| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Conjugarea bacteriana
Conjugarea este o modalitate de transfer de material genetic de la o celula donoare, la una receptoare, prin intermediul unei legaturi celulare directe. Procesul este conditionat de existenta, in celula donoare, a unei plasmide cu functie de conjugon (transferon sau plasmida conjugativa), care trebuie sa codifice structurile necesare realizarii contactului celular, iar pe de alta parte, sa pregateasca transferul in celula receptoare, adica sa fie o plasmida mobilizabila. Absenta uneia sau a ambelor functii, semnifica existenta plasmidelor neconjugative si/sau nemobilizabile.
Plasmidele conjugative sau plasmide de sex sunt factorii F, la care se adauga unele plasmide R si unele plasmide Col.
Fenomenul conjugarii bacteriene a fost descoperit de Lederberg si Tatum (l944). Ei au remarcat ca tulpinile dublu auxotrofe complementare de E. coli K12, dupa cultivarea in amestec in mediu nutritiv lichid, genereaza celule prototrofex, care cresc pe un mediu solidificat minimal si produc colonii.
*Tulpina de E. coli K12 este prototrofa, adica face sinteza tuturor constituientilor chimici, ceea ce ii permite sa creasca si sa se multiplice pe medii minimale, care nu contin factori de crestere (vitamine, aminoacizi). Se mai numesc si tulpini de tip salbatic, deoarece, din punct de vedere metabolic corespund celor existente in natura.
De la tulpinile prototrofe, sub actiunea mutagena a radiatiilor UV, se obtin mutante nutritionale, denumite auxotrofe (auxein, grec = strain), care au drept caracter marcant, incapacitatea de a face sinteza uneia sau mai multor substante necesare (esentiale). Prin urmare, substanta ce nu poate fi sintetizata, trebuie adaugata mediului de baza, ca o conditie obligatorie pentru ca bacteria respectiva sa se poata dezvolta. Substantele adaugate pentru a suplini incapacitatile de sinteza se numesc factori de crestere.
Lederberg si Tatum au utilizat in experientele lor, tulpini mutante dublu auxotrofe si triplu auxotrofe, cu incapacitatea de a sintetiza treonina, leucina, tiamina, biotina, fenil-alanina si cisteina. Aceste incapacitati de sinteza au fost luate ca markeri genetici pentru analiza ulterioara.
In exprienta cruciala, autorii au urmarit comportarea a doua tulpini mutante - A si B - triplu auxotrofe, dar selectionate astfel incat cele trei incapacitati de sinteza ale tulpinii A sa fie complementate de tulpina B si invers (sunt tulpini auxotrofe complementare) (fig. 131).
thr+ , leu+, thi+ thr--, leu-, thi-
Tulpina A bio-, phe-, cis- Tulpina B bio+, phe+, cis+
Cultivate izolat, pe mediul minimal, cele doua tulpini bacteriene nu cresc (nu formeaza colonii). In experienta propriu-zisa de conjugare, cele doua tulpini bacteriene (cate lo8 celule din fiecare), au fost puse in contact pentru un interval, in mediul lichid si ulterior au fost insamantate pe mediul minimal solidificat. Dupa 24 de ore se inregistreaza aparitia coloniilor prototrofe. In experienta de control, cultivarea tulpinilor A si B, separate, pe mediul minimal, nu genereaza aparitia coloniilor.
Toate coloniile crescute pe mediul minimal, din punct de vedere genetic sunt prototrofe: thr+, leu+, thi+, bio+, phe+, cis+, ca si cum cele doua tulpini auxotrofe au facut schimb de material genetic.
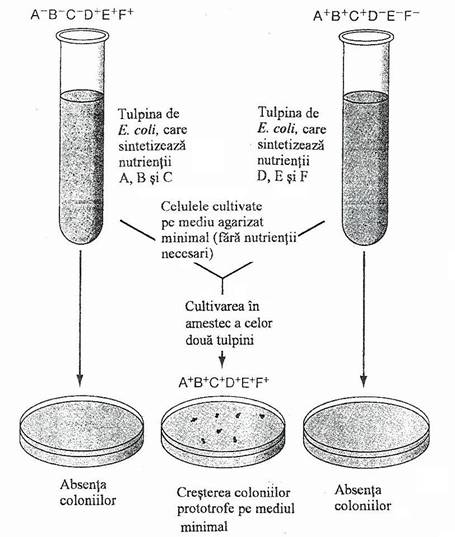
Fig. 131. Reprezentarea schematica a experimentului initial al lui Lederberg, care a condus la descoperirea procesului de conjugare.
Pentru a explica originea celulelor prototrofe s-a emis ipoteza retromutatiei. Retromutatiile in experiente de acest tip sunt fenomene rare: se produc cu o frecventa de l/l0-7 celule/24 de ore pentru un singur caracter. Pentru trei caractere, rata statistica a producerii retromutatiei, este de l/l0-21 celule/24 de ore, practic imposibila datorita caracterului triplu auxotrof al mutantelor.
Ipoteza reversiei la prototrofie prin transformare genetica, a fost eliminata prin experienta cultivarii celor doua tulpini triplu auxotrofe, intr-un tub in forma literei U. Cele doua brate ale tubului sunt separate printr-o membrana poroasa, permeabila pentru moleculele mediului de crestere, dar care nu permite trecerea celulelor dintr-un brat in celalalt al tubului. Atata timp cat celulele celor doua tulpini sunt fizic separate, reversia la prototrofie nu se realizeaza. Singura explicatie pentru geneza mutantelor prototrofe, in experienta evocata, ramanea posibilitatea unui contact celular direct, argumentata si de imagini la microscopul electronic, care sugereaza acest fapt.
Determinismul genetic al procesului de conjugare
Conjugarea este conditionata de prezenta in celula donor, a plasmidei F, care contine determinantii genetici tra (circa 28 de gene) si secventele de origine a transferului - ori T. Alte gene codifica sinteza pilinei, sinteza moleculelor de recunoastere intre celula donoare si receptoare, replicarea plasmidei. Cel putin alte 11 gene sunt necesare pentru asamblarea pilului functional.
Factorul F prezinta un grad important de omologie genetica cu cromosomul bacterian, in special pentru genele tra. Astfel s-a emis ipoteza ca la origine, genele factorului F ar fi fost cromosomale, dar s-au desprins si au devenit autonome.
In functie de prezenta si de raportul plasmidei F cu cromosomul, exista patru tipuri de celule, care se comporta diferit in procesul de conjugare:
celulele F- sunt acelea care nu au plasmida F. In procesul de conjugare, ele sunt receptoare de material genetic. Sunt echivalentii genetici ai organismelor de sex feminin;
celulele F+ poseda plasmida de sex in stare fizic autonoma. In procesul conjugarii, ele sunt donoare de material genetic. Sunt echivalentii genetici ai organismelor de sex masculin;
celulele Hfr (High frequency of recombination) prezinta plasmida F integrata in structura cromosomului. Aceste celule au capacitatea de a realiza cupluri de transfer, urmata de recombinare genetica, cu o mare frecventa. Fiziologic sunt considerate a fi celule "supermascul", donoare de material genetic;
celulele F' contin un factor F modificat prin procesul de recombinare cu cromosomul. Factorul F are proprietati episomale (se integreaza, reversibil, prin recombinare cu cromosomul). Dupa excizia incorecta, factorul F se desprinde impreuna cu un mic fragment de ADN cromosomal si devine factorul F'. In procesul de conjugare, celulele F' se comporta ca si celulele F+.
Etapele proceslui de conjugare
Conjugarea bacteriana este un proces foarte specific prin care ADN este transferat de la bacteria donoare la cea receptoare printr-un complex de proteine si organite care formeaza aparatul de conjugare. O conditie importanta pentru transferul conjugativ este asocierea intima a suprafetelor celor doua celule.
Initierea procesului de conjugare este conditionata de prezenta pililor, structuri care se evidentiaza dupa "marcajul" cu fagii care recunosc receptori piliari: la extremitatea libera a pililor se gasesc receptori pentru fagii filamentosi, iar pe suprafata pilului, pe toata lungimea sa se leaga fagii ARN "masculi", (de exemplu, MS2). Flagelii sunt structuri mai lungi si ondulate, iar fimbriile nu au receptori pentru fagi.
Dupa amestecul populatiilor de celule conjugante, in proportii corespunzatoare, intre celule se produc contactele (coliziunile) intamplatoare, urmate de formarea unor cupluri specifice de perechi, care la inceput sunt instabile, adica se pot dispersa printr-o agitatie usoara.
Cuplurile instabile se formeaza prin legarea extremitatii libere a pilului de o suprafata receptoare a celulei F-. Faza de legare instabila este urmata uneori, dupa cateva secunde, de faza de legare stabila. Cele doua celule sunt aduse in contact direct. Aceasta faza corespunde formarii perechilor eficiente. Asocierea celor doua celule este mult mai rezistenta la actiunea fortelor de dispersie. In aceasta etapa are loc transferul propriu-zis al materialului genetic, probabil prin formarea unui canal de conjugare. Cuplurile de transfer eficient, se formeaza la circa l5 minute, de la amestecul celulelor in proportii echivalente.
Rolul pililor in procesul de conjugare
Rolul pililor in procesul de conjugare este argumentat de mai multe fapte de observatie:
cultivarea bacteriilor in conditii de mediu, in care numarul pililor scade foarte mult, diminua corespnzator frecventa proceselor de conjugare;
pilii sunt prezenti numai la celulele care contin o plasmida cu functii conjugative;
indepartarea pililor prin agitare mecanica, este insotita de incapacitatea formarii cuplurilor de conjugare, pana cand pilii se refac;
blocarea suprafetelor receptoare ale pililor cu fagi ARN "masculi" sau a extremitatii libere cu fagi ARN filamentosi, diminua sau anuleaza capacitatea de formare a cuplurilor de conjugare;
pilul este un cilindru de 20 mm lungime, cu diametrul extern de 8 nm, iar cel intern de 2-2,5 nm, suficient pentru trecerea ADN monocatenar. Aranjarea moleculelor de pilina in structura pilului, este similara cu aceea a capsomerelor fagilor filamentosi;
la microscopul cu contrast de faza, se observa cupluri de celule care se deplaseaza sincron, separate de o distanta fixa;
la microscopul electronic cu baleiaj se observa structuri care leaga celulele cuplului.
Pentru rolul pilului in transferul de material genetic pledeaza rezultatele experientelor de conjugare intrerupta. Daca, dupa formarea cuplului, suspensia celulara este agitata si pilul este rupt, lungimea fragmentului de ADN care a trecut in celula receptoare este direct proportionala cu durata existentei pililor. Prezenta ADN in lumenul pilului nu s-a demonstrat cu certitudine.
A II-a ipoteza considera ca pilii ar avea un rol esential in faza initiala a procesului de conjugare, aceea a formarii cuplurilor. Pilul ar avea rolul unui carlig de "agatare" a celulei receptoare, dupa care ar urma retractia sa in celula donoare, printr-un proces de dezasamblare la baza. Exista dovezi ca interactiunea finala intre celulele cuplului este un canal de conjugare, asamblat de proteinele celulei donoare.
Studiul electrono-optic al unor celule Gram negative asociate tecii cianobacteriilor, a evidentiat un canal intercelular, delimitat de membrana citoplasmatica si de peretele celular (fig. 132). Lumenul canalului are un continut de aceiasi textura cu a materialului nuclear.
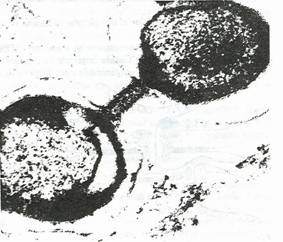
Fig. 132. Imaginea electrono-optica a unui posibil canal de conjugare la bacteriile Gram pozitive. Continutul canalului este asemanator nucleoplasmei (original).
Transferul ADN plasmidial in cuplul F+ x F-
Plasmidele conjugative pot sa transfere o copie a lor, de la celula donoare la celula receptoare. Transferul plasmidei F se realizeaza cu o frecventa mare. De exemplu, daca o singura celula de E. coli F+ se adauga la o cultura de celule F- , aflate in faza de crestere, dupa l5-20 de generatii, o mare proportie a celulelor contine factorul F, ca urmare a transferului unei copii a plasmidei de la celula F+ la celula F-, fara ca factorul F sa fie pierdut din celulele donoare. Fiecare celula receptoare dobandeste o plasmida F si la randul ei devine donoare. Intre doua diviziuni, o celula F+ realizeaza 1-2 transferuri, astfel ca factorul F se raspandeste repede in populatia bacteriana receptoare. Fenomenul se numeste "masculinizare epidemica" a celulelor F-.
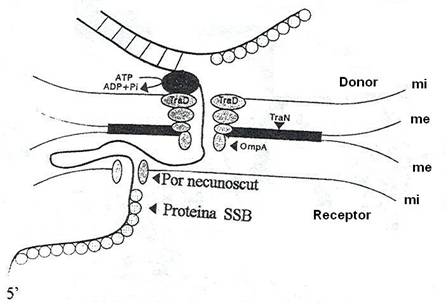
Fig. 133. Transferul ADN monocatenar al factorului F prin membranele celulei donor si receptor in timpul conjugarii. Canalul transmembranar al celulei donor ar putea fi asamblat din proteinele de transfer (mi = membrana interna ; me = membrana externa) (dupa Dreiseikelmann, 1994).
Dupa formarea cuplurilor eficiente, transferul conjugativ al ADN are loc in trei etape:
mobilizarea ADN pentru transfer
transferul propriu-zis
formarea unei plasmide functionale (cu capacitate de replicare in celula receptoare).
Etapele conjugarii se succed numai daca in celula donoare se gaseste o plasmida conjugativa si mobilizabila. Plasmidele conjugative poarta gene ce determina formarea perechilor eficiente, iar cele mobilizabile pregatesc ADN pentru transfer. Plasmidele autotransmisibile (de exemplu, plasmida F) sunt atat conjugative cat si mobilizabile.
O celula poate avea doua plasmide diferite: F si Col E1. Ultima este mobilizabila, dar neconjugativa. Intr-un cuplu de conjugare, plasmida F poate suplini incapacitatea conjugativa a plasmidei Col E1, astfel incat aceasta va fi transferata. Procesul in care o plasmida neconjugativa este transferata intr-un proces mediat de o plasmida conjugativa, se numeste donatie.
O plasmida conjugativa poate sa medieze transferul unei plasmide nemobilizabile, sau chiar al genelor cromosomale. In cuplul de conjugare F+ x F- se transfera, cu o frecventa mica, gene cromosomale sau plasmide neconjugative. Fenomenul se numeste conductie si defineste procesul de mobilizare pasiva a oricarui replicon, inclusiv a cromosomului celulei F+, prin intermediul Tn l000. Conductia este rezultatul recombinarii genetice intre cei doi repliconi, mediat de Tn l000. Treapta intermediara a procesului de transpozitie, este o structura cointegrata, in care cele doua copii ale transpozonului au rolul unor punti intre cei doi repliconi. Transferul ADN cromosomal este initiat in stadiul de cointegrare a celor doi repliconi.
La nivel molecular, conjugarea si transferul ADN sunt procese complexe, reglate de gene si proteine multiple. Genele implicate in procesul conjugarii sunt organizate intr-un singur operon - operonul tra - organizat in circa 35 cadre de citire (ORF).
Dupa retractarea pilului si formarea canalului de conjugare, are loc mobilizarea plasmidei F. Mobilizarea incepe in momentul in care o proteina cu functie nucleazica - proteina Tra - codificata de plasmida, realizeaza incizia unei catene, la o secventa unica de baze, denumita originea de transfer (ori T) si ramane legata covalent de capatul 5'P al catenei incizate. Proteina Tra are si functie de helicaza si despiralizeaza plasmida F in timpul transferului conjugativ. Energia necesara pentru separarea catenelor este furnizata de hidroliza ATP.
Mecanismul transferului conjugativ este ipotetic. In acord cu unul dintre mecanismele de transfer, incizia monocatenara declanseaza replicarea plasmidei dupa modelul cercului simplu si ramura lineara ce rezulta din replicare este transferata. Proteina Tra, legata la capatul 5'P are, probabil, rolul de protectie a ADN linear monocatenar, de actiunea nucleolitica si este transferata concomitent cu ADN, in celula receptoare. Aceiasi proteina ar avea si rol de ligaza, reunind capetele catenei lineare in celula receptoare.
Una dintre proteinele codificate de operonul tra, care insoteste ADN in timpul transferului, este SSB. Ea tapeteaza ADN transferat, protejandu-l de actiunea endonucleazelor.
In cursul transferului, sinteza ADN se desfasoara atat in celula donoare cat si in celula receptoare. In celula donoare, sinteza conjugativa inlocuieste catena ce se transfera, iar in celula receptoare, sinteza conjugativa converteste ADN monocatenar transferat, la forma dublu catenara.
Un alt model al transferului considera ca proteina Tra ramane in celula donor si ADN trece in celula receptoare sub forma monocatenara circulara, care nu necesita protectie fata de actiunea endonucleazelor.
In cuplul de conjugare F+- F-, genele transferate codifica factori accesorii : rezistenta la antibiotice, UV, metale grele sau degradarea unor compusi xenobiotici.
Transferul ADN cromosomal in cuplul Hfr x F-
In celulele F+, cu o frecventa mica, plasmida F se integreaza in cromosomul bacterian, printr-un proces de recombinare. Situsul de recombinare a plasmidei F in cromosom are loc, predominant, in transpozonul IS3. Se cunosc peste 20 de situsuri majore ale cromosomului, la nivelul carora poate avea loc integrarea plasmidei F, dar afinitatea factorului F pentru fiecare situs este variabila. Afinitatea semnifica probabilitatea schimbului de secvente nucleotidice intre aceste elemente. Factorul F se poate integra in orientare directa sau inversata.
Celula bacteriana purtatoare a factorului F integrat devine Hfr, deoarece are o capacitate foarte ridicata de a initia formarea cuplurilor de conjugare. Factorul F linear, integrat in cromosomul bacterian, isi pierde capacitatea de replicare independenta si este replicat odata cu genele cromosomale. Odata cu diviziunea celulei, factorul F este transmis pe verticala, tuturor celulelor, de la generatie la alta. In stare integrata, factorul F nu mai este infectios - nu se transmite pe orizontala prin conjugare - decat in mod exceptional, cand conjugarea dureaza foarte mult.
O particularitate functionala distincta a bacteriilor Hfr, decurge din tendinta foarte marcata de rupere a cromosomului. De aici rezulta capacitatea foarte crescuta (de l00%) a cuplurilor de conjugare, de a transfera gene cromosomale. Genele transferate sunt diferite de la un cuplu de conjugare la altul, datorita numarului mare de situsuri cromosomale la care se poate face integrarea factorului F. Totdeauna sunt transferate genele in care s-a inserat factorul F.
Ruperea cromosomului se face, de regula, la nivelul structurilor plasmidei, foarte aproape de una dintre extremitatile ei, astfel incat cromosomul linear care se genereaza, poarta o extremitate, un mic fragment al plasmidei, iar restul se gaseste la extremitatea opusa. Rareori, incizia se face la limita dintre plasmida F si cromosom.
In functie de pozitia lor fata de situsul integrarii plasmidei F, fiecare gena cromosomala are sansa sa fie transferata in celula receptoare.
Mecanismul transferului de material genetic in cuplul Hfr x F- este foarte asemanator cu acela descris pentru transferul factorului F in cuplul de conjugare F+ x F-. Transferul este precedat de incizia monocatenara a cromosomului, la nivelul plasmidei integrate. Incizia este
catalizata de proteina Tra, cu activitate nucleazica. Punctul inciziei semnifica originea transferului (ori T). Transferul se initiaza la circa 3 minute dupa amestecul celulelor conjugante si progreseaza cu o rata constanta, cuprinsa intre 45 000 - l00 000 nucleotide/minut. Rata uniforma de transfer a ADN in conjugare, face ca transferul conjugativ sa fie instrumentul esential de lucru, pentru cartarea genetica a cromosomului.
Din celula donoare se transfera ADN monocatenar. Concomitent cu transferul are loc replicarea cromosomului celulei donoare, dupa modelul cercului simplu. Catena ramasa in celula donoare are rol de matrita pentru sinteza unei noi catene. Rezulta astfel, o copie dublu catenara a ADN transferat. Nu se cunoaste originea energiei de transfer, care deplaseaza catena ADN din celula donoare in cea receptoare.
In cuplul Hfr x F- se transfera un mic fragment al plasmidei F, gene cromosomale si in mod exceptional se transfera o copie completa a factorului F.
De cele mai multe ori, conjugarea se intrerupe datorita factorilor de mediu, la intervale variabile de timp. Din aceasta cauza, in conjugare se transfera, de obicei, numai gene cromosomale. Nu se transfera factorul F, deoarece este localizat la capatul opus al ori T, astfel ca celula receptoare ramane F-. Factorul F se transfera numai daca durata procesului de conjugare este de 90-l20 minute, mult mai mare decat durata unei generatii celulare. Faptul este explicabil prin aceea ca, procesul conjugarii stopeaza cresterea si diviziunea celulelor cuplului.
Fragmentul de material genetic transferat are dimensiuni diferite, in raport cu durata procesului de conjugare. Celula receptoare a cuplului, dupa conjugare, este un merozigot (un zigot partial si temporar diploid). Daca nu este degradat de enzimele de restrictie, fragmentul de ADN primit, se poate integra in cromosom, printr-un proces de recombinare genetica. Daca ADN exogen poarta gene noi, dupa recombinare se exprima in celula receptoare, care astfel dobandeste caractere noi.
In general, conjugarea se realizeaza intre celulele aceleiasi specii, dar are loc si interspecific si chiar intergeneric, dar cu o frecventa mai mica. De exemplu, E. coli formeaza cupluri de conjugare cu celule ale propriei specii, dar si cu Salmonella sp., Proteus mirabilis etc.
Conjugarea la bacteriile Gram pozitive
La cocii Gram pozitivi (Str. pneumoniae, E. faecalis, Streptomyces) procesul conjugarii are particularitati distincte si este mediat de plasmide si de transpozonii conjugativi.
Transpozonii conjugativi sunt EGT, in mod obisnuit integrate in genomul bacterian si au functie de transpozitie. Din punct de vedere functional, transpozonii conjugativi cumuleaza proprietati ale transpozonilor propriu-zisi si ale plasmidelor:
se aseamana cu transpozonii propriu-zisi, pentru ca se excizeaza si se reintegreaza in ADN;
se aseamana cu plasmidele pentru ca se excizeaza si formeaza un intermediar circular inchis covalent, fizic independent, ce se poate integra in genomul aceleiasi celule (transpozitie intracelulara) ori se transfera prin conjugare, la o celula receptor si se integreaza in genomul ei (transpozitie intercelulara).
Diferenta fata de Tn propriu-zisi consta in existenta unui intermediar dublu catenar inchis covalent. Se deosebesc de plasmide, deoarece intermediarul nu se replica.
La bacteriile Gram pozitive, strategia formarii perechilor sau a agregatelor de imperechere este diferita. Plasmidele si transpozonii conjugativi sunt prea mici pentru a codifica numeroasele proteine Tra ale aparatului de conjugare.
Conjugarea la Streptomyces
Este un proces distinct de schimbul genetic al bacteriilor, atat in ceea ce priveste mecanismul molecular, ci si din punct de vedere fenotipic. Fuziunea hifelor nu s-a evidentiat microscopic. Proteina Tra codificata de plasmida, singurul factor esential pentru transferul conjugativ, poate sa conditioneze si procesul de fuziune.
Plasmida nu codifica structurile anatomice care sa medieze formarea perechilor la varfurile hifelor celor 2 parteneri. Streptomyces este imobil si creste ca un miceliu cu ramificare multipla, pe mediul solidificat. Varfurile miceliului pot fuziona numai daca se sintetizeaza proteina de transfer (Tra), iar raspandirea plasmidei prin peretii transversali ai miceliului receptor necesita alte 3-5 gene.
Semnul distinctiv fenotipic al transferului plasmidei este aparitia zonelor pustulare care semnifica inhibitia cresterii. Dupa cultivarea pe mediu agarizat, pe suprafata placii apar zone vizibile macroscopic, de 1-3 mm diametru. Acestea sunt ariile in care miceliul receptor a primit o plasmida. {n interiorul structurii pustulare, diferentierea morfologica a miceliului receptor care a dobandit o plasmida, este temporar intarziata. Aceasta trasatura fenotipica face din Streptomyces, singurul microorganism la care schimbul genetic este vizibil cu ochiul liber.
Transferul conjugativ are loc in conditii restrictive: numai pe mediul solidificat, numai in faza de crestere timpurie, cand Streptomyces creste ca miceliu de substrat. Dupa inceperea diferentierii morfologice, transferul conjugativ este abolit.
Diseminarea plasmidei transferate in compartimentele miceliene ale receptorului este mediata de proteinele Spd (Spreading). Ele par sa se asambleze formand o structura de forma unui por in peretii transversali ai miceliului receptor si favorizeaza diseminarea plasmidei la partile mai vechi ale coloniei de Streptomyces.
Astfel, cea mai mare parte a miceliului este colonizata de plasmida. Este indusa transcrierea genei tra, deoarece compartimentele miceliene care abia au primit plasmida nu au un represor functional. Supraexpresia tranzitorie a proteinei Tra este cauza aparitiei zonelor inhibitoare ale cresterii si intarzierii diferentierii morfologice, ce apare sub forma structurilor pustulare specifice.
Conjugarea a fost descrisa si la alte actinobacterii: Mycobacterium, Corynebacterium, Rhodococcus.
Evenimentele moleculare ale procesului de transfer genic nu se cunosc.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 4994
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved