| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Structura membranei celulare
Membrana (7,5 nm) - este un ansamblu de structuri supramoleculare aflat la periferia celulei (sau a organitelor celulare) care separa mediul intern de cel extern (celular de interstitial) si care indeplineste diferite functii legate in special de procese metabolice fundamentale. Membranele vii au proprietatile de semipermeabilitate si selectivitate. Datorita acestor proprietati ele nu reprezinta simple bariere care separa 2 medii cu proprietati diferite, ci structuri active care asigura:
- transportul unor molecule, ioni, macromolecule, complexe supramoleculare, dintr-o parte in alta a ei;
- traducere si transfer de informatie adusa de diferiti stimuli (mecanici, electrici, electromagnetici, chimici, termici etc.) prin receptorii specifici pe care ii contine;
- interconversia diferitelor forme de energie, prin enzimele sau complexele enzimatice pe care le contine.
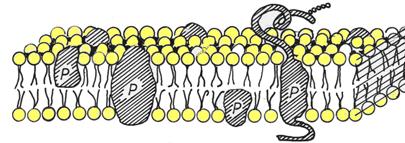 Actualmente
se accepta ca, in general, structura membranei celulare este cea
descrisa de modelul mozaicului fluid proteolipidic al lui Nicholson
si Singer (1972). Conform acesui model, membrana
este formata dintr-un bistrat lipidic, in care sunt inserate proteine
si glicoproteine (fig.)
Actualmente
se accepta ca, in general, structura membranei celulare este cea
descrisa de modelul mozaicului fluid proteolipidic al lui Nicholson
si Singer (1972). Conform acesui model, membrana
este formata dintr-un bistrat lipidic, in care sunt inserate proteine
si glicoproteine (fig.)
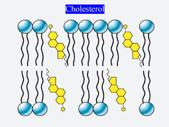
Bistratul lipidic este constituit in special din fosfolipide (fosfatidilcolina, fosfatidilserina, fosfatidilinositol etc.) dar sunt prezente si sfingolipide, glicolipide si colesterol.
Lipidele membranare sunt:
Fosfogliceride (fosfolipide)(compuse din acizi grasi, glicerol, fosfat, alcool)(au un cap polar si doua cozi hidrofobe) - reprezinta constituentul major al bistratului lipidic membranar Capetele polare pot fi : acid fosfatidic- PA, fosfatidilglicerol - PG, fosfatidiletanolamina - PE, fosfatidilcolina - PC, fosfatidilserina - PS, fosfatidilinositol - PI.
Sfingolipide (glicosfingolipide - GS-capul polar contine zaharuri, sfingomieline- SM- au capete polare fosforilcolina sau fosfoetanolamine)) (sfingozina - inlocuieste glicerolul si un lant hidrofob din cazul fosfogliceridelor)
Colesterol
Glicolipide
Insule de colesterol cu sfingilipide formeaza faze separate - rafturi lipdice- mai ordonate, contin proteine transmembranare si proteine periferice pe partea extracelulara. Se afla pe monostratul extracelular.
Moleculele lipidice sunt amfifile. Ele au un cap polar (extremitate polara) si o extremitate hidrofoba (doua lanturi de hidrocarburi - cozi hidrofobe). Moleculele de acest tip se organizeaza spontan, pe baza principiului de minimizare a energiei potentiale, astfel incat cozile lor hidrofobe sa evite contactul cu apa. Ele realizeaza:
- structuri micelare
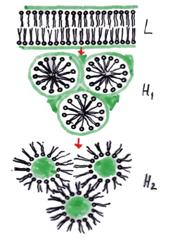 - bistraturi (vezicule membranare sau membrane
plane)(fig.)
- bistraturi (vezicule membranare sau membrane
plane)(fig.)
In acest fel, intre cozile hidrofobe iau nastere legaturi hidrofobe, iar capetele polare sunt expuse mediului apos. Bistratul lipidic este o structura dinamica, prezentand fluiditate: moleculele lipidice executa miscari de translatie si rotatie. De exemplu:
-translatie in stratul in care se afla (difuzie laterala)
- rotatie in jurul propriei axe
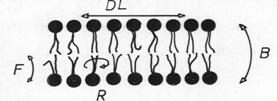 - basculare dintr-un monostrat in celalalt
- basculare dintr-un monostrat in celalalt
- flexie (indoire) (fig.)
Proteinele membranare, in functie de modul in care se insereaza in membrane, sunt:
a) - proteine intrinseci (integrale) - traverseaza membrana celulara o data (glicoforina) sau de mai multe ori (exemplu - proteinele transportoare, pompe ionice constituite din mai multe a -helixuri, enzime, receptori, canale ionice
b)- proteine extrinseci (periferice) - patrund in membrana pe o anumita distanta, pe una din cele doua fete, sau sunt atasate la suprafata membranei (receptorii membranari, proteine cu rol imunologic etc.). Ele pot participa la reactii enzimatice si semnalizare celulara, intra in componenta scheletului membranar de la suprafata citoplasmatica a celulei, asigura legatura cu citoscheletul. Membrana celulara se sprijina pe un citoschelet alcatuit din proteine fibrilare.
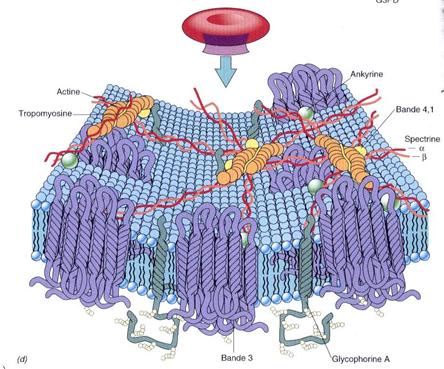
De
exemplu, in cazul hematiei (fig), doua proteine, Banda 3 (transportor de
anioni) si glicoforina, ancoreaza o retea bidimensionala de
proteine fibrilare, componenta principala fiind spectrina. Ankirina se
leaga atat la spectrina cat si la Banda 3. Filamente scurte de
actina cu proteinele asociate formeaza noduri care interconecteaza
reteaua de spectrina. Asigura elasticitatea membranei plasmatice
a hematiei.
Si proteinele prezinta miscari de difuzie laterala si rotatie, intr-un mod mai restrictiv (depinzand de interactiunile cu alte proteine). Unele dintre aceste miscari sunt orientate dar in acest caz intervin proteine motoare.
Membranele artificiale (model) - permit studiul unor procese membranare. Dintre acestea mai raspandite sunt lipozomii si bistratele lipidice (ex. BLM - membrane lipidice negre si SSM - membrane lipidice pe suport solid).
Clasificarea proteinelor integrale
Pompele ionice - enzime - utilizeaza diverse surse de energie - predominant ATP - pentru a deplasa ionii (mai ales cationii) si alti solviti prin membrana celulara. In general stabilesc gradienti de concentratie ai speciilor respective de-o parte si alta a membranei. Sunt implicate in transportul activ primar.
Transportorii sunt proteine (enzyme like) care asigura cai pasive de deplasare a diferitelor specii de solviti prin membrane conform gradientului lor electrochimic (de la concentratie mai mare spre concentratie mai mica). Unii dintre acestia, transportori uniport, asigura difuzia facilitata, tip de transport pasiv. O alta categorie etse implicata in transportul activ secundar, forma de transport in care translocarea unei specii conform gradientului electrochimic este insotita de translocarea altei specii contra gradientului sau electrochimic.
Canalele ionice sunt structuri proteice care au in interior un canal cu specificitate ionica, ce poate sa se deschida si sa se inchida tranzitoriu. Cand canalul este deschis, prin el va trece un flux de ioni cu orientare neta in sensul gradientului electrochimic. In acest fel se pot controla valorile potentialului electric transmembranar si pot fi generate semnale electrice in diferite tipuri de celule (nervoase, musculare, glandulare). Asigura transportul pasiv al ionilor.
Transportul membranar
1. Macrotransport (endocitoza, transcitoza, exocitoza):
- fagocitoza
- pinocitoza
- endocitoza prin intermediul receptorilor membranari
2. Microtransport
- pasiv
- activ
1. Macrotransport
Fagocitoza este un proces in care celula inglobeaza particule de substanta solida, invelite in pseudopode (prelungiri citoplasmatice) care fuzioneaza in spatele lor.
Pinocitoza reprezinta invelirea picaturilor de lichid
si macromoleculelor intr-un bistrat lipidic si formarea de vezicule
care fuzioneaza cu membrana celulara. In acest mod speciile
respective pot fi transportate dintr-o parte in cealalta a membranei.
Exemple: exocitoza in terminatiile nervoase si in celulele secretorii (acetilcolina)
- trecerea proteinelor plasmatice din sange in spatiul extravascular - transcitoza prin endoteliul capilar.
2. Microtransport
Transportul pasiv reprezinta deplasarea moleculelor si ionilor in sensul gradientului electrochimic sau de presiune - aparent fara consum de energie metabolica. Aparent, deoarece gradientul electrochimic respectiv este rezultatul unor procese anterioare realizate cu consum de energie.
Prin transport pasiv sistemul are tendinta de a ajunge la echilibru termodinamic. Daca in expresia diferentei de potential electrochimic notam:
c1 = cin c2 = cex V1 = Vin V2 = Vex obtinem:
DW = Win - Wex = RT ln cin/cex + zF (Vin - Vex)
DW > 0 - ionii au tendinta de a parasi celula;
DW < 0 - ionii au tendinta de a patrunde in celula, daca membrana este permeabila pentru acestia. Transportul unei specii ionice inceteaza la echilibru DW = 0, de unde rezulta:
E = Vin - Vex = (RT/zF) ln cex/cin ( ecuatia Nernst)
Modalitati de transport pasiv ;
Difuzia simpla
Difuzia facilitata
Difuzia prin canale si pori
Difuzia simpla se produce prin dizolvarea speciei moleculare transportate in membrana si depinde de raportul dintre solubilitatea substantei respective in bistratul lipidic si solubilitatea ei in apa, deci de coeficientul de partitie b. Cum s-a aratat, conform legii lui Fick, in cazul membranelor coeficientul de permeabilitate P este P = bD/d.
Difuzia facilitata - se face prin utilizarea unor molecule transportoare existente in membrana sau introduse artificial in aceasta. Asemenea molecule transportoare au o anumita specificitate, recunoscand specia moleculara sau ionica pe care o transporta. Exista transportori pentru glucoza, colina, pentru diferiti ioni (in acest caz transportorul se numeste ionofor). Transportorii pot distinge speciile levogire de cele dextrogire. Ei actioneaza in sensul gradientului electrochimic. Mecanismul de transport se bazeaza pe proprietatea transportorului de a se putea gasi in doua stari conformationale T1 si T2. (fig.)
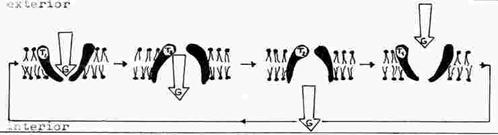
Molecula transportata (substratul S) se leaga pe una din fetele membranei. Se produce in urma legarii o modificare conformationala in starea T2 si situsul de legare este expus partii opuse (diagrama) cu scaderea afinitatii pentru specia respectiva si eliberarea acesteia. Prin eliberare se revine la conformatia initiala si ciclul se repeta. Procesul se desfasoara conform cineticii Michaelis-Menten pentru reactiile enzimatice. Un exemplu de ionofor este antibioticul valinomicina, molecula hidrofoba care poate incorpora ionii de K+, translocandu-i prin membrana (si Rb, mai slab). Valinomicina face ca ionii de K+ sa iasa din celula bacteriana, provocandu-i moartea. Nigericina (K+, H+, schimb neutru).
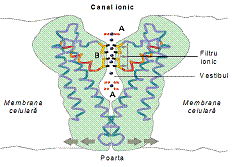
Difuzia
prin canale ionice si pori Canalele ionice sunt proteine specializate care
strabat bistratul si permit trecerea unor substante care nu sunt
liposolubile. Permit trecerea ionilor in ambele sensuri si sunt selective.
Porii sunt structuri neselective, conteaza doar diametrul particulei.
Specia transportata se leaga de proteina canal, formand un complex
enzima- substrat care evolueaza pe baza cineticii Michaelis-Menten.
Pentru ca ionul sa treaca dintr-o parte in cealalta este necesar
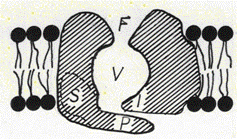
ca un canal sa fie deschis. Componentele unui canal ionic sunt (fig.): filtrul de selectivitate F, vestibulul V, senzorul S, poarta P, inactivator I. Canalele ionice prezinta o mare
specificitate, filtrul nu lasa sa treaca decat anumite specii.
Deschiderea si inchiderea canalului sunt rezultatul unei modificari conformationale care este comandata printr-un mecanism specific. Acesta poate fi ;
- electric - modificarea potentialului membranar (canalele de Na+, K+ in membrana axonala);
- chimic (acetilcolina, canalul de Na+, Ca++ controlat de GMPc in membrana celulelor fotoreceptoare);
- alte mecanisme (presiune -mecanic, ex. canalul de K+ de la polul apical al stereocililor celulelor ciliate din organul lui Corti).
Cand canalul este deschis are loc trecere pasiva a ionilor. Aceasta se poate face in ambele sensuri, dar este predominanta in sensul gradientului electrochimic. Canalele pot fi inhibate cu blocanti specifici - ex. toxine (tetrodotoxina - canalul de Na+). Asemenea experiente de inhibitie permit studierea proprietatilor canalelor.
Deosebiri intre cele doua tipuri de difuzie (facilitata, canale):
- transportorii - specificitate mai mare (disting intre L si D)(mii ioni/s);
- canalele - viteza mult mai mare (milioane - sute de milioane de ioni/s, sunt mai adecvate pentru modificari bruste;
- transportorii sunt in numar mult mai mare;
- transportorii pot participa la transportul activ secundar.
Transportul activ este o forma de transport care necesita energie metabolica - cuplare energetica imediata. Poate fi transport activ primar si transport activ secundar.
Transportul activ primar
Se realizeaza cu ajutorul pompelor ionice membranare, structuri proteice transportoare din clasa proteinelor integrale. O pompa ionica este caracterizata prin prezenta unui centru activ cu acces alternativ spre partea extracelulara si spre cea citoplasmatica. Accesul este modificat ca urmare a unei tranzitii conformationale.
Clasificarea pompelor ionice:
- Pompe ionice activate de lumina (proteine retinale cum ar fi bacteriorodopsina BR sau halorodopsina, proteine Fotoredox)
- ATP-aze ( familia F0F1, familia ATP-azelor de tip P, familia transportorilor ABC) care folosesc energia provenita din hidroliza ATP pentru a transloca diferite specii de pe o parte pe alta a membranei celulare.
Pompe ionice activate de lumina
Cel mai cunoscut si mai bine studiat reprezentant este bacteriorodopsina, membru al familiei proteinelor retinale, familie din care fac parte si rodopsinele vizuale. Bacteriorodopsina, prezenta in bacteria Halobacterium salinarum, este o pompa protonica activata de lumina care transporta protoni din citoplasma inspre exteriorul celulei. Gradientul protonic astfel creat activeaza o ATP-sintaza.
ATP-azele
Familia F0F1 are la randul sau doua subfamilii: F0F1 sau F-ATP-aze- care activeaza sinteza ori hidroliza ATP in functiile de conditiile mediului si V0V1 sau V-ATPaze - care controleaza acidifierea continutului diferitelor vezicules acestea transporta protoni. Se gasesc in sistemul vacuolar al eucariotelor.
F- ATP azele se gasesc in eubacterii, mitocondrii, cloroplaste si utilizeaza gradienti de protoni creati de alte proteine membranare (redox sau activate de lumina) pentru sinteza ATP (ex. ATP-sintaza). Au doua parti (fig.), F1 globulara, solubila in apa, catalizeaza hidroliza sau sinteza ATP (subunitatile beta) in functie de conditii. Subunitatile C sunt cele prin care trec protonii.
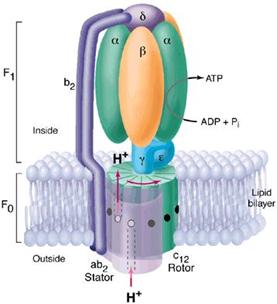
La trecerea protonilor se roteste partea globulara si prin rotire se stabileste o conformatie favorabila sintezei de ATP. Invers, cand exista ATP in exces, acesta este hidrolizat si se elibereaza energie care duce la rotatia subunitatilor C. Astfel este favorizat transportul de protoni. Mecanismul transportului de protoni in subunitatile C nu este cunoscut.
V-ATP azele sunt exclusiv pompe protonice, nu pot sintetiza ATP, doar il hidrolizeaza. Au structura asemanatoare cu cea a poteinelor F-type.
Familia ATP-azelor de tip P (ATP-aze E1E2) contine proteine care produc diversi gradienti transmebranari. Din aceasta familie fac parte pompe ionice bine cunoscute cum ar fi:
Na , K -ATP-aza care transporta 3Na spre exteriorul celulei si 2K spre interior. Se gaseste in membrana plasmatica celulara, genereaza gradienti de Na si K
H , K -ATP-aza, transporta 1 H spre exteriorul celulei contra a 1 K spre interiorul celulei, se gaseste in membranele plasmatice ale celulelor stomacului si rinichilor, este implicata in secretia gastrica si duodenala.
Ca -ATP-aza SERCA, transporta 2 Ca spre SR ori ER si 2 H spre citosol, se gaseste in reticulul sarcoplasmic si in cel endoplasmic, scade concentratia de Ca intracelular.
Ca -ATP-aza PMCA, transporta 1 Ca spre exterior contra a 1 H , se gaseste in membrana plasmatica, scade concentratia de Ca intracelular.
H -ATP-aza, transporta in afara celulei 1 H , se gaseste in membrana plasmatica a drojdiilor, plantelor, protozoarelor, genereaza gradient protonic.
P-ATP-azele pot fi implicate in unele boli, de ex.: mutatii ale Ca ATP-azei induc rigiditate si crampe musculare, ale Cu ATP-azelor pot fie sa provoace sindromul Menks - pacientii au deficit de Cu , fie boala Wilson, incapacitatea de a elimina Cu din ficat cu efect toxic. In caz de ulcer se foloseste Omeprazolul care inhiba H ,K ATP-aza gastrica, scazand aciditatea gastrica.
Transportorii ABC formeaza familia cea mai larga si diversificata. Se gasesc in toate organismele. Substratele acestora pot fi ioni anorganici, zaharuri, aminoacizi, polizaharide complexe, peptide si chiar proteine. Unii dintre acestia pot sa fie in acelasi timp canale (CFTR). Dintre acestia mentionam doua tipuri:
CFTR (cystic fibrosis transmembrane conductance regulator) care permite ionilor de Cl sa iasa din celula pe baza gradientului lor electrochimic. Deschiderea canalului este comandata de hidroliza ATP. NaCl la polul apical al celulelor pulmonare creeaza o presiune osmotica datorita careia apa din celule este extrasa prin canalele pentru apa - aquaporinele. Speciile ionice sunt din nou transportate in celule prin pompa de Na, K, respectiv transportorul Na, K, 2Cl si circuitul se reia. In acest fel se pastreaza un echilibru hidric ce permite curatarea de bacterii si secretii a lumenului bronhiolelor. In lipsa Cl, prin mutatii la nivelul CFTR (nu se exprima o fenilalanina) plamanul este uscat si se acumuleaza secretii, mucus, prin activitatea bacteriilor. Apar infectii si nu se mai pot face corect schimburile de gaze. Boala se numeste fibroza cistica sau mucoviscidoza si se termina prin moarte.
MDR1 si MDR2 (multiple drug resistance proteins). Pot transporta o multitudine de molecule hidrofobe, molecule care pot intra in celula prin bistratul lipidic si care pot fi toxice pentru aceasta. MDR scot afara aceste molecule. Dar asemenea molecule pot fi si medicamentele utilizate in chimioterapie. Celulele canceroase pot sa supraexprime asemenea transportori reducand eficienta chimioterapiei.
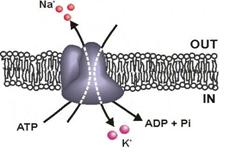 Cea mai cunoscuta pompa ionica este Na+,K+-
ATP-aza. Este alcatuita din
patru subunitati proteice (2 a si 2 b);
subunitatea a are activitate ATP-azica. Hidroliza ATP furnizeaza energia
care permite translocarea a 3 ioni de Na in exteriorul celulei si a doi
ioni de K in interior.
Cea mai cunoscuta pompa ionica este Na+,K+-
ATP-aza. Este alcatuita din
patru subunitati proteice (2 a si 2 b);
subunitatea a are activitate ATP-azica. Hidroliza ATP furnizeaza energia
care permite translocarea a 3 ioni de Na in exteriorul celulei si a doi
ioni de K in interior.
Este o pompa electrogenica intrucat activitatea ei are ca rezultat un transfer net de o sarcina pozitiva per ciclu in exteriorul celulei. Conform schemei Albers-Post, in esenta mecanismul de transport este: (schema).
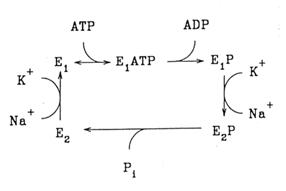
Enzima in conformatia E1 ia Na+ pe partea citoplasmatica si leaga ATP (in prezenta Mg++). ATP este hidrolizat, complexul fosforilat sufera o tranzitie conformationala E1 - E2 in urma careia scade afinitatea pentru Na+, creste pentru K+, acesta se ataseaza, are loc o defosforilare, se pierde afinitatea pentru K+, enzima trece din nou in conformatia E1. In repaus ATP-aza de Na, K este responsabila de cca. 30% din consumul energetic celular, iar in celula nervoasa consumul ajunge la cca. 70%.
Prin acest tip de transport are loc patrunderea speciilor transportate intr-un compartiment (extracelular sau intracelular) impotriva gradientului lor electrochimic, prin asocierea cu molecule care se deplaseaza normal conform propriului lor gradient de concentratie. Gradientul este insa mentinut prin transport activ primar. Transportorul intalnit in difuzia facilitata poate lega cele doua molecule fie in aceeasi stare conformationala (pe aceeasi parte) - simport sau co-transport, fie pe cele doua parti, in stari conformationale diferite - antiport (contra-transport).
In simport - enzima leaga pe aceeasi parte, in T1, ambele specii si sufera tranzitia T1 - T2 numai dupa legarea ambelor (vezi figura de la difuzia facilitata)..
Exemplu (fig.): glucoza intra in celulele mucoasei intestinale prin asociere cu Na+ care intra pasiv conform gradientului sau electrochimic. Transportul este electrogenic. Ionii de Na+ sunt apoi eliminati activ prin transport primar, prin hidroliza ATP. Glucoza ramane in celula si apoi este transportata in sange printr-un mecanism uniport, conform gradientului ei de concentratie.
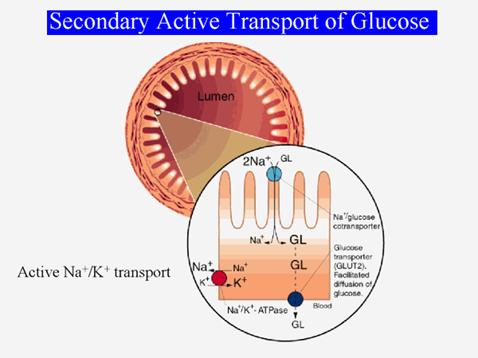
In antiport - proteina transportoare leaga in starea T2 un alt ion decat cel care sufera difuzia facilitata, care va fi evacuat in compartimentul in care potentialul lui electrochimic este mai mare. Exemplu (fig.): muschiul cardiac - antiport 3 Na+/ 1 Ca++ - electrogenic (sarcina neta +1). Asigura concentratia scazuta a Ca++ in interior pe seama pomparii active a Na+.
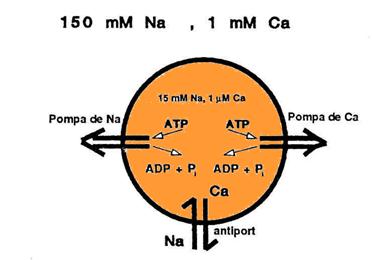
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 5471
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2026 . All rights reserved