai mare de guanina si citozina sunt mai greu de denaturat din cauza legaturii de hidrogen in plus dintre acestea.
Hibridizarea reprezinta procesul de renaturare a doua catene provenind de la 2 acizi nucleici diferiti, tehnica prin care se poate aprecia de exemplu, gradul de potrivire a acizilor nucleici provenind de la specii diferite.
Exista mai multe posibilitati de structurare a dublului helix (A,B,Z), cea mai raspandita fiind forma B. Aceasta forma se prezinta ca o elice formata din doua lanturi antiparalele, stabilizata prin legaturile de hidrogen din interior. Diametrul este de 20 A, pasul elicei (distanta pentru o rotatie completa de 360) este de 34 A si cuprinde o spira cu 10 perechi de nucleotide. Deci distanta dintre 2 nucleotide de pe acelasi lant este de 3 A. Exista doua zone numite jgeaburi (unul mare si unul mic), importante penru ca aici se poat face un contact mai intim intre diferite substante (medicamente, proteine) si molecula de ADN.
Se poate vorbi deci de mai multe posibilitati de abordare structurala a ADN-ului:
Structura primara, care ne arata insiruirea bazelor azotate pe scheletul format din pentoze unite prin punti fosfodiesterice. (vezi figura 2)
Structura secundara, care ne arata forma de dubla elice stabilizata de puntile de hidrogen dintre bazele azotate complementare.
Structura tertiara, ce explica superhelicarea si compactarea moleculei cu ajutorul proteinelor, fenomen necesar potrivirii cu dimensiunile reduse ale nucleului.
ACIZI NUCLEICI - PROCESE BIOCHIMICE GENETICE
Dogma centrala a geneticii postuleaza urmatoarea transformare: ADN ARN proteine sau O GENA UN ARN MESAGER O PROTEINA. Descoperirile din ultimii ani nu au contrazis transformarile clasice care sunt fundamentul acestei stiinte ci au completat in mod fericit dogma centrala, facand-o mai complexa si mai apropiata de adevar.
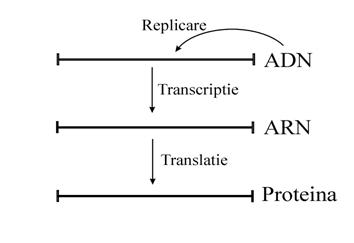
Exista 3 procese esentiale in manipularea informatiei genetice pentru orice celula vie:
Replicarea ADN - reprezinta procesul prin care cantitatea de ADN dintr-o celula se dubleaza si se regaseste practic neschimbata in fiecare dintre celulele fiice.
Transcriptia - reprezinta procesul prin care o parte din informatia de pe ADN este copiata pe ARN (o gena un ARN mesger).
Translatia - reprezinta procesul prin care informatia din ARN este folosita la sinteza unei proteine in ribozomi.
REPLICAREA ADN
Toate celulele se divid in timpul vietii, unele dintre ele sufera mai multe diviziuni si altele se divid de un numar limitat de ori.
In timpul diviziunii celulare informatia prezenta la nivelul ADN-ului se conserva de la o generatie la alta din punct de vedere cantitativ si calitativ. Acest proces care se numeste replicare este semiconservativ deoarece in celulele noi constituite se regaseste o structura a ADN-ului construita dintr-o catena mama si o catena noua, complementara.
Replicarea semiconservativa a fost demonstrata clar la bacterii: administrandu-se ca unica sursa de azot, clorura de amoniu marcata cu azot radioactiv sa observat ca dupa prima diviziune bacteriana azotul radioactiv s-a regasit doar in procent de 50%. La urmatoarea diviziune acest procent scade inca odata la jumatate, s.a.m.d
Replicarea ADN necesita un sistem biochimic si enzimatic complex format din:
ADN matrita (ADN-ul mama)
Deoxinucleotidele necesare sintezei (dATP, dGTP, dCTP, dTTP)
Ribonucleotidele necesare sintezei unui ARN care va folosi ca punct de plecare pentru sinteza ADN (ATP, GTP, CTP, UTP)
Ioni de magneziu
Un sistem multi enzimatic compus din:
a. topoizomeraze - enzime care detensioneaza elicea ADN-ului in timpul replicarii.
b. ADN primaza - enzima care produce mici fragmente de ARN, necesare inceperii procesului de replicare.
c. ADN polimeraza ADN dependenta si care are o serie de caracteristici: sintetizeaza ADN in directia 5`-3`, are si activitate exonucleazica pentru ca elimina unele nucleotide in timpul replicarii, are o inalta fidelitate pentru ca exercita un control strict al replicarii. La procariote existe 3 ADN polimeraze.
d. ADN ligaza - enzima care reuneste fragmentele de ADN rezultate in timpul replicarii.
Intregul complex de factori care participa la acest proces se mai numeste replizom.
Exista diferente in replicarea la procariote si eucariote.
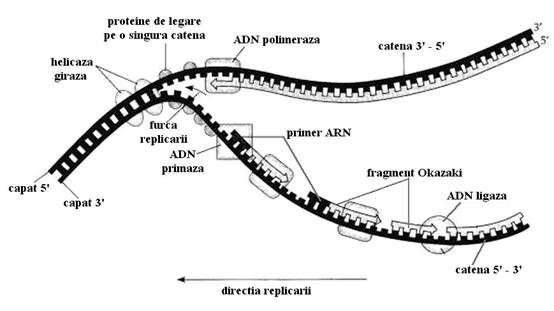
Replicarea la procariote
ADN-ul procariotelor este compus dintr-un singur cromozom circular. Replicarea incepe in puncte bine determinate numite regiuni de origine (ori). La unele bacterii exista o singura regiune de origine (punct de plecare). De aici procesul se desfasoara pa ambele catene ale ADN-ului circular astfel incat la un moment dat un ADN bacterian poate lua forma literei grecesti theta.
![]() Initierea replicarii incepe prin desfacerea legaturilor
de hidrogen dintre nucleotidele de pe cele doua catene ale ADN-ului mama, in
punctul de origine (reamintim ca adenina se leaga de timina prin 2 legaturi de
hidrogen si guanina de citozina prin 3 legaturi de hidrogen, conform
principiului complementaritatii;A=T, G=C). Enzimele
implicate in acest proces se numesc helicaze. Astfel pe
catenele eliberate se poate construi o noua catena de ADN. Deci in fata
enzimelor care realizeza replicarea se afla aceste helicaze care separa cele 2
catene.
Initierea replicarii incepe prin desfacerea legaturilor
de hidrogen dintre nucleotidele de pe cele doua catene ale ADN-ului mama, in
punctul de origine (reamintim ca adenina se leaga de timina prin 2 legaturi de
hidrogen si guanina de citozina prin 3 legaturi de hidrogen, conform
principiului complementaritatii;A=T, G=C). Enzimele
implicate in acest proces se numesc helicaze. Astfel pe
catenele eliberate se poate construi o noua catena de ADN. Deci in fata
enzimelor care realizeza replicarea se afla aceste helicaze care separa cele 2
catene.
Prin separarea celor 2 catene dubla elice a ADN-ului devine din ce in ce mai tensionata. Aceasta situatie ar putea duce la oprirea replicarii daca nu ar exista topoizomerazele care detensioneaza ADN-ul. Topoizomeraza 2 numita si ADN giraza poate induce tensionari negative, inverse tensionarilor produse de helicaza permitand continuarea procesului. Astfel se poate explica actiune antimicrobiana a acidului nalidixic (negram) care inhiba ADN giraza si blocheaza deci diviziunea bacteriana. Topoizomeraza 1 detensioneaza ADN-ul suprahelicat pentru ca poate sa desfaca o legatura fosfodiesterica de pe o catena a ADN-ului permitand relaxarea elicei (catena rupta se roteste in jurul celelalte catene). Dupa detensionare catena desfacuta se poate reface deoarece fenomenul este reversibil, mai mult decat atat neavand nevoie de energie pentru ca aceasta energie este conservata cu ajutorul unui rest de tirozina din enzima.
Odata desfacut si detensionat, pe ADN-ul mama se poate incepe construirea celor 2 catene a noului ADN. Pentru aceasta este nevoie de mici molecule de ARN numite primeri sau initiatori (au 5-10 nucleotide). Acestea sunt realizate cu ajutorul ADN primazei.
De la capatul 3` al ARN-ului initiator pleaca sinteza ADN-ului propriu zis prin adaugarea succesiva a deoxinucleotidelor care se unesc prin legaturi fosfodiesterice. Molecula fiica nou constituita respecta cu strictete ADN-ul matrita astfel incat daca pe ADN-ul mama exista timina acolo va veni adenina, pentru citozina va veni guanina, s.a.m.d. , conform principiului complementaritatii. Enzima necesara este ADN polimeraza III.
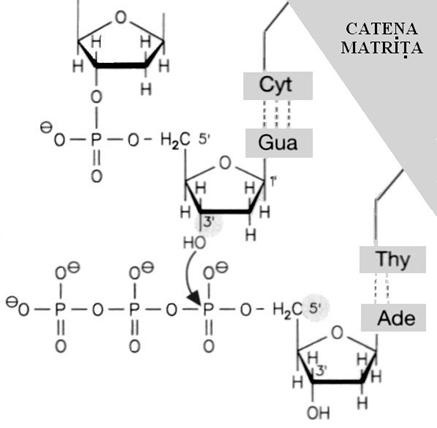
Energia necesara procesului provine din legaturile fosfat tmacroergice ale nucleotidelor folosite. Pirofosfatul rezultat este desfacut in 2 molecule de fosfat impingind reactia ireversibil spre dreapta. Replicarea continua astfel pana la intalnirea unei noi unitati de origine.
Finalul replicarii necesita eliminarea ARN-ului initiator si asigurarea continuitatii catenei nou formate. Enzima necesara este ADN polimeraza 1 care este deci si polimeraza si exonucleaza. Asta inseamna ca ribonucleotidele din ARN-ul initiator sunt inlocuite pe rand cu deoxinucleotidele necesare noului ADN.
Exista proteine specifice legate pe catena ADN, cu rol in reglarea si optimizarea interactiunii dintre catena si enzime.
Acest mecanism explica foarte bine sinteza lantului polinucleotidic cu directia 5`-3`. Insa este demonstrat ca ADN polimeraza nu poate functiona decat in aceasta directie, deci la prima vedere s-ar putea crede ca cealalta catena a ADN-ului mama care are directie inversa nu poate fi folosita in acelasi fel.
S-a demonstrat ca si cealalta catena poate fi copiata cu ajutorul
aceleiasi ADN polimeraze printr-un mecanism asemanator dar care pleaca in sens
invers.
Deci de data asta punctul de plecare a polimerazei este
bifurcatia de replicare construindu-se astfel catena in directia 5`-3`.
Procesul este discontinuu, pentru ca dupa ce punctul
de bifurcatie avanseaza suficient de mult o noua sinteza pleaca din dreptul
noului punct de bifurcatie. Astfel se vor forma mai multe fragmente mici de ADN numite
fragmente Okazaki.Fragmentele Okazaki au o lungime de cateva sute de nucleotide
la eucariote si cateva mii la procariote. Fragmentele vor fi reunite
dupa excizia fiecarui fragment de ARN initiator care insoteste lanturile
In final de pe fiecare catena a ADN-ului mama se construieste cate o catena noua rezultand 2 molecule polideoxinucleotidice bicatenare cu un lant vechi (matrita sau mama) si un lant nou sintetizat de complexul enzimatic numit replicare.
Intregul mecanism este extrem de fidel astfel incat erori nu apar decat cu o frecventa de una la un milion -zece miloane de nucleotide. ADN polimeraza are o extraordinara capacitate de autocontrol astfel incat daca o nucleotida nu este pusa conform principiului complementaritatii aceasta este imiediat eliminata si inlocuita cu nucleotidul corespunzator. Viteza replicarii este de sute de nucleotide pe secunda si se face aproape perfect.
Replicarea la eucariote
Se face asemanator cu cea de la procariote. Exista totusi o serie de particularitati:
ADN polimerazele: aici sunt in numar de trei a b si c a polimeraza este implicata in replicarea ADN-ului nuclear, c polimeraza este implicata in replicarea ADN-ului mitocondrial. Pe de alta parte nici una din enzime nu are activitate exonucleazica.
originea replicarii: la eucariote viteza este mai mica si s-ar parea ca nu exista explicatie cum aprope 2 metri de ADN care contine in jur de 3 miliarde jumatate de baze nucleotidice pot fi copiate doar in aproximativ 8 ore. Dar explicatia este foarte simpla pentru ca exista mai multe origini de replicare, aproximativ una la cateva zeci de mii de baze care functioneaza in acelasi timp. Aici, in momentul inceperii copierii fiecare origine are cate un replizom care se ocupa de ambele catene ale ADN-ului. Fidelitatea este si mai mare, riscul erorii fiind de una la cateva miliarde.
exista un mecanism propus pentru catena 5`-3` a ADN-ului mama in momentul copierii. Se pare ca la nivelul replizomului aceasta catena poate face o bucla care permite folosirea complexului de sinteza in acelasi sens ca in cazul catenei 3`-5`.
biosinteza ADN apare in faza S a diviziunii celulare, intreaga cantitate de ADN si histone (necesare "impachetarii") fiind dublata complet.
ADN ligazele si fragmentele
Diferite situatii (radiatii puternice, toxice, etc.) pot provoca mutatii in timpul replicarii. De exemplu radiatiile ultraviolete pot genera formarea dimerilor intre 2 nucleotide pirimidinice (exemplu timina-timina). Acest defect poate fi inlaturat prin actiunea unor enzime specializate care excizeaza si repara bazele anormale.
Molecula de ADN poate fi sintetizata folosindu-se ca matrita o molecula de ARN. Este o situatie mai rar intalnita, de exemplu in cazul anumitor virusuri ARN. Acestia prezinta o enzima speciala numita revers transcriptaza care este o ADN polimeraza ARN dependenta si are capacitatea de a copia informatia de pe ARN-ul virusului care a patruns in celula pe o molecula de ADN care dupa ce devine dublu catenar se insera in genomul gazdei. Celula astfel parazitata va fabrica "inconstient" proteine si ARN, acesta fiind modul de inmultire a unui virus. Cel mai bun exemplu este virusul HIV care infecteaza in special limfocitele T helper si care poate fi partial inhibat cu medicamente care blocheaza revers transcriptaza.
Revers transcriptaza modifica intr-o mica masura teoria clasica mendeliana a geticii care postuleaza procesul ADN ARN proteine. Intre ADN si ARN informatia poate fi copiata in ambele sensuri. Pe de alta parte revers transcriptaza este utila obtinerii genelor sintetice plecandu-se de la diferiti ARN mesageri, usor de izolat.
Exista o multitudine de baze azotate modificate artificial care pot sa inhibe replicarea normala a AND-ului. Unele dintre acestea pot fi folosite ca medicamente antivirale sau citostatice (ex 5-fluorouracil, 5-bromouracil etc.)
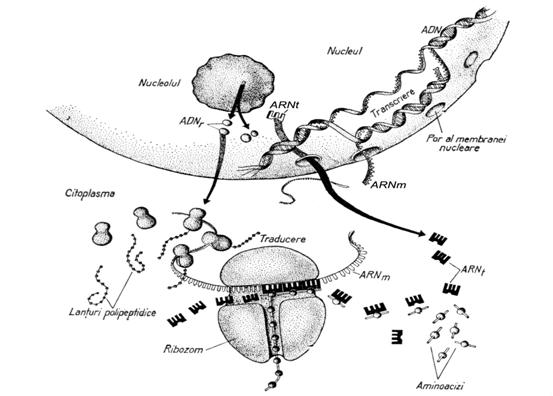
Reprezinta un proces absolut necesar de transfer a informatiei genetice de pe ADN (o portiune a ADN-ului numita gena) pe ARN-ul mesager. Acesta este modalitatea de transport a informatiei care se gaseste in nucleu la ribozomii din citoplasma in vederea sintezei proteinelor.
Exista 3 tipuri importante de ARN:
ARN mesager (ARNm).
ARN de transfer (ARNt).
ARN ribozomal (ARNr).
Deci transcriptia reprezinta sinteza ARN-ului mesager. Structura acestuia este dictata de matrita ADN-ului copiat (gena). Aceasta matrita sau portiune de ADN numita gena se poate afla pe oricare dintre cele 2 catene ale ADN-ului si este transcrisa in functie de necesitatile proteice ale fiecarei proteine.
La procariote exista o singura ARN-polimeraza care catalizeaza formarea tuturor tipurilor de ARN (ribozomal de transfer si mesager).
La eucariote procesul este mai specializat in sensul ca exista 3 tipuri de ARN-polimeraza:
ARN-polimeraza I pentru sinteza ARN ribozomal
ARN-polimeraza II pentru sinteza ARN mesager
ARN-polimeraza III pentru sinteza ARN de transfer
a. Transcrierea informatiei pe ARN necesita la inceput un semnal de initiere. Deci ARN-polimeraza se fixeaza pe o portiune a ADN-ului numita promotor. Acest locus promotor este constituit din aproximativ 40 de perchi de nucleotide specifice inceputului fiecarei gene. Mai exista o secventa de 6 nucleotide situata inaintea celei dintai care mareste exactitatea initierii transcrierii, si un factor sigma care creste afinitatea enzimei pentru promotor.
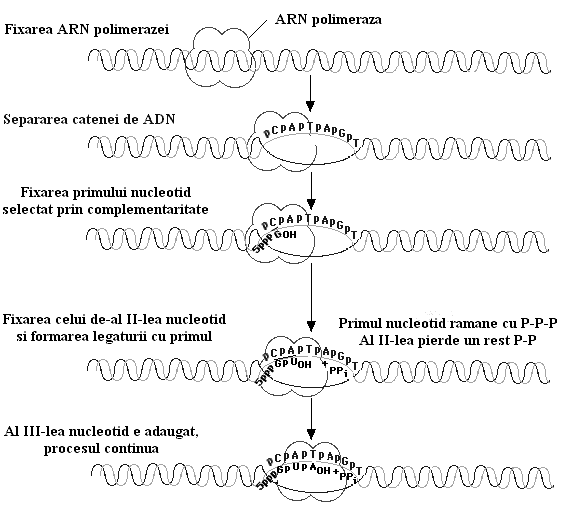
![]() b. Odata fixata, ARN-polimeraza va
incepe elongarea moleculei de ARN folosind ribonucleotide care se ordoneaza
conform matritei ADN-ului ce se copie. Deci gena impune cu ajutorul enzimei aranjarea nucleotidelor
intr-o ordine stricta care respecta principiul complementaritatii (A=U, C=G).
Acum factorul sigma se disociaza de ADN permitand ARN-polimerazaei sa gliseze dea lungul moleculei si sa adauge rand pe rand
ribonucleotidele necesare elongarii moleculei de ARN. Prin atacuri nucleofile
ribonucleotidele se leaga una de alta fiecare la
capatul 3`-OH a celei din urma.
b. Odata fixata, ARN-polimeraza va
incepe elongarea moleculei de ARN folosind ribonucleotide care se ordoneaza
conform matritei ADN-ului ce se copie. Deci gena impune cu ajutorul enzimei aranjarea nucleotidelor
intr-o ordine stricta care respecta principiul complementaritatii (A=U, C=G).
Acum factorul sigma se disociaza de ADN permitand ARN-polimerazaei sa gliseze dea lungul moleculei si sa adauge rand pe rand
ribonucleotidele necesare elongarii moleculei de ARN. Prin atacuri nucleofile
ribonucleotidele se leaga una de alta fiecare la
capatul 3`-OH a celei din urma.
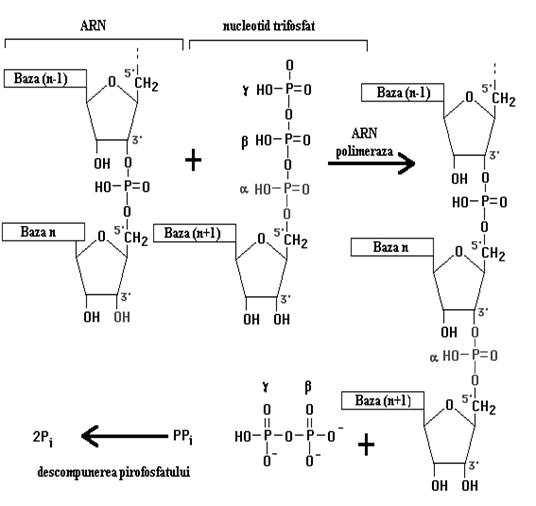
c. Terminarea transcrierii se face cand ARN-polimeraza ajunge la capatul genei unde intalneste un numar mare de nucleotide C si G. Aici, cu ajutorul factorului r aceste semnale sunt recunoscute, enzima paraseste gena matrita si poate sa isi reia functionarea in alta parte prin reasocierea cu factorul sigma.
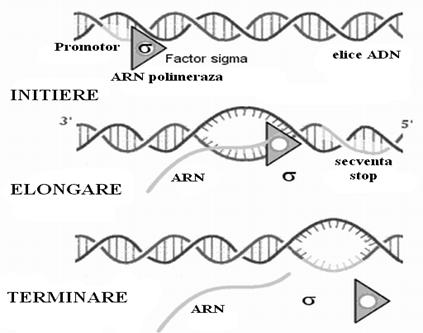
Exista situsuri reglatoare ale transcriptiei situate in fata situsurilor promotoare care supravegheaza frecventa transcrierii. Pe de alta parte promotorii pot imprima viteze diferite procesului ceea ce face ca anumite gene sa fie mai active decat altele.
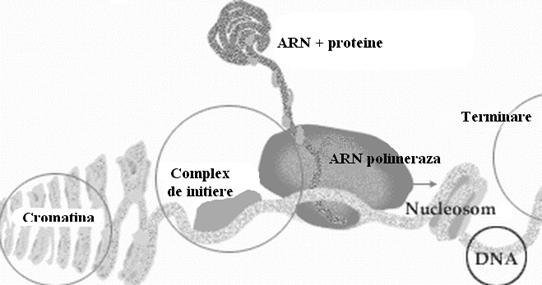
Exista diferite substante care pot bloca transcrierea. Exemple: actinomicina modifica structura ADN-ului matrita si acesta nu mai poate fi copiat; rifampicina, chimioterapic folosit in tratarea TBC-ului si activ pe un spectru larg microbian inhiba ARN-polimeraza bacteriana.
ARN-ul mesager astfel obtinut difera la procariote fata de eucariote. Astfel in ARN-ul procariot molecula este continua deci intreaga informatie de aici poate fi folosita pentru sinteza proteinelor. In ARN-ul eucariot s-a observat prezenta unor portiuni repetitive de nucleotide numite introni care nu detin nici o informatie pentru sinteza proteica. Acesta este motivul pentru care ARN-ul mesager primar este mult mai lung decat ARN-ul mesager folosit de ribozom. Deci intronii trebuie excizati si pastrate doar portiunile cu informatie utila numite exoni.
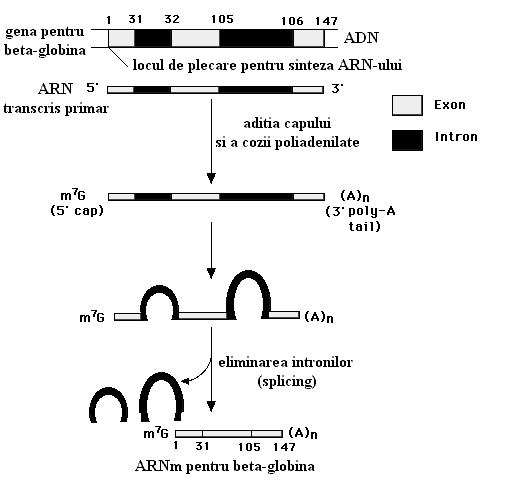
Excizarea intronilor se poate face in 2 feluri:
Cu ajutorul unor enzime speciale care recunosc capetele fiecarui intron (exemplu: ARN U1) acestia sunt eliminati si capetele libere ale exonilor sunt sudate.
Prin autocataliza, fenomen remarcabil care pune pentru prima data in discutie posibilitatea catalitica a unui compus neproteic. Deci chiar ARN-ul ribozomal isi autoelimina intronii printr-un proces care genereaza o bucla (reprezentata de intron), eliminarea acestei bucle cu ajutorul guanozinei si coaserea exonilor.
O astfel de posibilitate surprinzatoare a facut ca primul ARN mesager descoperit cu aceasta insusire sa fie denumit ribozim. S-a reconsiderat viziunea asupra acizilor nucleici care erau vazuti doar ca molecule informationale si care acum sunt priviti si ca enzime.
Rolul intronilor, departe de a fi pe dea intregul cunoscut, se conturreaza in directia reglarii procesului de diferentiere celulara. Mai mult chiar, s-ar parea ca detin controlul transportului ARN-ului mesager din nucleu in citoplasma.
Este posibila biosinteza ARN pe matrita de ARN, de exemplu in cazul unor virusuri. Acestea isi injecteaza materialul genetic in celula gazda si cu ajutorul unei ARN-polimeraze ARN dependenta numita si replicaza reuseste sa isi multiplice molecula informatinala. Procesul este analog unei replicari obisnuite.
Un exemplu clasic de inhibitie a transcriptiei este mecanismul de blocare a ARN polimerazei II de catre amanitina. Aceasta molecula toxica este un octapeptid ciclic produs de Amanita phalloides (buretele viperei) - cea mai toxica ciuperca din aceasta zona geografica. Astfel, o singura ciuperca matura poate sa omoare doi oameni.
TRNSLATIA(BIOSINTEZA PROTEINELOR)
Este un fenomen foarte complex care necsita conlucrarea a peste 300 de enzime si diverse tipuri de acizi ribonucleici. Informatia necesara ordonarii aminoacizilor in proteine (deci constituirea structurii primare) este transportata de pe gena de la nivelul ADN-ului nuclear in citoplasma cu ajutorul ARN-ului mesager. Deci fiecare acid ribonucleic are fuctii specifice:
a. ARN-ul ribozomal intra in structura ribozomului care reprezinta "fabrica" de proteine.
b. ARN-ul de transfer este molecula care transporta fiecare din cei 20 de aminoacizi la locul de sinteza.
c. ARN-ul mesager detine informatia constructiei corecte a proteinei.
a.
ARN-ul ribozomal prin asociere cu proteine si lipide constituie ribozomul sau
corpusculul lui Palade. Ribozomul este construit din 2
subunitati, una mica (30S la procariote si 40S la eucariote) si una mare (50S
la procariote si 60S la eucariote). Astfel ribozomul va
avea
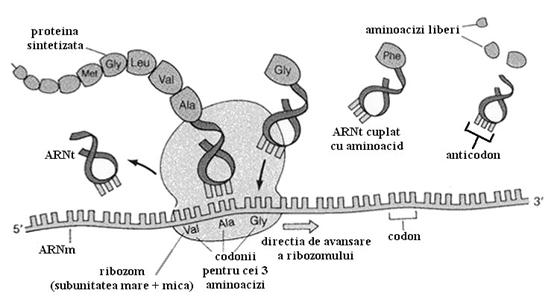
Precum am vazut, ribozomul prezinta 2 situsuri speciale: situsul A care este sediul ARN-ului de transfer ce vine cu aminoacizii necesari sintezei proteice si situsul P ce gazduieste ARN-ul de transfer legat de un lant polipeptidic deja sintetizat.
Procesul de sinteza proteica poate fi schitat sumar prin interactiunea celor 3 tipuri de ARN. Cu alte cuvinte informatia din ARN-ul mesager este citita in ribozom si transpusa in proteine, aminoacizii necesari fiind adusi de ARN-ul de transfer.
Mai multi ribozomi pot citi simultan acelasi ARN mesager pe care il parcurg in acelasi sens. Se constituie astfel un poliribozom, structura ce permite accelerarea sintezei proteice.
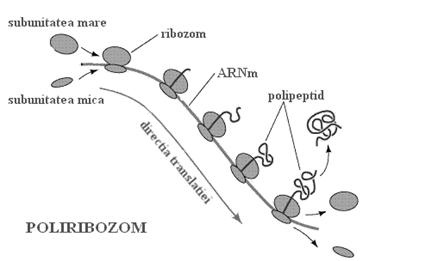
b. ARN-urile de transfer sunt molecule mici cu o forma care poate fi asemanata cu un "trifoi cu patru foi" sau cu o "cruce". La un capat al unuia din brate (capatul 3` terminal) toti ARNt au secventa CCA. Aici se fixeaza unul din cei 20 de aminoacizi. Deci exista cel putin 20 de tipuri de ARNt. In realitate exista mai multe tipuri pentru ca un aminoacid poate fi transportat de diferiti ARNt. O regula este insa sigura: fiecare ARNt ce transporta un aminoacid are la bratul diametral opus o secventa de 3 nucleotide numita anticodon care este strict specifica aminoacidului transportat. Anticodonul trebuie sa se potriveasca perfect prin complementaritate cu codonul corespunzator de pe ARN-ul mesager.
S-a demonstrat ca informatia genetica este foarte bine organizata. Astfel la nivelul ADN-ului, exceptand portiunile care nu codifica nici o informatie si sunt doar zone polinucleotidice repetitive sau moderat repetitive, informatia pentru fiecare proteina este organizata in gene. O gena reprezinta deci un grup mai mare sau mai mic de nucleotide purtatoare de informatie. Un codon reprezinta 3 nucleotide. Fiecare codon codifica un aminoacid. Fiecare gena codifica o proteina. Deci ARN-ul mesager copie toti codonii unei gene transportand in acest fel informatia pentru fiecare aminoacid.
La nivelul ribozomului fiecare codon se potriveste cu anticodonul complementar de pe ARNt si aceasta este modalitatea prin care aminoaciziii sunt adusi in ordine, uniti prin legaturi peptidice si transformati in proteine.
Se poate face o comparatie intre modul de stocare a informatiei in meomoria calculatoarelor si acizii nucleici. Pentru calculatoare limbajul primar este in baza 2 (binar), pentru acizii nucleici limbajul este in baza 4 (patru nucleotide) si este transferat proteinelor ca limbaj in baza 20 (20 de aminoacizi). Codul genetic reprezinta "alfabetul" necesar sintezei de proteine si ne arata care codoni (3 nucleotide) corespund fiecaruia dintre cei 20 de aminoacizi.
Biosinteza proteinelor (translatia) se realizeaza in 5 etape:
Activarea aminoacizilor.
Initierea lantului polipeptidic.
Elongarea lantului polipeptidic.
Terminarea lantului polipeptidic si eliberarea acestuia.
Prelucrari post traducere ale proteinei sintetizate.
Activarea aminoacizilor este necesara inceperii biosintezei si poate fi explicata in 2 etape:
In prima etapa aminoacidul reactioneaza cu ATP rezultand un complex aminoacil-AMP. Pirofosfatul eliberat este hidrolizat si in acest fel reactia este impinsa spre dreapta, fiind ireversibila.
In a doua etapa complexul cedeaza aminoacidul unei molecule de ARNt, specifica acelui aminoacid (deoarece anticodonul ARN-ului corespunde codonului de pe ARN-ul mesager care codifica aminoacidul respectiv).
Precum se observa, activarea se face cu consum de energie, enzimele implicate sunt ligazele (aminoacil-ARNt sintetaze) care au o specificitate absoluta. Aminoacidul este legat deci de carausul sau specific la capatul CCA al acestuia, la gruparea OH din pozitia 2 sau 3 de pe riboza restului adenilic. Enzima poate recunoaste daca un aminoacid gresit s-a fixat pe ARNt, situatie in care il elimina si il inlocuieste cu aminoacidul corespunzator deoarece prezinta si un locus hidrolitic.
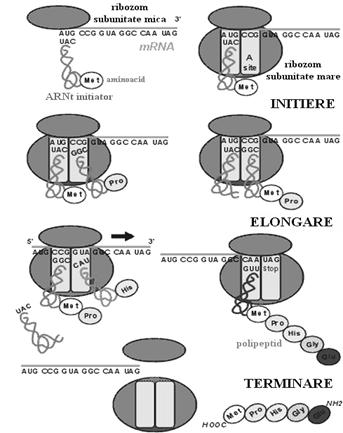
Initierea lantului polipeptidic incepe prin legarea ARN-ului mesager de subunitatea mica a ribozomului. Reasocierea prematura a celor 2 subunitati ribozomale este prevenita de factorul de initiere 3 (IF3). Legarea se face la codonul aminoacidului amino-terminal, aminoacid de initiere care este formil-metionina sau metionina, deci codonul AUG sau GUG este codonul de initiere. Se consuma energie generata de GTP.
Unele molecule de ARN mesager pot codifica mai multe proteine pe acelasi lant, deci asta inseamna ca vor fi prezenti mai multi codoni de initiere pe lantul acestui tip de ARN. Exista o secventa polinucleotidica bogata in purine situata in fata codonului de initiere care mareste afinitatea subunitatii mici ribozomale pentru acesta.
Urmeaza legarea primului aminoacid care reste bineinteles formil-metionina. Acesta este adus de ARN-ul de transfer specific si pozitionat in situsul p ribozomal in dreptul codonului corespunzator prin potrivirea codon-anticodon(deci anticodonul ARN-ului de transfer se potriveste numai cu codonul initiator al ARN-ului mesager). Etapa necesita factorii de initiere 1 si 2(IF1, IF2).
Ultima etapa a procesului de initiere implica legarea subunitatii ribozomale mari care reintregeste ribozomul. S-a format acum un complex intre ribozom, ARN mesager si primul ARN de transfer. Constructia proteinei va necesita mai departe miscarea lantului ARNm in ribozom care este citit, si informatia va servi sintezei proteinei.
Elongarea lantului polipeptidic se face prin adaugarea succesiva a aminoacizilor necesari sintezei. Fiecare dintre acestia este adus de ARN-ul sau de transfer specific si pozitionat corect prin potrivirea codon-anticodon intre ARN-ul mesager si ARN-ul de transfer. Este nevoie de factorul de elongare si energie sub forma de GTP. Intregul mecanism consta de fapt in miscarea ARN-ului mesager in ribozom care isi expune codonii rand pe rand pentru a fi cititi. "Citirea" reprezinta contactul intre acesti codoni si anticodonii ARN-urilor de transfer, care vin rand pe rand cu aminoacizii ce vor constitui proteina. Intregul proces se desfasoara deci in ribozom, la nivelul situsurilor p si a. ARNt vine cu aminoacidul 2 in situsul a, aminoacidul initiator se leaga de al doilea formandu-se un dipeptid pozitionat deci in situsul a, ARNt ramas liber pleaca din situsul p si ARNt-dipeptidul se muta in situsul p prin miscarea ARN-ului mesager in ribozom.
Procesul continua in aceeasi maniera pana la elongarea completa a proteinei. Deci, spre exemplu, aminoacidul 3 vine in situsul a, se formeaza legatura peptidica intre aminoacizii 2 si 3, ARN-ul mesager se muta in ribozom astfel incat complexul ARNt-tripeptid se va muta la randul lui din situsul a in situsul p, s.a.m.d.
Miscarea ARN-ului mesager in ribozom consuma GTP si este posibila datorita unei translocaze. Aminoacidul initiator (formil-metionina) va fi inlaturat din lantul polipeptidic deorece nu are decat un rol de start al sintezei proteice.
Pentru realizarea legaturii peptidice intre aminoacizi este nevoie de o enzima numita peptidiltransferaza, un factor de elongare G si consum de energie.
Toxina difterica, molecula foarte toxica fabricate de Corynebacterium diphtheriae - microorganismul care provoaca difteria - blocheaza sinteza proteica prin inhibarea unui singur factor de elongare. Astfel, o singura molecula poate distruge o celula.
Terminarea sintezei se face cand pe ARN-ul mesager ce trece prin ribozom apare un codon care nu codifica nici un aminoacid. Acesta se mai numeste codon stop sau nonsens si nu exista un ARN de transfer cu anticodonul corespunzator. In acest moment polipeptidul sintetizat se desface de ARNt, acest ultim ARNt este eliberat din ribozom si ribozomul se desface din nou in cele doua subunitati fiind pregatit pentru o noua sinteza. ARNm dupa copiere este eliminat prin hidroliza.
Sinteza de proteine necesita deci un complex de factori, o mare cantitate de energie si se desfasoara foarte rapid (100 de aminoacizi pot fi legati in 5 secunde).
La eucariote aminoacidul initiator este metionina, si viteza de sinteza este mult mai mare decat la procariote.
Prelucrarile postraducere ale proteinelor sunt necesare definitivarii sintezei si transformarii acestor molecule in compusi functionali. In aceasta ultima etapa se va elimina formil-metionina initiatoare, se hidroxileaza unii aminoacizi (prolina, lizina), unlele proteine se pot iodura (tireoglobulina), altele se pot combina cu molecule neproteice (fosfoproteine, glicoproteine), altele se pot desface in fragmente mai mici (preproinsulina).
Biosinteza proteinelor dispune de mecanisme exacte de reglare. Cel mai acceptat este cel descris de Jacob si Monod care au dezvoltat teoria operonului. S-ar parea ca exista inductori si represori ai biosintezei care mentin o balanta a procesului.
Inhibitori ai sintezei proteice
Sunt in general antibiotice, compusi care pot astfel sa blocheze activitatea celulara la procariote.
|
Inhibitorul |
Procesul inhibat |
Mecanismul de actiune |
|
Mitomicina |
Replicare |
Impiedica separarea catenelor de ADN |
|
Acid nalidixic |
Replicare |
Inhiba ADN giraza |
|
Rifampicina |
Transcriere |
Inhiba ARN polimeraza |
|
Actinomicina |
Transcriere |
Se fixeaza pe ADN |
|
Streptomicina |
Translatie |
Inhiba legarea ARNt initiator la subunitatea 30S |
|
Cloramfenicol |
Translatie |
Inhiba peptidil transferaza |
|
Tetraciclina |
Translatie |
Inhiba legarea ARNt la ribozomi |
|
Eritromicina,Azitromicina |
Translatie |
Blocheaza subunitatea 50S |
|
Puromicina |
Translatie |
Blocheaza elongarea inhiband competitiv ARNt |
|
Kanamicina |
Translatie |
Inhiba citirea codului genetic |
Sinteza proteica - vedere de ansamblu: (dupa Gray's anatomy)
Se pot da exemple de substante care interfera sinteza proteica in celulele eucariote. Acestea sunt toxinele proteice produse de bacterii (exotoxine termolabile), heteropolimeri compusi in general din 2 fragmente: un fragment A cu rol determinant in actiunea toxica a toxinelor si fragmente B cu rol de liganzi la diferite tipuri de receptori specifici ai celulelor eucariote. Fragmentul A poate sa catalizeze transferul ADP-ribozei din molecula de NAD pe diverse molecule tinta implicate in sinteza proteica. Astfel functia acestora va fi modificata si apare efectul toxic ca in urmatoarele exemple:
|
Toxina (bacteria) |
Mecanisme de actiune (acceptori de ADP-riboza) |
Efecte biologice sau clinice |
|
Difterica (C. Diphtheriae) |
Factorul de elongare 2 (EF-2) |
Stopeaza sinteza proteica: efecte necrotice, miocardo- si neurotoxice |
|
Toxina A (Bacilul pioceanic) |
EF-2 |
Stopeaza sinteza proteica: efecte necrotice |
|
Toxina Shiga (S. dysenteriae) |
Fractiunea ribozomala 60S si factorul EF-1 |
Stopeaza sinteza proteica: efecte enteroneurotoxice |
Capitol realizat de Bogdan Stoica. S-a folosit:
Biochimie medicala - Aurora Popescu, 2002
Imagini de la diferite universitati de pe Internet ( unele imagini ar putea avea drept de copyright ; imaginile sunt folosite in scop necomercial)
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 5607
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved