| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Biosinteza proteinelor timpurii
Sinteza proteinelor cu specificitate virala este evenimentul esential al ciclului de multiplicare virala si necesita existenta unui mesaj viral generat in etapa transcrierii. Sinteza proteinelor virale semnifica traducerea unui mesaj strain celulei si este supusa unor restrictii.
O restrictie majora a sintezei proteinelor virale deriva din faptul ca in celula infectata, exprimarea genelor virale este in competitie cu activitatea numeroaselor gene celulare functionale. O singura copie a genomului viral, initiaza programul de dominanta asupra programului celular, de 104 - 106 ori mai mare, care este deja exprimat. Mesagerii virali sunt in competitie, pentru traducere, cu mesagerii celulari.
Proteinele virale sunt sintetizate de aparatul celular de sinteza proteica (aminoacizi, ARNt, ribosomi, enzimele catalizatoare).
Aparatul de sinteza proteica al celulei traduce numai mesaje unitare, si nu recunoaste situsurile multiple de intiere a traducerii unui ARNm viral policistronic. La unele virusuri, ARNm este policistronic, copie a catorva gene si este tradus intr-o poliproteina giganta, clivata ulterior in proteine individuale de marime adecvata. Poliproteina nu se evidentiaza la electroforeza in gel de poliacril-amida, deoarece este clivata chiar in timp ce se sintetizeaza. Inhibitia clivajului, prin incorporarea analogilor aminoacizilor, sau prin cresterea temperaturii face posibila detectarea ei la electroforeza.
Proteinele virale se sintetizeaza pe polisomii citoplasmatici. Pentru virusurile care se multiplica in nucleu, proteinele sintetizate in citoplasma trebuie sa fie transportate in nucleu, pentru a-si indeplini functia specifica: enzime de replicare a genomului sau proteine virale structurale.
Proteinele virale precoce apartin urmatoarelor categorii:
l) Proteine de reglare au rol de instituire si mentinere a ordinii virale. Ele inhiba sinteza macromoleculelor specifice celulei, prin modificarea specificitatii sistemelor enzimatice de replicare, transcriere si traducere, astfel incat sintezele macromoleculare ale celulei sunt stopate si metabolismul este orientat in sensul sintezei constituientilor virali. Proteinele virale cu rol in controlul expresiei genelor sunt multifunctionale ;
2) Proteine matriceale au rolul de a delimita viitoarea "fabrica de virus", adica teritoriul celular in care virusul se multiplica;
3) Proteine-enzime de tipul polimerazelor: ADN- si ARN-polimeraza, nucleaze (enzime de clivare), ligaze, proteaze. Virusurile codifica serin-proteaze, cistein-proteaze si aspartic-proteaze, dar nu codifica metaloproteaze. Serin-proteazele, la situsul activ contin o Ser reactiva, care impreuna cu Asp si His formeaza triada catalitica, ce cliveaza legaturile peptidice. Cistein-proteazele au o diada catalitica formata din resturi de Cys si His. Ambele categorii formeaza intermediari enzima-substrat. Aspartic-proteazele virale sunt asemanatoare cu omologele lor celulare(pepsina, gastrina, catepsina D, renina), in general sunt active la pH acid si nu par sa formeze intermediari enzima-substrat.
Cele mai multe virusuri, in faza timpurie exprima o cantitate foarte limitata de informatie genetica. Numai poxvirusurile exprima 30-50 de functii in faza precoce.
Replicarea genomului viral
In etapa timpurie se sintetizeaza proteine virale cu rol enzimatic, care conditioneaza procesul replicarii virale si implicit, evolutia complexului virus-celula gazda.
Pentru replicarea ADN viral sunt utilizati precursorii din mediul de crestere a celulelor, deoarece ADN celular nu este degradat. Celula sintetizeaza nucleotide, din care se sintetizeaza acizii nucleici virali. Procesul replicarii genomului viral a fost studiat prin tehnica autoradiografiei, ce consta in adaugarea in mediul nutritiv, a precursorilor nucleotidelor, marcati radioactiv si detectarea lor ulterioara in ADN viral. Replicarea genomului viral ADN este dependenta de actiunea catalitica a unei ADN-polimeraze.
Virusurile mici utilizeaza ADN-polimeraza celulara, iar cele complexe (adeno, herpes), codifica sinteza ADN-polimerazelor proprii. Virusurile herpetice codifica 7 proteine-enzime, cu rol in replicarea genomului.
Replicarea ADN viral se face dupa modelul semiconservativ. Mecanismul replicarii s-a studiat la toate familiile de dezoxiribovirusuri.
Replicarea ADN la adenovirusuri
La adenovirusuri, genomul este o molecula dublu catenara, lineara, de 2000-2500 kDa (circa 36 kbp), cu secvente repetate invers, lungi de l00-l40 nucleotide, la ambele extremitati ale celor doua catene(secventa de baze la un capat al unei catene (de exemplu 5') este repetata in ordine inversa pe catena opusa, in acelasi sens (5'-3'). Ele permit circularizarea moleculelor monocatenare care apar in timpul replicarii. La cele doua capete 5', catenele genomice sunt asociate covalent cu o proteina de 55 kDa.
Replicarea ADN, catalizata de ADN-polimeraza proprie este semiconservativa si asimetrica. Asimetria consta in faptul ca numai una dintre catene este copiata intr-o catena complementara.
Replicarea se initiaza la capatul 3' al uneia dintre cele doua catene ale moleculei genomice parentale, la care se ataseaza o proteina de 80 kDa, cu o secventa asemanatoare de aminoacizi cu a proteinei de 55 kDa de la capatul 5'. Proteina de 80 kDa de la capatul 3' are rol de amorsa a replicarii. Sinteza ADN incepe cu adaugarea dCMP (deoxicitidin-monofosfat), la proteina amorsa, formand o legatura esterica. Apoi ADN-polimeraza* alungeste catena ADN in sensul 5' - 3', prin polimerizare succesiva la capatul 3'.
ADN-polimeraza nu initiaza sinteza de novo a unei catene de ADN, avand nevoie totdeauna de un primer, ci alungeste o catena preexistenta. De obicei, primerul este un fragment de ARN complementar fata de una dintre catene. Ulterior, secventa de ARN este inlocuita cu una de ADN.
La adenovirusuri, rolul de primer revine proteinei de 80 kDa, legata covalent la capatul 5'. Catena se alungeste in directia 5' - 3' pe catena matrita si nu necesita fragmente Okazaki. Catena nascenta deplaseaza catena preexistenta de aceiasi polaritate si impreuna cu catena matrita de polaritate opusa, formeaza o molecula dublu catenara.
Catena deplasata are rol de matrita pentru sinteza unei catene complementare. Dupa acelasi mecanism, se formeaza o molecula dublu-catenara, lineara.
Virusurile din grupul Papova au genomul format dintr-o molecula de ADN dublu catenara, circulara. Replicarea moleculei circulare se realizeaza dupa modelul semiconservativ, bidirectional si este simetrica. Dupa acelasi mecanism se replica ADN bacterian, precum si ADN din organite (mitocondrii, cloroplaste).
Replicarea este precedata de despiralizarea celor doua catene, ca rezultat al inciziei succesive. Inciziile sunt reparate rapid dupa ce s-a produs despiralizarea.
Enzimele care incizeaza si repara rapid bresele moleculei de ADN se numesc topoizomeraze. Rolul lor este acela de a relaxa ADN supraspiralizat in sens pozitiv sau negativ. Ele produc modificari ale configuratiei spatiale (topologice) a moleculei de ADN, prin modificarea gradului de spiralizare.
Virusurile Papova nu au topoizomeraze proprii, ci utilizeaza enzimele celulare.
Replicarea moleculei circulare incepe la un punct fix, denumit originea replicarii, iar punctul in care catenele moleculei parentale se separa si sunt sintetizate catenele noi se numeste bifurcatie de replicare.
De la origine, bifurcatia de replicare se deplaseaza cu foarte putine exceptii, in ambele directii ale moleculei circulare. Din aceasta cauza, o molecula de forma circulara, in cursul procesului de replicare are aspectul literei grecesti theta ( ), iar mecanismul de replicare se numeste theta.
Rezultatul replicarii este o pereche de molecule circulare, denumite catenani. Catenarea este procesul de legare a doua molecule circulare de ADN intr-o structura supramoleculara, iar decatenarea este procesul invers, de separare a celor doua molecule.
La herpesvirusuri, genomul este o molecula de ADN dublu catenara, lineara, de l50 kbp. Molecula este alcatuita din doua domenii cu secventa unica: unul mai mare, notat cu L (Large), si altul mai scurt, S (Short).
La Herpes simplex virus (HSV), fiecare secventa este flancata la extremitati, de cate o secventa de nucleotide repetate in ordine inversata (SRI), de cate 300-400 de baze. O particularitate a genomului HSV este orientarea intamplatoare a celor doua secvente unice, astfel incat intr-o populatie de virioni se gasesc simultan cele 4 variante de genom linear: P (Prototip), IL (Inverted L), IS (Inverted S) si ISL (Inverted S si L ). Ele apar ca o consecinta a asocierii aleatorii in oricare dintre cele doua orientari a celor doua domenii genomice.
Replicarea genomului se face prin intermediul moleculelor circulare. Dupa ce ADN viral ajunge in nucleul celulei, secventele terminale repetate in ordine inversa (SRI) sunt supuse actiunii limitate a unei exonucleaze, la nivelul uneia dintre catene. Secventele ramase se imperecheaza, formand o molecula circulara, dublu catenara, covalent inchisa.
Intr-o prima etapa, replicarea moleculei circulare se face ciclic, dupa modelul semiconservativ al cercului simplu. Ulterior, moleculele circulare generate in prima etapa a transcrierii se replica dupa modelul cercului rotativ. Un rol esential in procesul de replicare il au helicazele*.
Helicazele sunt enzime care catalizeaza despiralizarea catenei duble a acizilor nucleici, prin disocierea legaturilor de H si utilizeaza E eliberata prin hidroliza unui nucleotid 5'-trifosfat (NTP), de obicei ATP. Energia eliberata in timpul hidrolizei NTP este utilizata pentru derularea AN, desi nu se cunoaste modalitatea in care se cupleaza cele 2 reactii.
Helicazele ADN s-au descris cu peste 20 de ani in urma, dar helicazele ARN s-au evidentiat recent la virusuri, la bacterii, la levuri, pana la om. La virusuri, helicazele au raspandire larga, cu exceptia retravirusurilor si a celor cu genom de polaritate negativa.
Modularea structurii ARN este o etapa esentiala in numeroase procese fundamentale: sinteza ARN, transcrierea, replicarea si traducerea. La virusuri, helicazele au rol esential in ciclul de replicare. Ele deruleaza ARN si ADN dc, dar si heteroduplexul ARN-ADN in timpul replicarii, transcrierii si traducerii genomului viral. Helicaza ar fi necesara oricarui virus, indiferent de natura genomului sau, datorita existentei nucleotidelor complementare.
La virusurile cu genom d c, ARN sau ADN, helicaza este necesara disocierii catenelor in timpul replicarii si transcrierii. La ribovirurile monocatenare, genomul se gaseste tranzitoriu sub forma dc: in timpul replicarii ARN de polaritate pozitiva, cand aparatul de replicare trebuie sa functioneze, iar matrita libera trebuie sa fie permanent disponibila pentru un nou ciclu de replicare. Rolul helicazei ar fi acela de a disocia perechile intramoleculare de baze si de a preveni formarea perechilor extinse intre matrice si catena complementara nascenta. Virusurile cu genom mai mic de 5,8 kb nu codifica helicaza.
Una dintre catenele moleculei circulare (catena leading*) este incizata la un situs specific, sub actiunea unei endonucleaze virale si rezulta doua capete libere: 3'OH si 5'P. Cealalta catena ramane circulara, inchisa covalent.
La capatul 3' al catenei incizate (catena "leading") este initiata sinteza replicativa. Capatul 3'OH are rol de primer si creste prin polimerizarea nucleotidelor, catalizata de ADN-polimeraza. Catena circulara functioneaza ca matrita rotativa.
Polimerizarea are loc totdeauna, prin alungirea unei catene primer in directia 5'- 3', incepand de la capatul 3'OH al catenei incizate. Cand bifurcatia de replicare a parcurs cel putin odata catena circulara cu rol de matrita si catena incizata (leading*) este de doua ori mai lunga in raport cu momentul initial, ea poate fi copiata in sens invers, pe calea replicarii discontinue a fragmentelor Okazaki, de un alt complex enzimatic de replicare. Astfel, molecula nou sintetizata devine dublu catenara.
Catena "leading"(to lead, englez = a conduce) este cea care expune capatul 3'OH liber si este transcrisa intr-o catena complementara continua in directia 5' -- 3'.
Catena leading continua sa se roteasca pe matrita circulara, fiind copiata succesiv. Se formeaza astfel mai multe molecule concatemere, in tandem, in conexiune cap-coada.
Rezultatul intermediar al replicarii este un cerc cu o ramificatie laterala, care se aseamana cu litera greceasca sigma ( ) si de aceea, replicarea dupa modelul cercului rotativ se numeste replicare sigma.
Concatemerul poate fi separat de molecula circulara prin clivare, sub actiunea unor endonucleaze. La randul lor, capetele catenei incizate ale moleculei circulare sunt reunite sub actiunea unei ligaze si se reface continuitatea covalent inchisa a structurii circulare dublu catenare.
In procesul morfogenezei virale, concatemerii lineari sunt clivati in molecule dublu catenare lineare, de lungimea genomului.
Replicarea dupa modelul cercului rotativ are urmatoarele caracteristici: a)catena leading este legata covalent de catena parentala; b)replicarea repetata pe matrita circulara genereaza o ramificatie laterala concatemera, legata covalent.
Parvovirusurile au ca genom o molecula de ADN monocatenara, lineara. Capetele moleculei au secvente palindromice*, repetate in ordine inversa, care formeaza "bucle in ac de par". Regiunile palindromice terminale reprezinta 5% din genom si sunt esentiale pentru replicarea acestuia, deoarece constituie secventele primer pentru actiunea catalitica a ADN-polimerazei. Replicarea este catalizata de aparatul enzimatic al celulei.
Intr-o prima etapa, genomul monocatenar este convertit la forma dublu catenara, prin imperecherea bazelor complementare, denumita intermediar de replicare.
Intermediarul dublu catenar este incizat intr-o pozitie opusa situsului de initiere pentru sinteza catenei complementare si genereaza un capat 3'OH, de la care secventa repetitiva este alungita pe catena complementara cu rol de matrita. Se sintetizeaza astfel, catene multiple cu polaritate genomica (pozitiva). Catenele nou sintetizate, cu polaritate genomica, raman in configuratie lineara si constituie genomul virionilor progeni sau se replica dupa acelasi mecanism, amplificand rata sintezei catenelor genomice.
*Palindromul este o succesiune de baze, simetrica fata de un punct central, astfel incat secventa bazelor este aceiasi atat in sensul 3' --- 5', cat si in sensul 5' --- 3'.
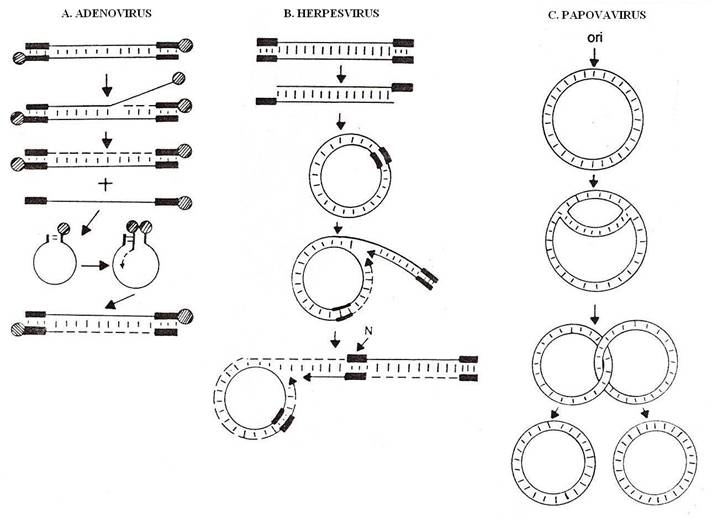
Fig. 88. Reprezentarea schematica a replicarii genomului ADN viral.
Replicarea si transcrierea genomului viral ARN
In citoplasma celulelor animale si umane, activitatea ARN-polimerazei nu este detectabila. Toate ARN-polimerazele dependente de ARN din celulele animale sunt codificate de ribovirusuri.
De cele mai multe ori, la ribovirusuri, procesele de replicare si transcriere nu sunt separate in timp si spatiu, ci interfera, datorita faptului ca ARN genomic functioneaza alternativ ca matrita pentru copierea replicativa a ARN genomic sau pentru transcrierea ARN mesager.
Replicarea genomului ARN monocatenar se face totdeauna dupa urmatorul model: catena genomica este convertita la forma dublu catenara, sub actiunea catalitica a unei enzime de replicare (o ARN-polimeraza dependenta de ARN), de origine virala, prin sinteza unei catene complementare de aceiasi lungime. ARN-polimeraza se deplaseaza totdeauna in directia 3' - 5' a matritei si sintetizeaza ARN in directia de elongatie 5' - 3'. Se formeaza o structura dc, denumita forma de replicare. Forma dc de ARN este suportul sintezei replicative a ARN genomic si transcrierii ARNm. ARN-polimeraza nu necesita existenta unui primer pentru a initia sinteza catenei noi.
Sinteza noilor catene are loc dupa mecanismul semiconservativ asimetric. Din forma de replicare dublu catenara, catena genomica este deplasata progresiv de catena noua, in curs de sinteza. Catenele nou sintetizate au aceiasi secventa de baze si aceiasi polaritate ca si catena genomica. Sinteza catenelor noi este succesiva si completa, adica sinteza catenei urmatoare nu incepe pana ce precedenta nu s-a eliberat.
Forma de replicare dublu catenara, impreuna cu o catena nascenta formeaza un intermediar de replicare.
Catenele nou sintetizate, cu polaritate genomica devin suportul sintezei replicative sau se asociaza cu proteinele capsidale.
La virusurile care au ca genom o catena de ARN cu polaritate pozitiva (Picorna, Togavirus), catena de sens pozitiv a formei dc este dislocata de catena nascenta a intermediarului de replicare, iar catena de sens negativ are rol de matrita pentru sinteza copiilor de polaritate pozitiva, care indeplinesc doua roluri:
a) unele, dupa prelucrarea prin bonetare*(metilare la capatul 5'), metilare interna si adenilare la capatul 3', au rolul de ARNm;
Bonetarea sau capping (cap, englez = boneta) semnifica o etapa a prelucrarii post-transcriere a moleculei de ARNm, ce consta in adaugarea la capatul 5' a metil-G. "Bonetarea" este esentiala pentru interactiunea moleculei de ARNm cu ribosomii in procesul traducerii informatiei genetice.
b) altele functioneaza ca matrite pentru sinteza unor catene noi, de polaritate negativa, amplificand rata sintezei ARN cu specificitate virala.
La virusurile cu genom ARN de polaritate pozitiva, deoarece ARN genomic functioneaza ca ARNm, ARN-polimeraza se sintetizeaza in celula, dupa infectie, si nu este componenta a virionului. Din aceasta cauza, ARN genomic este infectios (fig. 89).
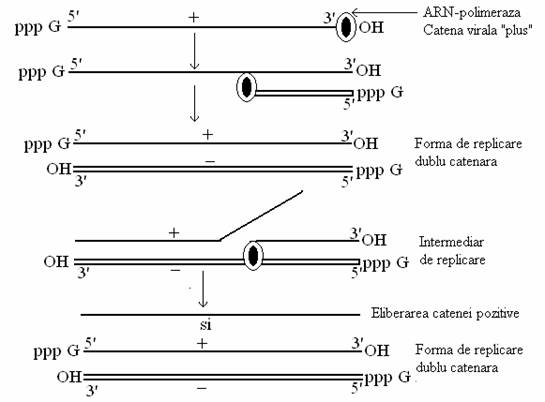
Fig. 89. Modelul replicarii genomului viral ARN cu polaritate pozitiva.
La virusurile cu genom ARN de polaritate negativa (grupul Mononegavirales- paramixo-, rhabdo-, filovirusuri), mesagerii si catenele genomice sunt transcrise de pe matrite diferite: catena de polaritate negativa a formei dublu catenare functioneaza ca matrita pentru transcrierea ARNm monocistronici, iar cea de polaritate pozitiva este transcrisa in copii de lungimea genomului (fig. 90), de polaritate negativa, cu rol de ARN genomic. Replicarea se deosebeste de transcriere prin modul in care progreseaza ARN-pol: continuu sau secvential.
Virionul contine o transcriptaza proprie - o ARN-polimeraza dependenta de ARN, care catalizeaza sinteza catenei de ARN complementar si a ARNm. De aceea, ARN purificat al acestor virusuri nu este infectios.
Virusurile cu genom ARN se multiplica in citoplasma, cu exceptia virusurilor gripale, la care, replicarea si transcrierea genomului au loc in nucleu. La mixovirusuri, transcrierea moleculelor cu rol de ARNm, ca si a celor genomice, este catalizata de aceiasi ARN-polimeraza virala, dar enzima este defectiva functional, deoarece alungeste o catena preexistenta, dar nu initiaza sinteza de novo*.
Polimeraza PB2 si proteina NS1 cliveaza capetele 5' bonetate ale mesagerilor celulari, pe care polimeraza PB1 le foloseste ca primeri pentru sinteza replicativa si pentru transcrierea genomului viral. Alta explicatie a dependentei de nucleu rezida in aceea ca mesagerii pentru sinteza proteinelor M1 si M2, respectiv NS1 si NS2 sunt prelucrati prin clivare si inadire.
Unele copii de polaritate pozitiva, cu rol de ARNm sunt "bonetate" (capping), metilate si poliadenilate de enzime celulare nucleare si sunt transferate in citoplasma, iar altele raman in nucleu, cu rol de matrita pentru sinteza ARN genomic (de polaritate negativa), pe toata durata procesului infectios.
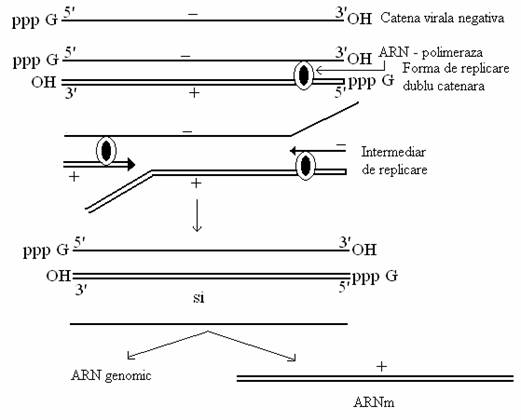
Fig. 90. Modelul replicarii genomului viral ARN cu polaritate negativa.
Replicarea genomului ARN dublu catenar multipartit al reovirusurilor* (l0 segmente) este simultana cu transcrierea (fig. 91).
Reovirusurile se transmit pe cale orala si sunt prelucrate de proteazele digestive in lumenul intestinal. Dezvelirea implica partial, digestia proteolitica a capsidei externe. Proteoliza nu inactiveaza virusul, ci remodeleaza capsida externa. Virusul este activat, dupa eliberarea proteinei majore 3, clivajul proteinei 1c si a HA δ1. Virusul prelucrat proteolitic adera de celulele M si de enterocite. Rezulta particule subvirale, identice cu cele obtinute in vitro sub actiunea chimotripsinei
Catena de polaritate negativa a fiecarui segment genomic este transcrisa de o ARN-polimeraza virala, in capsida partial deschisa. Se sintetizeaza l0 tipuri de ARN cu polaritate pozitiva, care parasesc capsida prin dizlocarea capsomerelor de la varfurile icozaedrului. Transcrierea este conservativa, deoarece ambele catene ale ARN genomic raman in regiunea centrala, nedezvelita a virionului.
Moleculele de ARN au doua functii:
a) sunt traduse in mesaje monocistronice si astfel se sintetizeaza proteine virale;
b) se asambleaza cu o precapsida si au rolul de matrite pentru sinteza catenelor complementare, de polaritate negativa. Catenele perechi constituie genomul viral.
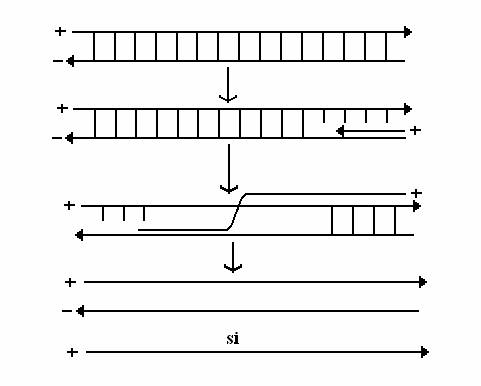
Fig. 91. Modelul replicarii genomului ARN dublu catenar al reovirusurilor.
Replicarea ARN genomic printr-un intermediar ADN este caracteristica retravirusurilor. Genomul lor este reprezentat de o catena de ARN, care, in fiecare virion se gaseste in dublu exemplar, ceea ce confera caracterul diploid al genomului acestor virusuri. Nu se cunoaste modul de asociere fizica a celor doua molecule de ARN.
Genomul ARN al retravirusurilor este replicat printr-un flux de informatie genetica ce trece prin ADN, proces denumit transcriere inversa* (fig. 92).
Fenomenul a fost descoperit de H. Temin si D. Baltimore (1970). Ei au tratat celulele infectate cu VSR, cu actinomicina D, un antibiotic care se insera intre bazele nucleotidice si inhiba transcrierea. S-a remarcat astfel ca actinomicina D, inhiba ciclul de multiplicare al retravirusurilor, dar nu inhiba multiplicarea ribovirusurilor obisnuite. Concluzia a fost ca in ciclul lor de multiplicare exista o faza sensibila la actinomicina D, reprezentata de un intermediar ADN.
Singura functie a ARN genomic este de a indeplini rolul de matrita pentru sinteza intermediarului de ADN in celula infectata. Celula nu poseda o enzima care sa indeplineasca aceasta functie. Virionul contine o ADN-polimeraza dependenta de ARN, denumita revers-transcriptaza (RT), precum si molecule de ARNt incorporate din celula in procesul asamblarii, care indeplinesc rolul de primer al transcrierii inverse, deoarece ADN-polimeraza nu initiaza sinteza unei catene noi, ci doar alungeste una preexistenta.
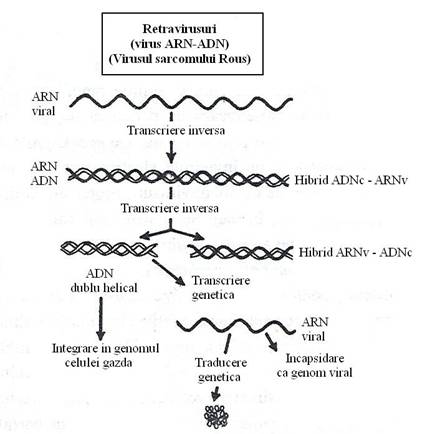
Fig. 92. Succesiunea etapelor transcrierii si traducerii informatiei genetice la retravirusuri.
Procesul transcrierii incepe timpuriu, deoarece RT se activeaza imediat ce virusul a ajuns in celula sensibila. Etapele replicarii ARN viral sunt urmatoarele:
a) Legarea complexului molecular al RT, de ARN genomic viral;
b) Sinteza unei copii de ADN complementar, catalizata de RT, care ramane legata prin punti de H, de ARN genomic. Rezulta un hibrid molecular ARN-ADN;
c) Digestia selectiva a ARN genomic, din hibrizii moleculari ARN-ADN, sub actiunea RN-azei H, sau chiar sub actiunea RT;
d) Sinteza unei catene complementare de ADN viral. Astfel se formeaza o molecula dublu catenara de ADN viral, care este translocata in nucleul celulei infectate si se integreaza covalent in structura unui cromosom al celulei. ADN viral este integrat stabil si din punct de vedere structural nu se distinge de secventele de ADN cromosomal.
ADN integrat este transcris in doua tipuri de molecule de ARN: unele scurte (subgenomice), cu rol de ARNm si altele lungi, cu functie genomica. La retravirusurile transductoare, copiile lungi de ARN incorporeaza secvente oncogene (protooncogene), derivate din celula gazda.
Activitatea ADN-polimerazica a RT necesita o matrita de ARN sau ADN si, ca toate ADN-polimerazele cunoscute, nu initiaza sinteza ADN de novo, ci necesita o molecula preexistenta (primer). In vitro, primerul poate fi ARN sau ADN, dar in vivo, totdeauna este ARN.
Pentru sinteza catenei ADN de polaritate negativa, rolul de primer il are molecula de ARNt, originara in celula. Pentru sinteza celei de a II-a catene de ADN, molecula de ARN genomic este incizata la nivelul secventei polipurinice (PP) si functioneaza ca primer.
Toate revers-transcriptazele, pentru activitatea in vitro au nevoie absoluta de cationi bivalenti: Mg2+ (l0 mM).
Activitatea RN-azei H este inerenta in toate preparatele de revers-transcriptaza. Ea hidrolizeaza ARN-genomic din hibridul ARN-ADN, pe masura progresiei celei de a II-a catene de ADN.
Biosinteza proteinelor tardive
Categoria proteinelor tardive cuprinde pe acelea care se sintetizeaza dupa replicarea genomului viral. Ele sunt in primul rand, proteine structurale, necesare asamblarii virionilor progeni si se sintetizeaza cu o rata inalta, prin transcrierea si traducerea copiilor genomice multiple, rezultate prin replicare. De aceea, proteinele tardive se sintetizeaza in cantitati mari, usor detectabile pe imaginile electrono-optice sau la analiza electroforetica. De exemplu, in celulele infectate cu adenovirusuri, proteinele tardive se sintetizeaza in mare exces si formeaza incluziuni mari, in care agregatele moleculare au o distributie ordonata, paracristalina. Alte proteine tardive au rol functional:
- proteinele reglatoare, al caror rol s-a demonstrat mai intai pentru bacteriofagi, iar ulterior, pentru adeno- si herpesvirusuri. Cantitatile de proteine necesare asamblarii diferitelor structuri virale sunt foarte diferite si proportiile sintezei trebuie corelate direct cu necesarul;
proteine de morfogeneza, necesare asamblarii virusurilor cu structura complexa;
proteine care usureaza eliberarea virionilor din celula.
Pentru sinteza proteinelor tardive, virusurile folosesc aparatul celular de sinteza si exploateaza mecanismele celulare pentru transportul si modificarea proteinelor traduse.
Ca si proteinele celulare, cele virale se sintetizeaza pe ribosomii legati de membrane sau pe ribosomii liberi in citoplasma, in functie de destinatie. In celula normala, ribosomii asociati membranelor, sintetizeaza proteine si glicoproteine de membrana, iar ribosomii liberi sintetizeaza enzime si alte proteine citoplasmatice.
Transportul si prelucrarea proteinelor
Pentru localizarea diferita a proteinelor este determinanta secventa de aminoacizi de la capatul N-terminal al polipeptidului, denumita secventa semnal, dar si alte secvente ale polipeptidului, ca si modificarile ce survin dupa sinteza. Toate orienteaza proteina spre diferite localizari: intrarea in nucleu, insertia in membrana etc. Determinantii care orienteaza destinatia proteinei se numesc semnale de triere. Semnalele au caracter de specificitate si constau din scurte secvente de aminoacizi care se adauga polipeptidului in cisternele aparatului Golgi. In functie de destinatie, proteinele vor fi segregate si impachetate in vezicule separate.
Adeseori, proteinele virale se sintetizeaza sub forma unor precursori de dimensiuni mai mari, care sunt supusi clivajului proteolitic, fie in timpul sintezei, fie ulterior. Clivarile sunt catalizate de proteaze celulare si mecanismul lor este similar cu acela al clivarii proteinelor celulare. Astfel, proteinele virale cu destinatie membranara pierd secventa N-terminala, inainte de terminarea sintezei. Proteinele structurale sunt, adeseori, clivate in timpul asamblarii virionilor.
Glicozilarea proteinelor virale, consecutiva traducerii informatiei genetice urmeaza acelasi mecanism ca si glicozilarea proteinelor celulare. Toate proteinele expuse pe suprafata invelisului sunt glicozilate, iar cele din structura nucleocapsidei nu sunt niciodata glicozilate. Catenele glucidice ale glicoproteinelor virale si celulare, formate din l-3 resturi, sunt N-lincate de asparagina si contin secventa Man-GlcNac-GlcNac. Glicozilarea incepe in timpul sintezei polipeptidului in reticulul endoplasmic.
Catenele glucidice O-lincate sunt legate de treonina si serina si sunt adaugate totdeauna in cisternele Golgi. Diferitele zaharuri ale catenei oligozaharidice sunt adaugate secvential, fiecare reactie fiind catalizata de o glicozil-transferaza specifica de origine celulara.
Unele proteine virale sunt fosforilate. Ele interactioneaza cu ADN si au rol in replicarea ADN sau in transcriere. Altele sunt acilate, prin legarea covalenta a acizilor grasi.
Proteinele virale tardive sunt transportate la situsuri diferite in functie de sediul morfogenezei virionilor: o parte ramane in celula, iar o alta parte este transportata la nivelul membranei, avand rol in procesul inmuguririi virale.
Transportul glicoproteinelor de invelis este cunoscut la mixo- si paramixovirusuri. Ele se sintetizeaza pe poliribosomii atasati cisternelor de reticul endoplasmic. De aici migreaza in cisternele golgiene si se glicozileaza sau sufera alte modificari (acilare cu acid palmitic sau miristic). Prin vezicule de transport, glicoproteinele sunt transportate la nivelul membranei si se insera pe fata externa a acesteia. Proteina matriceala (M), neglicozilata, se agrega prin interactiuni necovalente, pe fata interna a membranei citoplasmatice, in zone distincte ("petice").
Asamblarea (morfogeneza) virala
Studiul dinamicii asamblarii virusurilor infectioase pentru celula animala este ingreunat, pe de o parte, de complexitatea structurala a capsidei, iar pe de alta, de abundenta componentelor moleculare ale mediului celular in care are loc asamblarea. Cunostiintele referitoare la mecanismele dominante ale asamblarii virale sunt rezultatul studiilor morfogenezei fagilor si a virusurilor infectioase pentru plante.
Morfogeneza virala se supune principiului fundamental al autoasamblarii*, ca rezultat al faptului ca fiind determinante de forma, capsomerele se pot asocia intr-o modalitate unica.
Autoasamblarea semnifica faptul ca toata energia si informatia pentru asamblare sunt continute in monomerii individuali. Monomerii se asociaza unul cu altul, intr-o ordine determinata, intr-o combinatie de legaturi ionice, legaturi de H si interactiuni hidrofobe.
La virusurile cu simetrie helicala (de exemplu, virusul mozaicului tutunului - VMT), capsida se asambleaza pe masura ce capsomerele interactioneaza cu ARN genomic (fig. 93). La acest virus s-a demonstrat pentru prima data, existenta unei relatii fixe intre genom si capsomere, precum si mecanismul autoasamblarii. Genomul VMT este alcatuit din 6390 de nucleotide. Daca fiecare capsomera, in procesul morfogenezei capsidei se asociaza cu trei nucleotide, rezulta ca lungimea genomului viral, determina in mod obligatoriu, lungimea virionului. Altfel spus, in timpul asamblarii virionului, capsomerele se aseaza in jurul moleculei de ARN genomic. In absenta genomului, capsomerele se asambleaza in numar foarte mare, formand o capsida imensa, dar goala.
La VMT, molecula proteica este in acelasi timp, unitate chimica si unitate structurala a capsidei (capsomera). In anumite conditii de mediu (cresterea valorii pH), capsida se dezorganizeaza in capsomerele componente. Restabilirea conditiilor de mediu, induce autoasamblarea capsidei cilindrice, deoarece capsomerele sunt determinante de forma, adica se pot asambla intr-un singur mod, dupa principiul de constructie a simetriei helicale.
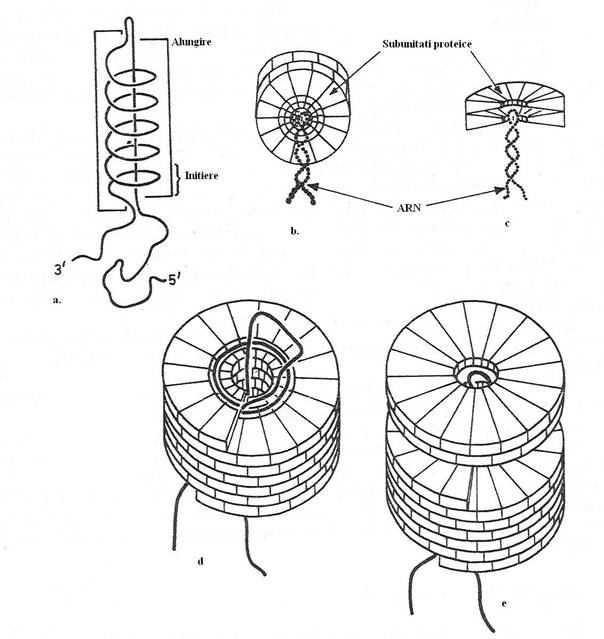
Fig. 93. Procesul autoasamblarii semnifica interactiunea genomului cu capsida. a. La VMT, structura in ''ac de par'' a genomului cu regiunea de initiere, se leaga de primul disc dublu format din capsomere pentru a initia asamblarea virionului. b, c. Insertia ARN viral in scobitura centrala a discului dublu format din capsomere. d, e. Alungirea virusului se realizeaza prin adaugarea succesiva a discurilor duble de capsomere.
Mecanismul autoasamblarii capsidei VMT este urmatorul: initial se formeaza un complex stabil, de l7 unitati proteice (capsomere), in forma de disc, asezate intr-un strat unic. Ele se agrega spontan pentru a forma un disc alcatuit din doua straturi ce contin 34 capsomere. Fiecare strat al discului contine l7 capsomere, aproape acelasi cu numarul de capsomere (l6,3), intr-un tur al helixului VMT. Discul alcatuit din doua straturi de capsomere este intermediarul cheie al asamblarii VMT. O proprietate importanta a discului este ca subunitatile pot sa alunece una fata de alta, pentru a forma un helix cu doua tururi.
Discul, alcatuit din doua tururi de capsomere interactioneaza cu ARN genomic. Asamblarea incepe prin insertia buclei de initiere a genomului (secventa de impachetare), in spatiul liber din centrul discului proteic. Discul proteic recunoaste specific secventa de impachetare a genomului. Cu exceptia secventei de impachetare, care initiaza procesul asamblarii, proteinele capsidei si ARN genomic interactioneaza nespecific. Prin interactiunea lor nu se formeaza legaturi covalente noi, ci numai legaturi secundare slabe. Ca dovada a interactiunii laxe, atat intre capsomere, cat si intre capsomere si ARN, capsida se disociaza prin cresterea valorii pH.
Virionul este complet asamblat dupa ce este acoperit capatul 5' al genomului.
In general, virionii cu structura complexa se asambleaza gradat din ansambluri distincte, construite din subunitati proteice, ce se formeaza spontan prin autoasamblarea moleculelor proteice individuale.
In esenta, asamblarea virionilor cu simetrie icozaedrica, este rezultatul a doua procese ce se desfasoara succesiv sau aproape simultan:
asamblarea proteinelor structurale in capsomere si a acestora in structura capsidei. La virusurile complexe, capsomera ca unitate de constructie este alcatuita din cateva molecule proteice;
asocierea capsidei cu acidul nucleic genomic.
La ribovirusurile nude, cu simetrie icozaedrica, asamblarea capsidei si asocierea ei cu ARN sunt procese aproape concomitente, datorita instabilitatii ARN viral in celula.
La virusurile ADN cu simetrie icozaedrica (adeno-, herpetice), ansamblurile proteice pot forma o structura goala, matura, denumita precapsida, inainte de a se asocia cu genomul. Interactiunea genomului viral (ADN sau ARN), cu proteinele capsidale si rolul acestor interactiuni in impachetarea genomului raman necunoscute. In capsida este impachetat, de regula, numai ARN genomic, ca dovada a existentei unei secvente de impachetare (identificata la ARN Sindbis - un alfavirus).
In procesul asamblarii capsidelor cu simetrie icozaedrica, participa proteinele de "cofraj". Ele au probabil, nu numai un rol mecanic, ci si catalitic in realizarea unor interactiuni stabile ale moleculelor componente. Dupa asamblarea capsidei si consolidarea interactiunilor intre capsomere, proteinele de "cofraj" sunt eliminate din structura virionului.
La reovirusuri, segmentele genomice de ARN dublu catenar se asambleaza cu o proteina de morfogeneza si formeaza regiunea centrala a virionului, pe suprafata careia se asambleaza capsida.
Studiul ultrastructural al replicarii virusului vaccinal a relevat procesul stadial complex al morfogenezei, cu participarea cisternelor aparatului Golgi, care formeaza invelisul extern al virionilor. In citoplasma celulelor infectate apar zone difuze sau bine delimitate alcatuite din material fin-granular, denumite viroplasma, in interiorul carora se asambleaza virionii progeni. Procesul asamblarii, complex si incomplet elucidat, comporta edificarea de novo a unor structuri membranare destinate sa inglobeze o parte din materialele viroplasmei.
Cresterea, turnover-ul si repararea membranelor se face prin intercalarea componentelor structurale (proteine, lipide) in structurile preexistente. Biogeneza de novo a membranelor ramane una dintre marile probleme ale biologiei. Pentru virusurile pox, exista dovada structurala ca unele componente ale invelisului, sintetizate sub control viral, incep sa se condenseze in membrane. Membranele sunt asamblate in focare viroplasmice discrete si raman separate de membranele celulare.
Membranele virale sintetizate de novo sunt detectabile in matricea viroplasmei si au structura trilaminara, fiind tapetate la exterior de un strat dens de spiculi. Pe masura ce membranele cresc, ele inconjura ADN nascent si proteinele acumulate in viroplasma.
Eliberarea virionilor
Eliberarea virusurilor din celula se face prin mai multe mecanisme, dependente in primul rand, de natura virusului.
Virusurile nude se elibereaza rapid dupa moartea si liza celulei. Intr-o mica masura, virusurile nude (cu genom ARN sau ADN) se elibereaza prin modificarea permeabilitatii membranei citoplasmatice.
Ribovirusurile invelite (mixo-, paramixo, toga-, retravirusuri) se elibereaza prin inmugurire din membrana citoplasmatica sau in cisternele reticulului endoplasmic granular (flavivirusuri). Ultimele etape ale asamblarii nucleocapsidei sunt asociate cu primele stadii ale inmuguririi, sau asamblarea capsidei precede inmugurirea (de exemplu, particulele de tip B si D ale retravirusurilor). Inmugurirea are loc la nivelul unor situsuri specifice ale membranei celulare, acolo unde proteinele virale s-au inserat, atat pe fata interna cat si pe cea externa a membranei plasmatice sau a membranelor reticulului endoplasmic. Asamblarea prin inmugurire necesita co-localizarea componentelor structurale (fig. 94).
Proteinele care proemina pe suprafata externa a membranei sau in cisternele reticulului endoplasmic, sunt glicozilate, iar cele care tapeteaza fata interna a membranei citoplasmatice sunt neglicozilate. Glicoproteinele virale se insera in zone distincte ale membranei, din care proteinele celulare sunt deplasate sau chiar inlocuite. Nucleocapsidele se asociaza specific cu proteinele virale care tapeteaza fata citoplasmatica a membranei (proteina M), sau cu domeniile citoplasmatice ale glicoproteinelor virale inserate in membrana (la togavirusuri).
La ortho- si paramixovirusuri, nucleocapsidele recunosc specific si se ataseaza de proteina matriceala. Ulterior incepe inmugurirea si treptat, virionul se elibereaza, invelit fiind de membrana citoplasmatica, in care se gasesc inclavate glicoproteinele virale. Proteina matriceala are rolul unei punti de legatura intre nucleocapsida virala si glicoproteinele inserate in membrana.
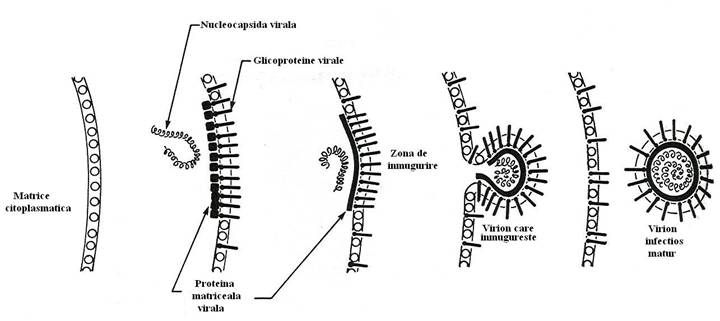
Fig. 94. Formarea invelisului viral prin inmugurire. Eliberarea particulelor virale progene.
Lipidele si glucidele din alcatuirea invelisului viral sunt proprii celulei. Din aceasta cauza, compozitia lor in structura unui virus difera in raport cu substratul celular in care se multiplica. Datorita acestui fapt, preparatele unui virus, obtinute pe linii celulare diferite se deosebesc prin proprietatile fizice, biologice si antigenice.
Unele virusuri care inmuguresc, nu sunt foarte eficiente in procesul eliminarii proteinelor membranare, iar fenomenul incorporarii proteinelor straine este real. De aceea, invelisul viral contine si glicoproteine de origine celulara si chiar ale unui alt virus, care infecteaza concomitent aceiasi celula. Faptul este ilustrat de neutralizarea retravirusurilor, atat in vivo cat si in vitro, de anticorpii specifici fata de proteinele membranare. Proteinele membranare par a fi incorporate pasiv in invelis, daca nu impiedica steric formarea acestuia.
Eliberarea virionilor prin inmugurire este mai eficienta pentru randamentul infectiei (producerea virusului progen), deoarece nu depinde de dezintegrarea imediata a celulei. Celula ramane viabila o perioada mai lunga de timp sau dupa transformarea maligna este chiar nemuritoare si elibereaza virus.
Efectele virusurilor care inmuguresc, asupra celulei gazda sunt foarte diverse: de la citoliza (toga-, paramixo, rhabdovirusuri), pana la supravietuirea nelimitata a acesteia, dupa transformarea cu retravirusuri.
La herpesvirusuri, virionii se invelesc la contactul cu lamela interna a membranei nucleare, traverseaza spatiul dintre cele doua lamele si la nivelul porilor nucleari trec in cisternele reticulului endoplasmic si in vezicule. Sistemele membranare inchise protejeaza virionii de contactul cu citoplasma si mediaza transportul la suprafata celulei. Membrana veziculei de transport, fuzioneaza cu membrana citoplasmatica si virionii sunt eliberati in spatiul extracelular. Herpesvirusurile sunt totdeauna citolitice.
Celulele infectate cu virusuri care inmuguresc la nivelul membranei citoplasmatice sau la nivelul altor membrane celulare, devin tinta antigenica pentru efectorii sistemului imunitar, care recunosc glicoproteinele codificate de virus. Cele infectate cu virusuri nude sunt recunoscute de efectorii sistemului imunitar, deoarece expun antigene virale asociate cu molecule membranare.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 2825
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved