| CATEGORII DOCUMENTE |
| Agricultura | Asigurari | Comert | Confectii | Contabilitate | Contracte |
| Economie | Transporturi | Turism | Zootehnie |
Microbiologia branzeturilor
Nevoia obtinerii culturilor starter a aparut odata cu observarea faptului ca se realizeaza o fermentatie mai buna la fabricarea branzeturilor, daca in laptele proaspat se adauga zerul din ziua precedenta. La inceput transferul culturii se realiza prin insamantari pe zer, si astfel era propagata o anumita cultura de microorganisme. Deoarece cultura transferata era folosita pentru a incepe fermentatia si a imbunatati activitatea culturii existente ea a fost denumita cultura starter.
La inceputul secolului XX oamenii de stiinta au inceput studierea microorganismelor implicate in fabricarea branzeturilor. Asa cum era de asteptat in afara de bacterii lactice s-a gasit si un numar mare de contaminati. Culturile au fost purificate progresiv si mentinute astfel prin insamantari in medii sterile. Din aceste culturi cele mai active au fost selectionate si comercializate sub forma de culturi starter comerciale.
Microorganismele adaugate la fabricarea branzeturilor pot fi impartite in doua mari grupe microorganisme starter si microflora secundara. Pe langa acestea in timpul fabricatiei se mai pot dezvolta in branza si microorganisme non-starter provenite din lapte, sau din contaminari in timpul prelucrarii.
Microflora starter este formata cu precadere din bacterii ce produc fermentatia lactica, proces anarob de metabolizare a glucidelor sub actiunea echipamentului enzimatic al microorganismelor, avand ca produs principal acidul lactic.
Microorganismele din cultura secundara nu contribuie la formarea acidului lactic, dar in general joaca un rol important in timpul maturarii.
Bacteriile lactice intalnite frecvent in culturile starter sunt: Lactoccocus lactis, Streptococcus therpmophillus, Lactobacillus helveticus, Lactobacillus delbrueckii, utilizate fie individual, fie in combinatii in functie de sortimentul de branza.
O serie de microorganisme cunoscute sub denumirea de microflora secundara, pot fi adaugate alaturi de cultura starter. Din aceasta categorie fac parte porpionibacterii, brevibacterii si specii de mucegaiuri (Penicillim). Aceste microorganisme sunt utilizate impreuna cu bacteriile lactice si sunt specifice unui anumit sortiment de branzeturi (de exemplu in branza Elvetiana gaurile specifice se datoraeza propionibacteriilor, culoarea galbena si aroma tipica branzei Brick se datoreaza lui Brevibacterium linens).
Producerea de acid lactic de catre microflora starter in timpul fabricarii branzeturilor determina o reducere a pH-ului laptelui care combinata cu tratamentul termic si amestecarea promoveaza sinereza prin expulzarea zerului din cas (Walstra, 1993) contribuind semnificativ la unul din procesele de baza ale fabricarii branzeturilor.
Maturarea branzeturilor este un proces complex care implica un numar de reactii biochimice la care iau parte si concentratii mari de microorganisme prezente in branza (Cogan, 2000). Atat continutul de acid lactic cat si procentul de transformare al lactozei in acid lactic reprezinta doi indicatori foarte importanti pentru evaluarea calitatii branzeturilor. In timpul maturarii, culturile de bacterii sufera procesul de autoliza, eliberand enzime intracelulare in reteaua de branza, iar aceste enzime vor continua sa actioneze asupra componentilor casului, dezvoltand aroma si producand modificarile dorite de textura in timpul maturarii.
Bacteriile lactice (BL)
Bacteriile producatoare de acid lactic (BL) indeplinesc mai multe functii:
T produc acid lactic si contribuie la coagularea laptelui;
T dau taria coagulului si influenteaza formarea branzeturilor;
T dezvoltarea acidului determina prezenta cantitatilor reziduale de coagulant, care afecteaza maturarea; mai mult acid in cas inseamna mai mult coagulant rezidual legat
T nivelul formarii acidului influenteaza disocierea fosfatului de calciu coloidal, care la randul sau influenteaza proteoliza in timpul maturarii si determina proprietatile reologice ale branzeturilor;
T formarea acidului si producerea altor substante antimicrobiene controleaza cresterea bacteriilor patogene in branzeturi;
T formarea acidului lactic contribuie la proteoliza si la formarea aromei in branza;
T cresterea bacteriilor producatoare de acid lactic produce scaderea potentialului redox (Eh), necesar pentru producerea si reducerea compusilor cu sulf (metantiolul), care contribuie la formarea aromei branzei Cheddar.
Caractere morfologice: bacteriile lactice prezinta heterogenitate morfologica. Principalele forme sunt derivate de la forma coccus si se gasesc sub forma de streptococi (g. Lactococcus si g. Streptococcus), de diplococi (g. Leuconostoc), de tetrade (g. Pediococcus), iar numeroase bacterii lactice se prezinta sub forma de bastonase (g. Lactobacillus).
Caractere fiziologice: BL folosesc ca sursa de carbon si energie pentozele (riboza, xiloza, arabinoza), hexozele (glucoza, galactoza) si diglucidele (lactoza, maltoza, zaharoza). Dintre acizi, acidul malic poate fi transformat in acid lactic, iar acidul citric in acetoina si diacetil.
Acidul lactic nu poate fi folosit de BL si acumularea sa prin fermentare conduce la inhibarea inmultirii si incetinirea vitezei de fermentatie. Ca surse de azot BL folosesc aminoacizii, peptidele si amidele. Echipamentul enzimatic intracelular proteolitic al BL contribuie semnificativ la maturare dupa autoliza celulelor microbiene.
BL necesita ca factori de crestere vitaminele B1, B6, biotina, acidul para-aminobenzoic, acidul folic si acidul pantotenic.
Acidotoleranta BL este exprimata de domeniul de pH in care are loc cresterea 4,3-4,8 pentru lactococi si 3,8-4,4 pentru lactobacili.
In raport cu oxigenul din aer bacteriile lactice subt anaerobe sau facultativ anaerobe, iar in ceea ce priveste domeniul de temperatura BL sunt adaptate sa creasca intr-un domeniu larg de temperaturi (0-55 0C) cu valori optime:
mezofile
domeniu de crestere 15-40 0C
- t opt 25-30 0C pentru streptobacterii
- t opt 30-35 0C pentru lactococi
termofile
domeniu de crestere 30-50 0C
- t opt 45-50 0C pentru lactobacterii
Culturile mezofile au temperatura optima de crestere in jurul valorii de 300C si sunt utilizate in branzeturi care nu sunt tratate la mai mult de 400C. Caracteristicile culturilor mezofile starter sunt prezentate in tabelul 1.
Branzeturile care se prelucreaza la temperaturi de 50-56oC utilizeaza culturi termofile.
Lactococcus lactis ssp. lactis
O parte din tulpinile izolate din laptele crud produc o bacteriocina denumita nizina. Nizina cunoscuta si numele de lantibiotic (contine lantionina Ala-S-Ala si β-metilantionina) este termostabila si este solubila la pH scazut, ceea ce reduce potentialul sau de utilizare. Producerea sa este legata de pretenta unei plasmide din gama de la 28 la 30 MDa. Plasmida este responsabila pentru abilitatea de a fermenta zaharoza si rezistenta nizinei.
Nizina este activa impotriva sporilor de Clostridium botulinum si a altor cateva bacterii G (+) cum sunt Bacillus, Listeria si Streptococcus. Multe tulpini de Lactococcus lactis ssp. lactis din laptele crud produc un miros "amar". Aceste tulpini metabolizeaza leucina si produc 3-metilbutanolul, care este un compus nedorit si o cantitate de 0,5 ppm este suficienta pentru a produce defectul de amareala.
Lactococcus lactis ssp. cremoris
Originea acestei bacterii nu este cunoscuta si ea nu a putu fi izolata din laptele crud. O serie de tulpini produc o gama restransa de bacteriocine, denumite diplocine. Aceste microorganisme nu cresc la 400C si sunt mult mai sensibile la prezenta sarii. Multe culturi comerciale contin predominant tulpini din aceasta specie.
Amestecuri de lactococi sunt utilizate drept culturi starter pentru ranza Cheddar, Colby si Cottage, atunci cand producerea gazului si a texturii deschise nu sunt de dorit.
Lactococcus lactis ssp. lactis var. diacetylactis este utilizata in combinatie cu alte culturi starter pentru producerea branzeturilor cu mucegai in pasta, maturate: Edam, Gouda si branzeturile crema. Aceste microorganisme sunt capabile sa produca CO2, diacetil, acetoina si acetati din citratii prezenti in lapte.
Tabelul 1. Proprietatile culturilor starter mezofile
|
Denumire veche |
Streptococcus lactis |
Streptococcus cremoris |
Streptococcus diacetylactis |
Leuconostoc lactis |
Leuconostoc cremoris |
|
Denumire noua |
Lactococcus lactis ssp. lactis |
Lactococcus lactis ssp. cremoris |
Lactococcus lactis ssp. lactis bvar. diacetylactis |
Leuconostoc lactis |
Leuconostoc mesenteroides ssp. cremoris |
|
To optima Crestere la 10oC Crestere la 40oC Crestere la 45oC Supravietuire la 72oC/15sec Crestere in NaCl 2% Crestere in NaCl 4% Crestere in NaCl 6,5% Producere de NH3 din arginina Metabolizare citratului Producere CO2 Izomer al acidului lactic produs % acid lactic in lapte Producere de bacteriocine Lactoza Glucoza Galactoza |
30oC L Nizinaa |
30oC L Diplocinaa |
30oC L |
30oC D |
30oC D +w |
a- nu toate tulpinile produc bacteriocine; + pozitive; -negative
Genul Leuconostoc include bacterii heterofermentative sub forma de coci, diplococi, care produc prin fermentatie: acid lactic D(-), alcool etilic si CO2. Bacteriile din g. Leuconostoc se gasesc in culturile starter si sunt considerate importante in formarea aromei, datorita abilitatii de a descompune citratii si formarea diacetilului din piruvatul produs. Leuconostocii sunt mai putin activi decat Lactococcus lactis ssp. lactis var. diacetylactis, atacand citratul doar in mediu acid. Leuconostoc reprezinta max. 5-10% din cultura starter. Atunci cand in cultura de lactococi se adauga Leuconostoc pentru producerea aromei, cultura mixta este de tip B sau L, iar cand componentul de aroma este Lactococcus lactis ssp. lactis var. diacetylactis cultura este de tip D. Culturile de tip BD sau DL contin atat Leuconostoc cat si Lactococcus lactis ssp. lactis var. diacetylactis. Lactococii fara aroma sunt N sau O. Leuconostoc ssp. se diferentiaza de alte bacterii starter prin capacitatea de a metaboliza zaharurile pe calea fosfocetolazei si prin capacitatea redusa de a creste in lapte. Aceasta se datoreaza, probabil lipsei unui sistem de proteinaze, care sa degradeze proteinele laptelui si sa permita cresterea. Aceasta problema poate fi depasita prin adaugarea a 0,3 (w/v) extract de drojdie ca stimulent, iar daca lactoza nu poate fi catabolizata, se poate adauga glucoza sa fie necesara (1% w/v).
Streptococcus salivarius ssp. thermophillus microorganisme G(+), anaerobe, sub forma de streptococi care sunt utilizate la fabricarea iaurtului si a branzei Mozzarella. Streptococcus nu creste la temperaturi de 10oC dar se dezvolta bine intre 40-45oC, iar majoritatea tulpinilor poate supravietui la 60oC timp de 30 minute. Bacteria este foarte sensibila la prezenta antibioticelor. Penicilina (0,05 UI/ml) poate interfera si bloca procesul de acidifiere a laptelui. Streptococcus salivarius ssp. thermophillus creste foarte bine in lapte si fermenteaza lactoza si zaharoza. O concentratie de 2% NaCl poate preveni dezvoltarea multor tulpini. Acesti streptococi poseda un sistem proteolitic slab, fiind adesea combinati cu lactobacilii proteolitici in cultura starter. Majoritatea streptococilor cresc mai rapid in lapte decat lactobacilii si produc acid mai repede. Tulpinile de streptococi poseda β-galactozidaza si utilizeaza numai galactoza din lactoza, lasand glucoza in mediu. Un studiu recent care a urmarit activitatea proteolitica a tulpinilor de Streptococcus salivarius ssp. thermophillus si a noua tulpini de Lactobacillus delbrueckii ssp. bulgaricus a aratat ca lactobacilii au prezentat activitate proteolitica intensa (14,6-61,0 μg tirozina/ml lapte) si Streptococcus salivarius ssp. thermophillus a prezentat o activitate proteolitica mai redusa (2,4-14,8 μg tirozina/ml lapte).
Culturile in amestec, cu exceptia unei singure combinatii, au eliberat mai multa tirozina (92,6-419,9 μg tirozina/ml lapte) decat suma cantitatilor produse de culturile individuale. Amestecurile de culturi produc de asemenea mai mult acid, scazand pH-ul. Din 81 de combinatii ale culturilor de Streptococcus salivarius ssp. thermophillus si Lactobacillus delbrueckii ssp. bulgaricus numai o combinatie s-a dovedit mai putin activa in ceea ce priveste proteoliza (92,6 μg tirozina/ml lapte) decat cantitatea de tirozina produsa de Lactobacillus delbrueckii ssp. bulgaricus (125 μg tirozina/ml lapte).
Lactobacillus microorganisme G(+), catalazo-negative, anaerobe si acidotolerante (tabelul 2). Lactobacillus delbrueckii ssp. bulgaricus, Lactobacillus delbrueckii ssp. lactis sunt microorganisme homofermentative termofile care produc numai acid lactic si cantitati minore de substante de aroma si sunt utilizate in combinatie cu Streptococcus salivarius ssp. thermophillus drept culturi starter pentru branzeturile Elvetiene, Parmesan si Mozzarella. Lactobacillus helveticus si Lb. acidophillus sunt microorganisme homofermentative mezofile care prezinta topt = 30-35 0C.
In genul Lactobacillus exita si tulpini de lactobacili heterofermentativi mezofili care produc acid lactic, acid acetic, diacetil si CO2: Lb. plantarum, Lb. casei si heterofermentativi termofili: Lb. brevis, Lb. fermenti, Lb. bifidum s.a. Proprietatile tulpinilor sunt prezentate in tabelul 2. Premi s.a., in 1972 au izolat tulpini si au gasit ca β-galactozidaza este enzima dominanta in Lactobacillus helveticus, Lactobacillus delbrueckii ssp. lactis si Lactobacillus delbrueckii ssp. bulgaricus. Lactobacillus casei nu poseda activitate β-galactozidazica. Acest tip de fermentatie influenteaza calitatea branzeturilor. Culturile cu activitate β-galactozidazica utilizeaza jumatate din cantitatea de lactoza, reprezentata de glucoza si elibereaza galactoza in mediu. Un exces de galactoza in branza Mozzarella poate cauza brunificarea branzei pentru pizza, iar galactoza poate servi ca sursa de energie pentru fermentatii nedorite produse de microorganismele rezidente din branzeturi. Se recomanda ca Lb. helveticus, care poate fermenta galactoza sa fie utilizat impreuna cu Streptococcus salivarius ssp. thermophillus.
O relatie de simbioza exista intre Lactobacillus delbrueckii ssp. bulgaricus si Streptococcus salivarius ssp. thermophillus. Intr-o cultura in amestec, cresterea asociata a bastonaselor si a cocilor duce la o producere mai mare de acid si o dezvoltare a aromei mai buna decat prin utilizarea culturilor singulare.
S-a stabilit ca numerosi aminoacizi eliberati de proteazele din Lactobacillus delbrueckii ssp. bulgaricus prin actiunea asupra cazeinei stimuleaza cresterea lui Streptococcus salivarius ssp. thermophillus. In schimb Streptococcus salivarius ssp. thermophillus produce CO2 si acid formic care stimuleaza cresterea lui Lactobacillus delbrueckii ssp. bulgaricus. In timpul primei parti a incubarii, Streptococcus salivarius ssp. thermophillus creste repede, indeparteaza excesul de oxigen si produce stimulentii amintiti. Ulterior, cresterea lui Streptococcus salivarius ssp. thermophillus incetineste datorita acumularii acidului lactic, iar cresterea aciditatii permite dezvoltarea lui Lactobacillus delbrueckii ssp. bulgaricus, care este mult mai acidotolerant (fig. 1).
In branza Emmental densitatea maxima a streptococilor lactici 109 ufc/g la periferie si 108 ufc/g in mijloc este atinsa dupa aproximativ 3 ore de presare. Acidul lactic si formic produs de streptococi stimuleaza stimuleaza cresterea lactobacililor pana la atingerea densitatii de aprox. 109 ufc/g dupa 10-20 ore de presare. Populatia de lactobacili si streptococi scade apoi in timpul presarii si sararii. Pentru utilizarea unui inocul cu 1:1 raportul dintre coci si bastonase, cantitatea de inocul, timpul si temperatura de incubare trebuie controlate si tancul starter trebuie racit rapid. Multe tulpini de Lactobacillus delbrueckii ssp. bulgaricus continua sa produca acid la temperatura ridicata.
Numerosi lactobacili au fost identificati in timpul maturarii branzeturilor. Lactobacilii ocupa o nisa specifica in maturarea branzeturilor. Lactobacilii au fost izolati din branzeturi si supusi identificarii, iar cele mai raspandite sunt subspecii gasite au fost Lactobacillus casei, Lb. fermentum, Lb. brevis.
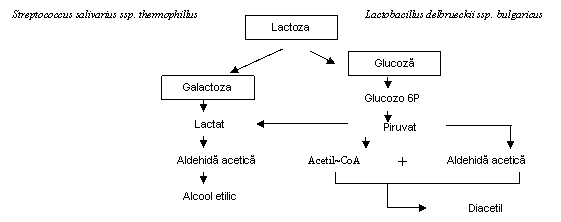
Fig. 1 Schema de metabolizare a lactozei de cultura starter
Prezenta microorganismelor heterofermentative - Lb. fermentum, Lb. brevis (> 106 ufc/g) a determinat defecte de textura la branza Cheddar. Adaugarea lactobacililor homofermentativi determina imbunatatirea calitatii branzeturilor in special prin accelerarea proceselor de maturare.
Tabelul 2. Caracteristicile lactobacililor utilizati la fabricarea branzeturilor
|
bacterii |
% acid lactic in lapte |
Izomer al acidului lactic |
Crestere |
Sensibilitate la sare |
Glu |
Gal |
Lac |
NH3 din arginina |
Bacterio cine |
||
|
15oC |
45oC |
50oC |
|||||||||
|
Lactobacillus delbrueckii ssp. bulgaricus Lactobacillus delbrueckii ssp. lactis Lactobacillus helveticus Lactobacillus casei ssp. casei Lactobacillus casei ssp. pseudoplantarum Lactobacillus casei ssp. rhamnosus Lactobacillus platarum Lactobacillus curvatus Lactobacillus fermentumA Lactobacillus brevisA Lactobacillus buchneriA Lactobacillus bifermentnasA, B |
|
D D DL L DL L DL DL DL DL DL DL |
C |
<2% <2% <2% 8%+a, 10%-a 6%+a, 8%-a 8%+a, 10%-a 6%+a, 8%-a 8%+a, 10%-a 4%+a, 6%-a | |||||||
a - necunoscuta /creste in mediu cu sare/4 zile la 35oC;
+ creste;
- nu creste;
A - produce gaz in branzeturi;
B - fermentatie lactica in branzeturi cu producere de CO2, etanol si acid acetic;
C - poate creste in branzeturi la 15oC.
Cultura secundara
Propionibacterium - bacterii G(+), catalazo-pozitive, anaerobe/facultativ aerobe, nesporulate. Patru specii sunt asociate cu branza Elvetiana: Propionibacterium freundenreichii, P. jensenii, P. thoenii si P. acidipropionici. Fermentarea conduce la producerea de acid propionic, acid acetic si CO2. Aceste microorganisme pot tolera temperaturi de 52 0C.
3 C6H12O6 → 2 CH3-COOH + 4 CH3-CH2-COOH + 2 CO2 + 2 H2O
P. thoenii si si P. acidipropionici pot produce pigmentarea nedorita in rosu, maro si galben-portocaliu. Cresterea propionibacteriilor este indusa atunci cand branza este transferata intr-o camera calda ( 20-25 0C si 80-85 % umiditate relativa). Aici densitatea celulelor bacteriene a propionibacteriilor ajunge la 109 ufc/g in mijlocul branzei dupa 3 saptamani.
Pediococcus - sunt asociati cu ingredientele si provin din plante. Sunt bacterii G(+), catalazo-negative sau slab pozitive, cresc in 6,5% NaCl, 45oC si produc amoniac din arginina. Pot fi uneori confundati cu micrococii datorita formei asemanatoare. Pediococii nu sunt utilizati in culturi lactice desi ei se pot inmulti in branza maturata si pot fermenta lactoza reziduala pentru o perioada lunga de timp. Doar doua specii: P. pentosaceus si P. acidilactici au fost gasite in produse lactate, dar nici una nu fermenteaza lactoza.
Pediococii au fost identificati prima data in Noua Zeelanda si mai tirziu in Anglia si s-a considerat ca imbunatatesc aroma. Ei produc D- si L-lactat din lactoza si racemizeaza L-lactatul. Efectul lor este neglijabil pana la 106-107 ufc/g, iar cresterea lor in branzeturi depinde de temperatura.
Pediococii apar in branza Cheddar si exista un interes crescut fata de activitatea acestora deoarece unele tulpini au activitate antimicrobiana fata de Listeria monocytogenes, Staphylococcus aureus si Clostridium perfrigens. Intr-un studiu a 49 tulpini de P. pentosaceus in toate tulpinile selectionate au fost gasite aminopeptidaze ale valinei si leucinei, catava lipaze si esteraze, α-glucozidaza, β-glucozidaza si N-acetil-β-glucozamidaze. Bhowmik si Marth, intr-un studiu mai amanuntit au gasit aminopeptidaze intracelulare, proteaze, dipeptidaze si dipeptilaminopeptidaze in 6 tulpini de P. pentosaceus si 2 de P. acidilactici. S-a observat de asemenea ca αS1- si β-cazeinele purificate din laptele degresat sunt hidrolizate de enzimele provenite din aceste microorganisme. Autorii nu au putut detecta insa activitate esterazica in toate tulpinile de P. acidilactici studiate.Utilizarea lactozei ca sursa de energie este redusa pentru aceste microorganisme si variaza de la tulpina la tulpina. Recent s-a demonstrat ca toate tulpinile de de P. pentosaceus si P. acidilactici poseda β-galactozidaza intracelulara, care este mai mare in celulele crescute in prezenta lactozei decat cele in glucoza, indicand natura inductiva a sintezei β-galactozidazei.
Sinteza enzimei a fost indusa de prezenta in mediu a galactozei si lactozei. Glucoza nu induce sinteza enzimei in tulpina de P. pentosaceus ATCC 25745. Desi aceste microorganisme sunt considerate homolactice, deoarece produc lactat, producerea etanolului si acetatului a fost observata atunci cand P. pentosaceus PC 399 a fost cultivata pe medii cu diferite hexoze si pentoze. Proportia de lactat si acetat a fost mai mare cu cat cantitatea de riboza a crescut in substrat.
Culturi starter
In concluzie principala functie a culturilor starter este aceea de a produce acid lactic in timpul procesului de fermentare. Culturile starter contribuie, de asemenea la maturarea branzeturilor, unde enzimele lor sunt implicate in proteoliza si in transformarea aminoacizilor in compusi de aroma (Wallace, 1997).
Deoarece definitia culturilor lactice starter este inca o problema controversata s-a propus ca denumirea de cultura starter sa se aplice izolatelor care produc suficient acid pentru a reduce pH-ul laptelui la mai putin de 5,3 in 6h la 30-37oC.
Culturile starter sunt fie adaugate in mod deliberat la inceputul prelucrarii sau pot contamina in mod natural laptele, cum este cazul mai multor sortimente artizanale obtinute din lapte crud. Ele cresc in timpul prelucrarii, atingand o concentratie de 108ufc/g, in cateva ore de la inceperea prelucrarii.
Se utilizeaza:
culturi starter mezofile - pentru branza Cheddar, Gouda, Edam, Brie si Camembert;
culturi starter termofile (50-55oC) - pentru branzeturile tratare termic, branzeturile cu pasta tare ca: Emmental, Gruyere, Parmesan si Grana.
Principalele culturi folosite la fabricarea unor sortimente de branzeturi romanesti sunt prezentate in tabelul 3. Bacteriile lactice cel mai des folosite fac parte din genurile: Lactococcus, Lactobacillus, Streptococcus, Leuconostoc si Enterococcus.
Atat culturile mezofile cat si cele termofile pot fi impartite in culturi mixte (nedefinite), culturi in care un numar de tulpini este necunoscut si culturi definite care sunt formate dintr-un numar cunoscut de tulpini.
Tulpinile nedefinite sau mixte de culturi mezofile sunt formate in special din Lactococcus lactis ssp. cremoris si Lactococcus lactis ssp. lactis, care pot contine si tulpini ce metabolizeaza citratul, pentru formarea aromei. Un studiu a 113 tulpini izolate dintr-o cultura mixta utilizata in mod curent a aratat ca o serie de tulpini au un profil diferit al plasmidului, capacitate diferita de a produce acid si rezistenta diferita la fagi, confirmand faptul ca reprezinta amestecuri (Lodics si Steenson, 1993). Bacteriile termofile pot fi formate din tulpini singulare sau multiple: Lactobacillus delbrueckii ssp. delbrueckii, Lactobacillus delbrueckii ssp. bulgaricus, Lactobacillus delbrueckii ssp.lactis sau Lactobacillus helveticus. Culturile nedefinite sau mixte de bacterii lactice termofile se utilizeaza pe scara redusa pentru obtinerea branzeturilor Gruyere, Emmental si Grana din Franta, Elvetia si Italia. Aceste culturi sunt produse de obicei prin incubarea zerului produs in ziua precedenta de productie la 40-45oC. Flora este formata din Streptococcus termophillus, enterococi si bacterii din genul Leuconostoc si mai pot exista specii de lactobacili: Lactobacillus helveticus, Lactobacillus delbrueckii, Lactobacillus acidophillus si Lactobacillus fermentum. O cultura starter cultivata pe zer, continand BL termofile este utilizata la fabricarea Parmesanului, sortiment de branza tare din nordul Italiei.
Tabelul 3. BL folosite pentru diferite tipuri de branzeturi
|
Culturi utilizate |
Tipuri de branzeturi |
|
Lactococcus lactis ssp. lactis Lactococcus lactis ssp. lactis var. diacetylactis |
Branza proaspata de vaci |
|
Lactococcus lactis ssp. lactis Lactobacillus casei |
Telemea, cas |
|
Lactococcus lactis ssp. lactis Lactococcus lactis ssp. cremoris Lactobacillus casei |
Branzeturi albe in saramura tip fetta, branzeturi frimantate: burduf, Moldova |
|
Lactococcus lactis ssp. lactis Streptococcus thermophillus Lactobacillus casei |
Branzeturi cu pasta oparita |
|
Lactococcus lactis ssp. lactis Streptococcus lactis ssp. cremoris Lactococcus lactis ssp. lactis var. diacetylactis |
Branzeturi semitari tip Trapist, Olanda, Edam |
|
Streptococcus salivarius ssp. thermophillus Lactobacillus delbrueckii ssp. bulgaricus Lactobacillus delbrueckii ssp. lactis Lactobacillus helveticus Propionibacterium |
Branzeturi tari: svaiter |
|
Lactococcus lactis ssp. lactis Lactococcus lactis ssp. lactis var. diacetylactis Lactococcus lactis ssp. cremoris Lactobacillus helveticus Lactobacillus casei |
Tip Cheddar |
Un studiu care a urmarit caracterizarea a 4379 izolate bacteriene din 35 de sortimente obtinute in mod traditional in Europa a aratat ca: 38% din izolate erau Lactococcus, 17% Entorococcus, 14% Streptococcus termophillus, 12% Lactobacillus mezofili, 10% Leuconostoc, 9% Lactobacillus termofili.
Din bacteriile genului Leuconostoc ssp si izolatele de Lactobacillus mezofili doar 8% si respectiv 2% produc suficient acid pentru a reduce pH-ul laptelui la valori mai mici de 5,3 in 6 ore si la 30oC.
Spre deosebire de acestea 53%, 32% si 13% din tulpinile de Streptococcus, Lactobacillus termophillus si izolatele de Enterococcus reduc pH-ul la valoarea 5,3. Acest lucru indica ca majoritatea izolatelor bacteriene mezofile din branzeturile fabricate prin procedee traditionale nu produc suficient acid lactic atunci cand sunt testate individual pentru fabricarea branzeturilor si deci nu pot fi denumite bacterii "starter".
Multe izolate termofile erau au fost gasite bune producatoare de acid lactic, deci potrivite pentru a fi denumite culturi starter. Enterococii prezenti ca parte naturala a microflorei unor produse traditionale au provocat multe dezbateri, deoarece exista dovezi ca pot fi asociati cu infectii clinice ale pacientilor cu sistemul imunologic compromis, sau pacienti aflati sub tratament (Boyce, 1992).
In ciuda acestor probleme, enterococii au fost gasiti in concentratii mari in branzeturile din zona mediteraneana si se considera ca au un rol pozitiv in dezvoltarea aromei.
Morfologia celulei, izomerii, rata de acid lactic produs si capacitatea de a metaboliza citratul, capacitatea de a creste la 10oC, 40 si 45oC, utilizarea glucozei, galactozei si lactozei sunt prezentate in tabelul 4.
Tabelul 4. Taxonomia si caracteristicile bacteriilor lactice
|
Microorganisme |
Denumire veche |
Forma |
Reducerea turnesolului in lapte inainte de coagulare |
% acid lactic produs in lapte |
Izomerul lactat produs |
Metabolizare citrat |
NH3 din arginina |
Crestere |
Fementeaza |
||||
|
10oC |
40oC |
45oC |
Glu |
Gal |
Lac |
||||||||
|
Streptococcus termophillus Lactobacillus helveticus Lactobacillus delbrueckii ssp. bulgaricus Lactococcus lactis ssp. cremoris Lactococcus lactis ssp. lactis Lactobacillus delbrueckii ssp. lactis Leuconostoc lactis Leuconostoc mesenteroides ssp. cremoris |
Neschimbata Neschimbata Lb. bulgaricus Str. cremoris Str. lactis Lb. lactis Neschimbata Leuconostoc cremoris |
streptococi Bastonase Bastonase Coci Coci Bastonase Coci diplococi |
<0,5 |
L DL D L L D D D |
| ||||||||
+/- - majoritatea tulpinilor pozitive;
-/+ - majoritatea tulpinilor negative.
Influenta CS asupra calitatii branzeturilor
Obtinerea branzei Cheddar omogena din punct de vedere calitativ necesita o fermentare uniforma a lactozei si proteoliza adecvata. La o rata mare, impreuna aceste procese depind de temperatura si de concentratia de sare, care trebuie sa fie distribuita cat mai uniform posibil. Rata de fermentare a lactozei de catre culturile starter din branza Cheddar depinde de raportul sare/umiditate in branza proaspata (Thomas, 1985). La niveluri scazute S/U, toata lactoza (4%) a fost utilizata, in 8 zile, in timp ce metabolismul a fost stopat la 6% S/U, astfel incat concentratia lactozei in branza a ramas foarte ridicata pentru mai multe saptamani dupa fabricare.
Lactoza reziduala a fost utilizata de BLNS, dar nu au existat corelatii puternice intre lactoza reziduala si cresterea BLNS, indicand faptul ca ele utilizeaza alte surse de energie in afara de lactoza si/sau alti factori de crestere. Conform lui Marley si Lawrance (1972), tulpinile de Lactococcus lactis ssp. lactis au supravietuit mai bine decat Lactococcus lactis ssp. cremoris, in prezenta sarii in concentratie de 4-5% NaCl.
Unul dintre rolurile de baza ale bacteriilor lactice starter este de a furniza un mediu adecvat, in acord cu potentialul redox, pH-ul si continutul de umiditate din branza, care sa permita o activitate enzimatica adecvata pentru enzimele din cheag si cultura starter si cresterea microflorei secundare (daca este necesar). Temperatura in timpul fabricarii si raportul S/U trebuie controlate pentru a se asigura ca o activitate corespunzatoare a culturilor starter este suficienta pentru a atingerea valorii de pH ceruta in timpul unei zile de prelucrare.
Cultura starter are o contributie semnificativa numai pentru biomasa microbiana a BL din branza tanara. Aceasta biomasa formata din cultura starter relativ mare reprezinta un potential biocatalitic considerabil pentru reactiile de maturare ale branzeturilor, care pot fi modelate prin autoliza celulelor bacteriilor starter. Activitatea autolitica a lactococilor se datoreaza unei muraminidaze (Mou, Sullivan, 1976, Niskasaari, 1989).
Autoliza celulelor starter poate fi influentata de concentratia de sare, de valorile sarii asociate cu umiditatea in branza (Bie si Sjostrom, 1975) si de variatiile conditiilor de fabricare, cum ar fi temperatura ridicata de prelucrare. Ar trebui ca microorganismele starter sa atinga concentratii mari si sa supravietuiasca prea mult pentru ca defecte de aroma precum aparitia gustului amar sa se produca.
Intr-un studiu realizat de Wilkinson, Guinee, O`Callaghan si Fox (1994), in care au fost monitorizate viabilitatea culturilor starter si eliberarea enzimelor starter intracelulare, s-a concluzionat ca tulpinile de Lactococcus lactis ssp. cremoris au avut modele diferite de autoliza.
Concentratii ridicate de aminoacizi liberi au fost produse de majoritatea tulpinilor autolitice (AM2) comparativ cu tulpinile mai putin autolitice (HP). Crow s.a. (1995a) au stabilit ca celulele starter intacte au fermentat lactoza, au consumat oxigenul si probabil au initiat un numar de reactii de formare a compusilor de aroma, in timp ce procesele peptidolitice au fost accelerate de celulele autolizate. In casul tanar, nivelul celulelor moarte este foarte important pentru maturarea branzei Cheddar (Crow, Martley, 1995). Produsii finali ai peptidolizei- aminoacizii- se acumuleaza mai repede daca se formeaza prin autoliza culturilor starter si sunt precursori majori ai compusilor de aroma (Urbach, 1995).
In branza Cheddar celulele de Lactobacillus helveticus autolizeaza foarte repede si formeaza niveluri mari de aminoacizi, care contribuie semnificativ la imbunatatirea aromei.
Microflora non-starter
Microflora non-starter sunt culturi mezofile de lactococi si pediococi, care reprezinta o parte importanta a florei microbiene in majoritatea branzeturilor in timpul maturarii. Ele nu fac parte din microflora normala starter si de regula nu cresc in bune conditii in lapte (Cogan, s.a, 1997) si nu contribuie la producerea acidului in vanele de coagulare.
Lactobacilii se impart in mod traditional in trei grupe:
(a)obligatoriu homofermentativi;
(b) facultativ heterofermentativi;
(c)obligatoriu heterofermentativi.
Lactobacilii care fac parte din BLNS (bacterii lactice non-starter) si sunt intalniti in mod obisnuit in branza fac parte din grupul II - facultativ heterofermentative si de aceea sunt denumiti LFH. Multe specii de lactobacili mezofile au fost izolate din branzeturi, dar cel mai frecvent se intalnesc: Lactobacillus casei, Lb. paracasei, Lb. plantarum, Lb. rhamnosus si Lb. curvatus (Jordan si Cogan, 1993).
Pediococcus acidilactici si Pediococcus pentosaceus sunt cei mai intalniti pediococi din branzeturi.
Influenta BLNS asupra calitatii branzeturilor
BLNS in branzeturile semitari maturate (Gouda si din Norvegia), fabricate din lapte pasteurizat sunt dominate de lactobacilii mezofili, predominanti fiind Lactobacillus casei, Lb. paracasei si Lb. rhamnosus. BLNS pentru branzeturile din Grecia sunt dominate de Lb. plantarum. In branzeturile Feta si Telemea 47,8% si 65,8% din lactobacilii izolati au fost reprezentati de Lb. plantarum (Tzanetakis, 1992).
Lb. plantarum a fost evidentiat in cantitati mari in branza de 2 zile si numarul microorganismelor a scazut in timpul maturarii, in timp ce Lb. paracasei prezent initial in cantitati mici a fost gasit dominant in microflora branzei la sfarsitul maturarii (Zarate, 1997).
Lb. paracasei domina microflora branzei Italiane. Acesta nu a fost depistat imediat dupa fabricare, dar niveluri ridicate (107 ufc/g) s-au gasit in primele luni de maturare si au ramas ridicate pe o durata de 120 zile.
Durata mare de maturare este o caracteristica a branzeturilor Parmigiano Reggiano. Evolutia microflorei bacteriene a fost studiata in 24 luni de la maturare.
Lb. paracasei, Lb. rhamnosus si Pediococcus acidilactici au fost gasite predominant ca BLNS in banzeturile italiene. In timpul maturarii, concentratia a scazut de la aproximativ 108 ufc/g (la 5 luni de la maturare) la aproximativ 104 ufc/g la 24 luni de maturare (Coppola, 1997).
Branzeturile Elvetiene (Emmental, Gruyere) pot fi fabricate din lapte crud, termizat sau pasteurizat, iar concentratia BLNS este de aproximativ 106 ufc/g. Populatia microbiana initiala este formata din Lb. paracasei, Lb. plantarum si Lb. brevis, in timp ce in branza maturata Lb. paracasei devine dominant in BLNS.
Branza Cheddar din Marea Britanie, dupa 6-9 luni de maturare a cuprins Lb. paracasei/ Lb. casei, Lb. plantarum -drept sepcii dominante, dar si Lb. curvatus, Lb. brevis, Lb. fermentum, Lb. bifermentas, Lb. buchneri, Lb. parabuchneri, Lb. farciminis si Lb. kefir (Williams si Banks, 1997).
Sursa de energie utilizata de BLNS pentru crestere este de obicei lactoza. BLNS pot transforma izomerul lactat L in izomerul D, dar nu se doreste utilizarea izomerului D ca sursa de energie. Citratul prezent in branza Cheddar tanara (aproximativ 8 mmol/kg) poate reprezenta o potentiala sursa de energie pentru BLNS (Jimeno, 1995). O serie de studii asupra branzei Cheddar au indicat o utilizare redusa a citratului in branza (Jordan, Cogan, 1993), deci concentratii mari de BLNS pot fi prezente si in absenta citratului.
Factorii care influenteaza cresterea microorganismelor in branza:
Umiditatea
Cresterea continutului de apa in masa de branza mareste susceptibilitatea la degradari. Activitatea apei este direct proportionala cu continutul de umiditate din branza si invers proportionala cu concentratia de sare si alti compusi cu masa moleculara mica (Esteban,1989). Indicele de activitate al apei aw este un concept termodinamic definit ca raport intre presiunea vaporilor de apa prezenti in sistem (p ) si presiunea celor din apa pura (p0) la aceeasi temperatura.
![]()
In timpul primelor stadii de fabricare a branzei aw este de aproximativ 0,99 ceea ce favorizeaza cresterea si activitatea culturilor starter.
Dupa scurgere, sarare si in timpul maturarii aw are valori cuprinse intre 0,917-0,988. Aceste valori sunt semnificativ mai scazute decat optimul necesar bacteriilor lactice. De aceea se poate considera ca aw contribuie la controlul metabolismului si multiplicarii bacteriilor lactice (Brow,1976).
BL (bacteriile lactice) in general au o valoare minima a aw mai mare decat a altor bacterii din branza. Indicele de activitate al apei minim pentru Lactoccocus lactis, Streptococcus therpmophillus, Lactobacillus helveticus si Popionibacterium freudenreichii ssp. shermanii este de 0.93;0,98; 0.96 si respectiv 0,96 (Weber, 1987).
O scadere a aw in timpul maturarii branzeturilor se datoreaza pierderilor de apa prin sarare, hidrolizei proteinelor la peptide si aminoacizi si trigileridelor la glicerol si acizi grasi, hidroliza fiecarei legaturi peptidice sau esterice necesitand o molecula de apa. La o proteoliza semnificativa in branzeturi, in timpul maturarii continutul de apa nelegata scade (Cogan 2000).
Tabelul 5. Valoarea minima a aw pentru dezvoltarea patogenilor
|
Patogeni |
aw |
|
Aeromonas hydrophila | |
|
Bacillus cereus | |
|
Campylobacter jejuni | |
|
Clostridium botulinum A, B,E, G | |
|
Escherichia coli | |
|
Listeria monocytogenes | |
|
Salmonella ssp. | |
|
Shigella ssp. | |
|
Staphilococcus aureus | |
|
Vibrio parahaemoliticus | |
|
Yersinia enterocolitica |
Controlul pierderilor de umiditate se efectueaza prin cresterea umiditatii relative in camera de maturare, sau prin ambalarea branzei in parafina sau material plastic. Pot exista variatii in valoarea aw pentru diferite zone ale branzei; pentru branzeturile tari si semitari in saramura valoarea aw creste de la periferie spre centru.
Branza Cheddar este obtinuta prin sarare uscata si ambalata in vid, de aceea nu apar pierderi de umiditate, daca sarea este uniform repatizata in branza.
aw influenteaza seminificativ si dezvoltarea bacteriilor patogene in branzeturi(tabelul 5).
Sarea
Sarea are un efect inhibitor pentru microorganismele din culturile starter si asupra bacteriilor de alterare, concomitent cu reducerea indicelui de activitate al apei. Concentratia ceruta depinde de natura alimentelor, pH si continutul de umiditate, dar in general 10-100 g/kg este o cantitate suficienta. Scaderea indicelui de activitate al apei apare atunci cand sarea este dizolvata in apa.
Relatia dintre concentratia de sare (g/kg branza) si aw este data de Cogan (2000):
![]()
Concentratia de sare variaza de la 0,7 la 7 g/100g, corespunzator valorilor pentru aw cuprinse intre 0,95-0,99 (tabelul 5). Multe microorganisme pot creste in aceste conditii. Efectul inhibitor este dat de raportul % S/U - sare dizolvata in umiditatea branzei, mai curand decat de sarea adaugata, sau sararea branzei.
NaCl inhiba activitatea enzimelor produse de microoganisme si afecteaza sinereza coagului, determinand eliminarea zerului, deci reducerea continutului de apa din branza, cu consecinte asupra activitatii microflorei si a enzimelor.
pH-ul si acizii organici
pH-ul optim pentru cresterea majoritatii bacteriilor este situat in jurul valorii neutre, iar cresterea este adeseori incetinita la pH mai mic de 5. Datorita acumularii acizilor organici, pH-ul branzei dupa prelucrare este cuprins intre 4,5-5,3. La astfel de valori scazute nu supravietuiesc speciile sensibile la aciditate ridicata. Inhibitorul real este de fapt forma nedisociata a acizilor organici (Beuchat si Golden, 1989).
Principalii acizi organici gasiti in branza sunt: acidul lactic, acidul acetic si propionic care au un pKas de 3,08; 4,75 si respectiv 4,87, asadar acidul lactic are cel mai slab iar acidul propionic cel mai puternic efect inhibitor, la aceeasi concentratie si pH al branzei. Oricum, lactatul in branza este prezent la o concentratie mult mai mare decat ceilalti doi acizi, cu exceptia branzei Elvetiene maturate, unde concentratia acidului propionic poate fi mai mare decat a acidului lactic (Steffen, Rueg, 1993).
Temperatura de maturare
Microorganismele implicate in fabricarea si maturarea branzeturilor sunt mezofile si termofile, avand temperatura optima de dezvoltare de aproximativ 30o sau 42oC. Temperatura la care se realizeaza maturarea branzeturilor este un compromis intre necesitatea de a promova reactiile de maturare si controlul cresterii microflorei secundare utile cat si nevoia de a preveni propagarea potentialelor bacterii de alterare si patogene. Temperaturi mari de maturare promoveaza accelerarea maturarii (McSweeny, 1996), dar modifica structura si aroma produsului, deseori in sensul nedorit.
Potentialul redox
Potentialul redox (Eh) este o masura a capacitatii unui sistem chimic/biochimic de a se oxida (de a pierde electroni) sau reduce (accepta electroni). O stare de oxidare sau reducere este indicata de valorile pozitive sau negative ale potentialului, exprimate in mV.
Eh-ul laptelui este de aproximativ 150 mV, in timp ce in branzeturi este de aproximativ -250 mV. In timp ce mecanismul reducerii Eh in branza nu a fost complet elucidat, se presupune totusi ca este legat de fermentarea acidului lactic de catre culturile starter in timpul cresterii si reducerea unor cantitati mici de oxigen din branza, la apa (Crow, 1995). Ca o consecinta a acestor reactii, in interiorul branzei exista un sistem in buna masura anaerob, care poate sprijini cresterea microorganismelor facultativ anaerobe sau anaerobe. In diferite culturi microbiene, Eh poate varia de la +300 mV pentru microorganismele aerobe la mai putin de -400 mV pentru microorganismele anaerobe (Brown, 1980).
Eh-ul branzei este unul din factorii majori pentru determinarea tipului de microorganism care poate creste in branza. De aceea, microorganisme aerobe ca Pseudomonas, Brevibacterium, Bacillus si Micrococcus nu cresc in interiorul branzeturilor. Aceste bacterii se pot dezvolta doar la suprafata branzeturilor.
Nitratii
Nitratii (NO3-) se adauga in laptele destinat fabricarii branzeturilor sub forma de KNO3 sau NaNO3, in special in sortimentele Olanda, Edam si Gouda pentru a preveni cresterea lui Clostridium tyrobutyricum, care fermenteaza lactatul la butirat, H2 si CO2:
2 lactat → 1 butirat + 2 CO2 + 2 H2
H2 si CO2 sunt responsabili pentru gaurile mari si lipsa aromei. La aceste sortimente de branzeturi, rata migrarii NaCl este relativ lenta si egalizarea concentratiei de sare in branzeturi poate lua cateva saptamani. In plus, continutul de umiditatea este mare. De aceea, inhibarea cresterii microorganismelor de alterare, precum Clostridium tyrobutyricum este necesara, in principal pentru a atinge echilibrul salin, iar nitratii joaca acest rol.
In timpul maturarii, nitratul se reduce la nitrit (inhibitorul real al cresterii) sub actiunea enzimei xantinoxidaza, de origine indigena (Niuwenhoff, 1977). Nitritii nu afecteaza cresterea BL, dar inhiba proprionibacteriile responsabile de formarea ochiurilor de fermentare in branza Emmental, si de aceea nu se utilizeaza pentru controlul lui Clostridium tyrobutyricum, acolo unde este dorita prezenta golurilor de fermentare. Nitritii pot reactiona cu aminoacizi aromatici din branzeturi producand nitrozamine. Majoritatea nitratului adaugat la fabricarea branzeturilor se regaseste in zer sau difuzeaza in saramura. Odata ce branzeturile sunt gata de consum, nivelul nitritului este mult sub nivelul admis (50 mg/kg), conform prevederilor pentru branza Olanda.
Cresterea culturilor starter de bacterii in lapte
Laptele este un mediu de cultura foarte bun pentru cresterea bacteriilor lactice. De fapt, Lactobacillus delbrueckii ssp. bulgaricus, Streptococcus salivarius ssp. thermophillus si Lb. helveticus au laptele ca mediu preferat de crestere si utilizeaza lactoza pentru crestere.
Producerea de acid lactic de catre culturile starter depinde chiar de calitatea laptelui. Cresterea poate fi promovata sau inhibata de anumiti factori care actioneaza in lapte. Abilitatea culturilor starter de a se multiplica in lapte depinde partial de activitatea lor proteolitica.
Lactococcus lactis ssp. lactis creste intr-un mediu cu cazeinat, ca unica sursa se azot, in timp ce Lactococcus lactis ssp. cremoris necesita o sursa suplimentara de aminoacizi. Toate bacteriile lactice utilizate in industria laptelui necesita sau sunt stimulate de prezenta aminoacizilor. Aminoacizi liberi existenti in lapte nu sunt corespunzatori, iar bacteriile lactice utilizeaza proteinazele, peptidazele si sistemele de transport pentru a-si indeplini cerintele lor nutritionale. Concentratii minime de aminoacizi sunt necesare pentru unele bacterii lactice pentru maximizarea cresterii. Aminoacizi ca: acidul glutamic, leucina, izoleucina, valina, arginina, cisteina, prolina, fenilalanina si metionina sunt considerati foarte importanti pentru nutritia lactococilor.
Nu este neobisnuit ca in timpul transferului si a propagarii BL microorganismele sa-si diminueze activitatea si sa fermenteze lactoza mai incet. Aceasta se poate datora pierderii uneia sau mai multor plasmide care controleaza metabolismul lactozei si al proteinelor.
Inhibitori ai bacteriilor starter
Bacteriocinele- Calitatea bacteriologica a laptelui si durata depozitarii inainte de utilizare sunt doua aspecte deosebit de importante. Laptele contine intotdeauna microorganisme care pot creste si utiliza aminoacizii si peptidele din lapte pentru a produce inhibitori (bacteriocine), activitatea inhibitoare fiind considerabila si la concentratii foarte mici. Maltick si Hirsch au izolat un inhibitor (nizina) din Lactococcus lactis ssp. lactis, care este foarte eficient impotriva bacteriilor G(+), inclusiv a starterilor, a lactobacililor si a microorganismelor sporulate. Din Lactococcus lactis ssp cremoris a fost izolata diplocina, cu un spectru limitat de activitate.
Lipoliza - In laptele depozitat se pot multiplica microorganisme psihotrofe si daca se gasesc intr-o proportie mai mare de 106-107 ufc/ml pot produce lipoliza. Acizii grasi cu C4 C12 in lant si acidul sorbic din branzeturi au efect inhibitor pentru cultura starter. Microorganismele starter acumuleaza acizii grasi liberi la suprafata celulei si nu ii metabolizeaza. Celulele ramase libere din grupul N al lactococilor la pH=4,5 metabolizeaza piruvatul cu formare de acetat (acizi volatili), acetoina si diacetil, CO2. In prezenta acidului oleic, utilizarea piruvatului a fost maxima la pH=6,5 si complet inhibata la pH= 4,5.
Apa oxigenata - metabolizata de grupa N a lactococilor prin actiunea nicotinamidadenin dinucleotid-oxidaza, care catalizeaza oxidarea NADH in prezenta O2 molecular. Enzima este activata de FAD. O parte din apa oxigenata formata este indepartata de NADH peroxidaza:
NADH + H+ + O2 ![]() NAD+ + H2O2
NAD+ + H2O2
NADH + H+ + H2O2 ![]() NAD+ + 2H2O
NAD+ + 2H2O
Laptele este agitat in timpul umplerii vanei si adaugarea culturii starter odata cu adaugarea cheagului la fabricarea branzeturilo conduce la formarea unei cantitati mici de H2O2 in lapte.
Adaugarea unor cantitati mici de H2O2 are un efect negativ asupra dezvoltarii lactococilor.
In lapte, culturile de lactococi si lactobacili produc la randul lor H2O2 in perioadele initiale de producere a acidului lactic, urmate de o reducere drastica a acumularii de H2O2 odata cu cresterea cantitatii de acid produsa.
Adaugarea sulfatului feros si a catalazei previne sau reduce acumularea H2O2 si stimuleaza producerea de acid. Adaugarea micrococilor reduce nivelul H2O2 in mediu si stimuleaza producerea de acid.
Sistemul lactoperoxidaza/Tiocianat/ H2O2
H2O2 produsa poate inhiba o serie de tulpini de lactococi si indirect culturile introduse in lapte, prin oxidarea tiocianatului prezent in lapte intr-un produs inhibitor, reactie catalizata de lactoperoxidaza. Concentratii mici de H2O2 formeaza un complex cu lactoperoxidaza (LP) care stabilizeaza puterea oxidativa a H2O2, catalizand oxidarea tiociantului SCN-, conform reactiei:
H2O2 + SCN- ![]() OSCN- + H2O
OSCN- + H2O
OSCN- + H2O2 ![]() O2SCN- + H2O
O2SCN- + H2O
O2SCN- + H2O2 ![]() O3SCN- + H2O
O3SCN- + H2O
Produsele finale ale oxidarii tiocianatului sunt CO2, NH4+, SO42-, care sunt inerte, dar reprezinta produse de oxidare intermediare. OSCN- este inhibitor pentru microorganismele G(+), microorganismele starter, coliformi, salmonele si alte microorganisme G(-). In conditii aerobe OSCN- afecteaza membrana interna a altor pereti celulari ai microorganismelor.
Wright si Tramer au aratat ca o serie de culturi starter sunt inhibate de prezenta peroxidazei din lapte, care poate fi prevenita prin adaugarea cisteinei sau generarea unor grupe -SH prin incalzire.
Tiocianatul a fost indepartat din lapte cu o coloana de schimb ionic si s-a aratat ca tulpina Streptococcus cremoris 972, peroxidazo-senzitiva nu a fost inhibata de producerea de acid lactic la fabricarea branzeturilor. Adaugarea tiocianatului previne substantial dezvoltarea acidului, similar comportamentului culturilor starter infectate de fagi.
Standhouders si Veringa au aratat ca exista o corelatie intre inhibarea lactococilor si prevenirea inhibarii streptococilor lactici de cisteina. Ei au explicat ca intr-un amestec de H2O2, cisteina si peroxidaza din lapte, cisteina este oxidata si se comporta ca donor de H+. Daca cisteina si peroxidaza din lapte sunt incubate in absenta H2O2, cisteina si enzima formeaza un compus ireversibil. Daca se adauga apa oxigenata atunci cisteina se comporta ca un inhibitor al enzimei. Ei au presupus ca varianta sensibila la peroxidaza a streptococilor lactici are probabil nevoie neaparat de cisteina libera, dar cisteina a fost complexata cu peroxidaza. Peroxidaza din lapte este inhibata de prezenta cantitatilor foarte mici de hidrogen sulfurat care este produs in timpul incalzirii laptelui.
Susceptibilitatea culturilor lactice starter la lactoperoxidaza/H2O2/tiocianat (LPS) este dependenta de:
sensibilitatea tulpinii;
abilitatea tulpinii de a produce H2O2, care activeaza sistemul LPS;
prezenta enzimelor nespecifice, de exemplu: xantinoxidaza sau hipoxantin oxidaza, care produc H2O2.
Sistemul inhibitor este labil la caldura si este distrus pinr tratamentul termic al laptelui care contine culturi starter.
Tratamentul termic
Laptele este tratat termic pentru a putea fi conservat si pentru a fi sigur in consum. Intensitatea tratamentului depinde de produsul finit si utilizarea intentionata. Multi specialisti au studiat efectul caldurii asupra activitatii culturii starter. Este unamim recunoscut ca utilizarea unor culturi diferite determina o activitate diferita atunci cand aceste culturi au fost inmultite in lapte si a fost necesara aplicarea unor tratamente termice diferentiate in functie de tipul de culturi.
Olson, Gilliland si Speck au observat ca producerea de lactococi a fost mai mare in laptele pasteurizat la 71,1o/30 minute urmata de laptele sterilizat la 121,1oC/15 minutesi la: 61,6oC/82,2oC si 98,8oC pentru 30 minute. Acele culturi care produc acid repede in laptele pasteurizat la 61,6oC/30 minute sau la 71,7oC/15 secunde au fost denumite "culturi pentru temperaturi joase". Culturile care produc acid repede in laptele tratat la temperatura inalta au fost denumite "culturi pentru temperaturi inalte".
Au fost testate 37 de culturi comerciale, din care 49% au fost clasificate ca fiind "culturi pentru temperaturi joase", 35% ca fiind "culturi pentru temperaturi inalte" si 16% au prezentat un comportament nespecific.
Pentru culturi termofile: Lactobacillus delbrueckii ssp. bulgaricus si Streptococcus salivarius ssp. thermophillus, a fost studiat efectul tratamentului termic a laptelui in regim HTST (80oC/10 minute). Nu s-a observat un efect semnificativ asupra cresterii lui Streptococcus salivarius ssp. thermophillus dar a fost stimulat in crestere Lactobacillus delbrueckii ssp. bulgaricus, efectul crescand cu severitatea tratamentului termic. La 95oC/10 minute a aparut un efect de stimulare doar la culturile in amestec. Factorul stimulativ a putut fi inlocuit de acidul formic. Producerea de acid formic de catre Streptococcus salivarius ssp. thermophillus a fost confirmata experimental.
Aglutinarea
Proprietatile inhibitoare ale anticorpilor cu insusiri de aglutinare este minor pentru culturile starter din tancuri, iar pe masura ce tratamentul termic este aplicat sau formarea coagulului are loc, in timpul fabricarii branzeturilor se distruge aceasta capacitate de inhibare. Oricum, aglutininele sunt importante si impactul negativ din branza Cotttage a fost evidentiat prin formarea unui depozit la partea inferioara a vanei, care a dus in timp la inactivarea culturii.
Antibioticele
Un nivel scazut al antibioticelor in lapte poate incetini activitatea culturilor starter si poate face mai dificila obtinerea branzei.
Heap a demonstrat ca in timp, lactococii pot creste si produce acid in laptele degresat reconstituit, continand diferite cantitati de penicilina. Producerea de acid a parut normala, dar activitatea culturii a fost mult mai scazuta (tabelul 6).
Tabelul 6. Niveluri critice ale penicilinei in laptele pentru diferite bacterii
|
Bacterii |
UI/ml |
|
Streptococcus cremoris Streptococcus lactis Streptococi (starter) Streptococcus thermophillus Streptococcus faecalis Lactobacillus bulgaricus Lactobacillus acidophillus Lactobacillus casei Lactobacillus lactis Lactobacillus helveticus Lactobacillus citrovorum Proprionibacterium shermanii |
Sisteme culturale
Culturile starter necesita conditii bune de pastrare in tancuri, circulatia corespunzatoare a aerului si un flux de productie adecvat, personal instruit in activitatea de insamantare, cu sau fara filtrarea zerului.
Amestecurile de culturi mezofile ce contin microflora nedefinita (Leuconostoc, Lactococcus lactis ssp. diacetylactis) sunt utilizate inca in Europa si SUA. Aceste culturi sunt utilizate fara a fi indeajuns cunoscute, sub forma concentrata si inghetata, si pot cauza probleme.
Culturi cu bacteriofagi, utilizate in Olanda (bacteriofagi care transporta culturi starter), denumite si culturi P. Aceste culturi sunt in echilibru cu fagii in mediul lor, iar fagii continuti nu influenteaza activitatea culturii. Cand un fag ataca tulpina dominanta poate apare o usoara diminuare a activitatii culturii, dar activitatea culturii este restabilita datorita prezentei unui numar mare de microorganisme cu fagi.
Institutul de Cercetari Olandez comercializeaza sub forma de starter P culturi concentrate si inghetate. Sistemul pare foarte bun, dar atunci cand starterii P sunt propagati in laborator fara contaminare cu fagi, ei devin sensibili la fagi. Acest comportament este atribuit unui numar mic de tulpini denumite starteri L.
Culturile DVS sunt utilizate din 1970. Ele sunt suspensii de celule foarte concentrate (1011ufc/g) a unor tulpini definite impreuna cu agenti crioprotectori ca: glicerolulul si lactoza, racite rapid in azot lichid si mentinute la temperaturi de
-196oC. Pentru transportul la fabrica se utilizeaza containere ambalate in gheata uscata.
Culturile DVS sunt alcatuite din 3 sau 4 tulpini definite, propagate impreuna sau separat.
Avantajele pe care le ofera culturile DVS:
convenabile - elimina necesitatea existentei unor tancuri, laboratoare si sisteme de aer steril. Pot contribui la suplimentarea sistemului conventional, in caz ca acesta nu mai este utilizabil;
culturi de incredere - ofera posibilitatea de standardizare a procedeului de fabricarea a branzeturilor;
imbunatattirea performantei zilnice - in fiecare zi au aceeasi activitate;
imbunatatirea calitatii branzeturilor
Dezavantaje:
necesita un refrigerator performant, costurile sunt compensate de munca de preparare a culturilor;
datorita cantitatii mici de acid dezvoltate sunt necesare cantitati mai mari de coagulanti. Pentru a creste taria casului, vana trebuie adusa la 32-33oC in loc de 30oC.
Motive pentru utilizarea restransa a culturilor DVS:
lipsa unui numar suficient de tulpini cu rezistenta la atacul fagilor;
multe tulpini sunt greu de controlat prin centrifugare;
activitatea culturilor congelate inoculate in vana este redusa in timpul taierii si incalzirii branzeturilor.
Etapele de fabricare trebuie adaptate la nivelul redus de acid produs in faza umeda si de nivelul ridicat de producere a acidului in faza uscata (cedarizare);
cultura DVS este mai scumpa;
In SUA au fost introduse sisteme automate de masurare a pH-ului culturilor starter si prin multiplicarea culturilor starter pe zer se pot aduce beneficii economice importane producatorilor, reducandu-se semnificativ pierderile.
Producerea si propagarea culturilor
In mod obisnuit, la obtinerea culturii de productie se trece de la cultura comerciala la cultura mama si la culturi intermediare, pentru a obtine in final cultura de productie ce va fi inoculata in tanc.
T Culturi starter sub forma lichida.
Primele procedee de obtinere a culturilor starter utilizau pentru insamantari un lapte praf cu 10-12% grasime, tratat termic la 90oC/45 minute, pentru inactivarea bacteriofagilor. Culturile erau livrate in butelii de sticla si utlizate in maxim 72 ore. Ele erau reinsamantate pentru a fi utilizate in fabrica.
T Pentru transport pe distante lungi se utilizau culturi sub forma de pulbere amestecate cu lactoza, neutralizate cu CaCO3 si ambalate sub vid. Astfel de culturi necesita numeroase reactivari (1-2% supravietuitori in pulbere).
T Culturile congelate si uscate contin 42-80% microorganisme viabile, in functie de tipul culturii. Aceste culturi cresc lent, avand o perioada lunga de lag. Pentru a reduce faza de lag se adauga stimulatori inainte de liofilizare, sau in substartul in care este reactivata cultura starter.
T Culturi concentrate au fost comercializate pentru prima data in 1973, eliminandu-se astfel etapa care necesita prepararea culturii mama si a culturilor intermediare. Aceasta metoda a minimizat riscul contaminarii cu bacteriofagi si a deschis drumul culturilor DVS. Laptele este pasteurizat la 90oC/45 minute, dupa care se face o racire la temperatura de inoculare (21-27oC), insamantare si termostatare la 27o pana cand pH-ul atinge valoarea 4,6. In acest moment o cultura cu 5-8x108ufc/ml prezinta o buna activitate. Daca celulele sunt mentinute la pH=5 pentru o perioada de timp indelungata, activitatea culturii se reduce. Populatia finala de lactococi este marita prin controlul pH-ului la 6-6,5. Cand cultura a fost propagata in mediu cu 2% triptona, 1% extract de drojdie 2,5% lactoza si 2,5% glucoza la pH constant 6 (mentinut prin adaugare de NaOH), populatia a crescut de 15 ori. La acest pH, rata de crestere si cantitatea totala au fost optime. Cand specii amestecate de tulpini bacteriene, continand specii formatoare de aroma au fost cultivate pe lapte degresat (9,1% SU), mediu de zer cu triptona la pH constant, producerea de acid lactic (%)/compusi de aroma (diacetil) a fost mai mare la pH 5,9.
Nu se recomanda cultivarea la pH sub 5,9-6,1. Cresterea cea mai mare (1010 ufc/ml)
s-a obtinut la 30-32oC/pH 6,0-6,3.
Adaugarea NH4OH in mediu a facilitat cresterea numarului de celule, mai mult decat NaOH.
Propagarea
Pentru prepararea culturii starter sunt necesare 106-107 ufc/ml.
Metode de pregatire a mediului de crestere pentru culturile starter si protejarea culturii de atacul bacteriofagilor:
(a) procedeul aseptic;
(b) utilizarea unor vase speciale pentru prevenirea contaminarii;
(c) utilizarea mediilor inhibitoare pentru fagi;
(d) utilizarea culturilor DVS liofilizate.
(a) Procedeul aseptic presupune existenta unei camere separate pentru inceperea insamantarii, clorinarea, sterilizarea si afumarea cu clor a incaperii inainte de inoculare. Acest procedeu este util, dar nu intotdeauna eficient pentru prevenirea contaminarii.
(b) Utilizarea tancurilor speciale
Sistemul Lewis - sticle speciale pentru cultura mama si culturile de alimentare, inchise cu sistem de sigilare Astell. Acest mediu este sterilizat si racit in sticla si cultura este transferata prin 2 ace hipodermice. Sistemul Lewis necesita vase sub presiune. Aerul nu circula in vase in timpul incalzirii/racirii.
Sistemul Jones - tancul nu este sub presiune. Deschiderile sunt protejate de sigilii cu apa. Aerul este fortat sa iasa in timpul incalzirii mediului si aerul steril (incalzit si filtrat) reintra in tanc in timpul racirii.
Sistemul AlfaLaval - sistemul in care cultura mama si culturile intermediare sunt propagate intr-un incubator si cultura este transferata intr-un tanc mare utilizand aerul sterilizat sub presiune.
Acest sistem utilizeaza filtre de aer particulate cu eficienta ridicata (HEPA). Producatorii olandezi utilizeaza starteri P preparati de Niro. Masuri: laptele este tratat la temperaturi mai mari de 95oC/1 minut si in timpul racirii, inocularii si cultivarii tancurile sunt presurizate cu aer steril. Filtre absolute tip Pall Enflonfilter ABI FR7PV permit trecerea a mai putin de 2,5x1010 fagi, asigurand ca tancul presurizat nu este contaminat cu fagi.
Tabelul 7. Cuturi starter utilizate pentru diferite sortimente de branzeturi:
|
Cheddar, Colby |
Lactococcus lactis ssp. lactis, Lactococcus lactis ssp. cremoris, Leuconostoc mesenteroides ssp. cremoris*, Lactococcus lactis ssp. lactis bvar. diacetylactis |
|
Elvetiana |
Streptococcus salivarius ssp. thermophillus, Lactobacillus helveticus sau Lactobacillus delbrueckii ssp. bulgaricus sau Lactobacillus delbrueckii ssp. lactis si Propionibacterium |
|
Parmesan, Romano |
Streptococcus salivarius ssp. thermophillus, Lactobacillus delbrueckii ssp. bulgaricus sau Lactobacillus helveticus sau Lactobacillus delbrueckii ssp. lactis |
|
Mozzarella, Provolone |
Streptococcus salivarius ssp. thermophillus, Lactobacillus delbrueckii ssp. bulgaricus sau Lactobacillus helveticus |
|
Blue, Roquefort si Stilton |
Streptococcus salivarius ssp. thermophillus, Lactococcus lactis ssp. lactis/cremoris, Lactococcus lactis ssp. lactis bvar. diacetylactis, Penicillium roqueforti |
|
Gorgonzola |
Streptococcus salivarius ssp. thermophillus, Lactobacillus delbrueckii ssp. bulgaricus, Penicillium roqueforti, Lactococcus lactis ssp. lactis bvar. diacetylactis sau drojdii |
|
Camembert |
Cultura Lactococcus/ Penicillium roqueforti |
|
Brick, Limburger |
Amestec de cultura de lactococi si Streptococcus salivarius ssp. thermophillus, Brevibacterium linens si drojdii |
|
Muenster |
Lactococcus lactis ssp. lactis |
|
Gouda si Edam |
Lactococcus lactis ssp. cremoris cu B sau BD-culturi de aroma |
|
Branza crema |
Lactococcus lactis ssp. lactis, Lactococcus lactis ssp. cremoris cu B sau BD-culturi de aroma |
|
Cottage |
Lactococcus lactis ssp. lactis, Lactococcus lactis ssp. cremoris |
B - Leuconostoc mesenteroides ssp. cremoris/Leuconostoc lactis; D - Lactococcus lactis ssp. lactis bvar. diacetylactis; BD - cultura mixta: Leuconostoc mesenteroides ssp. cremoris/Leuconostoc lactis si Lactococcus lactis ssp. lactis bvar. diacetylactis.
Recent, au fost realizate filtre groase, cu dimensiunea porilor de 0,015 μm, care pot retine bacteriofagii din aer. Aceste filtre sunt sisteme in-line de filtrare, care pot fi utilizate la 50 de cicluri.
Bactoscan utilizeaza o camera ultracurata cu 12 schimburi de filtre/ora. In acest sistem, vasele sub presiune sunt grupate astfel incat zonele de intrare sunt accesibile operatorului.
In tabelul 7 sunt prezentate diferiteculturi starter utilizate pentru obtinerea unor sortimente de branzeturi.
L-lactatdehidrogenaza
Bacteriile lactice
homofermentative produc fermentarea anaeroba a zaharurilor pe calea
Embden-Mayerhof-Parnas (EMP) pana la formarea acidului piruvic, apoi fiind
lipsite de enzima piruvat decarboxilaza, acidul piruvic devine acceptor de
hidrogen.
D-lactatdehidrogenaza
![]()
![]() L(+) lactat + NAD+ Piruvat
+ NADH + + H+
L(+) lactat + NAD+ Piruvat
+ NADH + + H+
![]()
![]() D(-) lactat + NAD+ Piruvat
+ NADH + + H+
D(-) lactat + NAD+ Piruvat
+ NADH + + H+
In fermentatia homolactica, forma predominanta a acidului lactic este dependenta de stereospecificitatea lactatdehidrogenazei (lactococii formeaza L(+)-lactat si lactobacilii D(-) lactat), dar si de prezenta lactat racemazei care atunci cand este activa in celula microbiana, rezulta prin fermentatie, amestecul racemic (D, L-lactat), catalizand reactia:
![]()
![]() L(+) lactat D(-)
lactat
L(+) lactat D(-)
lactat
Ecuatia globala a fermentatiei homolactice:
![]() C6H12O6
+ 2ADP + 2Pi 2 CH3-CHOH-COOH + 2ATP + 75 KJ
C6H12O6
+ 2ADP + 2Pi 2 CH3-CHOH-COOH + 2ATP + 75 KJ
Cai de formare a produsilor principali si secundari in fermentatia lactica

![]()
![]()
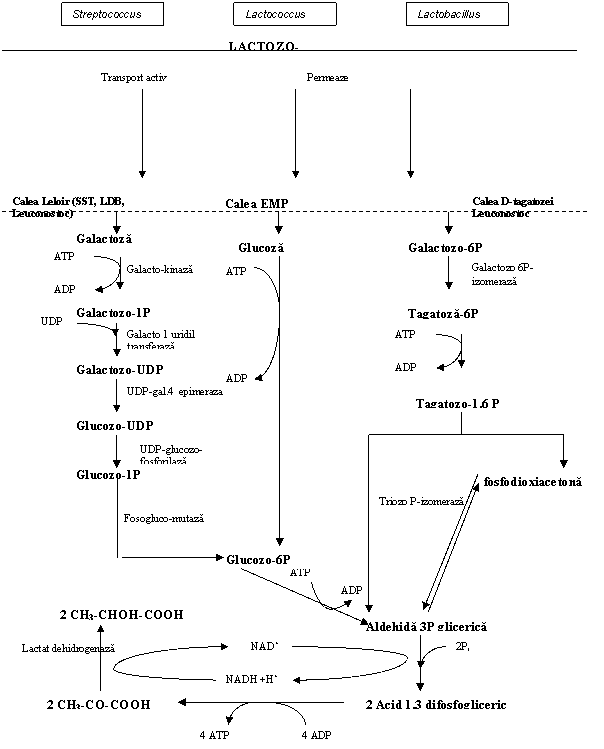
Fig. 2 Fermentatia lactica produsa de bacteriile homofermentative
Pentru a fi fermentate, glucoza, galactoza si lactoza pot fi transportate ca atare in celula printr-un sistem activ de permeaze prezent la Streptococcus salivarius ssp. thermophillus si lactobacili termofili, sau prin transport activ catalizat de sistemul fosfotransferaza-fosfoenolpiruvat (PT-PEP) prezent la bateriile din genul Lactococcus si Lactobacilus, cu formarea esterilor fosforici.
Lactoza sau lactozo-P in celula bacteriana sub actiunea β-galactozidazei (lactazei) este transformata in glucidele componente (Fig. 2).
In fermentatia homolactica se pot forma in cantitati mici substante de aroma, diacetil si acetoina, fie prin metabolizarea citratilor care asigura o sursa suplimentara de acid piruvic (fig.3).
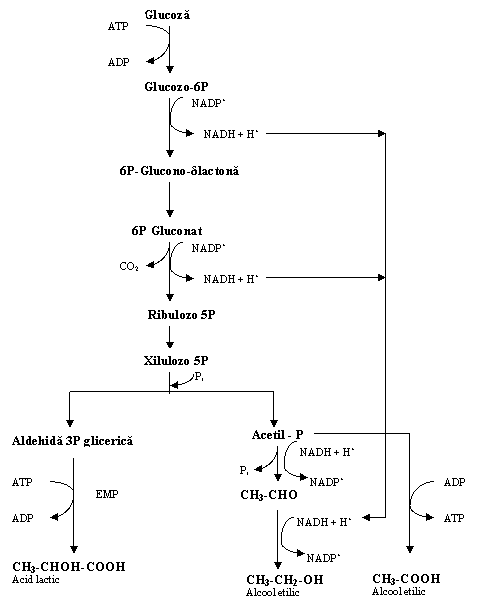
Fig. 3 Biochimismul fermentatiei heterolactice
Bacteriile lactice heterofermentative produc fermentatia anaeroba a glucidelor pe calea pentozo-fosfatului (6P-gluconatului) (Fig.3).
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 5964
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2026 . All rights reserved