| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Elementele genetice transpozabile ale bacteriilor si transpozitia
Elementele genetice transpozabile* (EGT) sunt secvente specifice de ADN, mobile, care se pot desprinde (exciza) din insertiile lor si se reintegreaza in alte situsuri ale aceluiasi replicon sau in repliconi diferiti.
Elementele genetice transpozabile s-au identificat initial la porumb (McClintock, l952), sub denumirea de elemente de control. Ele controleaza activitatea (trecerea in pozitia de start sau de stop), a genelor pigmentare a semintelor de porumb. Descoperirea a venit prea devreme, ideile au fost considerate erezii in raport cu dogma dominanta si nu au fost luate in consideratie. Interesul pentru EGT a fost reinoit odata cu evidentierea lor in structura genomului bacterian. EGT se gasesc la organisme foarte diferite ca pozitie sistematica (bacterii, levuri, porumb, Drosophyla) si probabil au o raspandire universala in lumea vie, iar la bacterii par a fi ubicvitare. S-au descris la E. coli, Citrobacter, Salmonella, Shigella, Pseudomonas, Klebsiella, Rhizobium, Staphylococcus, Haemophilus, Bacillus.
Termenul de transpozitie (sau translocatie) semnifica schimbul unui secvente de ADN intre repliconi diferiti sau intre regiuni neomologe ale aceleiasi molecule de ADN. Fenomenul transpozitiei se materializeaza prin aparitia unei secvente de ADN cu o lungime definita, intr-o localizare genomica noua, in care anterior nu a fost detectat. Caracterul mobil al EGT este reflectat in diferitele denumiri propuse: gene calatoare, elemente mobile, gene care sar, vehicule genetice autopropulsate etc. S-au denumit gene egoiste, deoarece functia unora dintre ele este numai aceea de a se replica, fara sa codifice o proteina detectabila.
Caracteristica generala a EGT este ca toate au capacitatea de a transpoza, adica de a se insera la diferite situsuri, fara omologie genetica, pe acelasi sau pe un alt replicon.
Se cunosc 4 clase de EGT:
a) secventele de insertie (SI) si transpozonii simpli
b) transpozonii complecsi
c) bacteriofagii transpozanti (fagii l si Mu)
d) transpozonii conjugativi.
Secventele de insertie (SI) si transpozonii (Tn) nu au existenta autonoma, ci se gasesc numai in stare integrata in structura unui replicon, ca segmente lineare de ADN, cu extremitati bine definite, in timp ce fagii prezinta o faza libera, extracelulara.
Nomenclatura. Este o adevarata explozie in ceea ce priveste nr diferitelor EGT izolate si caracterizate prin secventa Nt. Pana in '89 erau analizate numai 50 SI, iar in '98, nr lor a depasit 500. S-au gasit SI la peste 240 specii de insecte, la fungi, mamifere, pesti si plante.
SI sunt definite imprecis ca segmente criptice mici de ADN(<2,5 kbp), cu organizare genetica simpla si capabile sa se insere la situsuri multiple intr-o molecula tinta.
Au fost excluse retravir, retrotranspozonii si retropozonii, Tn cojugativi care folosesc un mecanism de tipul fagului pt translocatie si elemente ca fagul Mu, Tn7 si Tn 554 care sunt mari si relativ complecsi.
S-au folosit cel putin 2 sist de nomenclatura a SI:
- cel initiat de Lederberg in '78 atribuie un singur numar unei SI (de ex. IS 1). Sistemul nu include suficienta informatie si este insuficient, odata cu cresterea nr. de structuri genetice cunoscute.
- al II-lea sistem aduce informatie suplim, privind originea elementului pt ca include initialele speciei bacteriene de origine (de ex., IS Rm 1) pt Rhiz. meliloti.
Secventele de insertie
Secventele de insertie (SI) sunt cele mai simple elemente transpozabile descrise la bacterii. Ele contin circa l000 bp, desi unele sunt mai mici (IS1 contine 769 bp) sau mai mari (l53l bp pentru IS5o). Secventele de insertie nu contin nici o gena cunoscuta si nu au nici o alta functie in afara de cea de replicare (fig. 125).
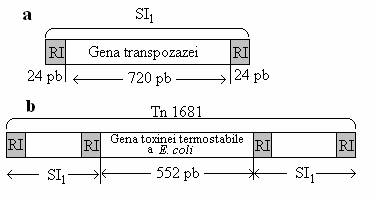
Fig. 125. a. Reprezentarea schematica a secventei de insertie SI1, care contine gena enzimei transpozaza, determinanta pentru mobilitatea ei, flancata la cele doua extremitati de secventele repetate invers. b. Transpozomul Tn1681, cu gena codificatoare a toxinei termostabile de E. coli, incadrata de SI1.
SI sunt mici si compacte, alcatuite dintr-o regiune centrala, codificatoare pentru functia de transpozitie, mediata de transpozaza (enzima care catalizeaza excizia acestor secvente din situsul de insertie). Regiunea centrala este flancata la ambele extremitati, de secvente terminale mai scurte (de 10-40 bp), repetate in ordine directa, (secvente repetate in aceiasi ordine la extremitatile unui segment de ADN, pe aceiasi catena) sau in ordine inversata (SRI* - secventa nucleotidelor in directia 5'---- 3' pe una din catene este aceiasi pe catena complementara, in aceiasi directie 5' --- 3', la capatul opus).
3' G T C G C A G A G ......... .. G T C G C A G A G 5'
5' C A G C G T C T C .......... C A G C G T C T C 3'
Secvente terminale Secvente nucleotidice Secvente terminale
directe interne directe
3' C C C A G A C ............ G T C T G G G 5'
5' G G G T C T G ............. C A G A C C C 3'
Secvente terminale Secvente nucleotidice Secvente terminale
inversate interne inversate
SRI pot fi divizate in 2 domenii functionale:
- unul e asezat in SRI si e implicat in legarea transpozazei
- celalalt include 2-3 bp terminale, e implicat in clivare si in reactiile de transfer, care duc la transpozitia elementului.
Secventele de insertie nu se replica autonom. O secventa de insertie se desprinde din insertia sa si se deplaseaza la un alt situs pe cromosom, in oricare din cele doua orientari posibile. Sunt elemente genetice criptice, adica prezenta lor nu este detectabila fenotipic, ci este dedusa indirect, prin mutatiile pe care le genereaza. Integrarea unei SI intr-o gena, suprima functia genei respective si rezulta o mutatie. Daca gena face parte dintr-un operon, integrarea SI in structura genetica a unui operon produce efecte polare severe.
Transpozonii
Transpozonii sunt segmente de ADN mobile
(cu potential de transpozitie), care includ in alcatuirea lor, o
serie de gene structurale, delimitate
la extremitati de SI, totdeauna repetate in ordine inversata.
Marimea lor variaza intre 2l00 - 9300 bp. Din
punctul de vedere al structurii genetice, Tn simpli sunt asemanatori
cu SI, dar se deosebesc de acestea prin faptul ca in secventa
centrala, poarta gene a caror expresie fenotipica este
decelabila si confera proprietati noi celulei
purtatoare (de exemplu, gene de rezistenta la Km, Cm, Tc etc.). Genele structurale sunt
flancate de secvente repetate lungi (l-2 kb), cel mai adesea in ordine
inversata. Secventele inversate confera o stabilitate
superioara structurii
Transpozonii complecsi - grupati in familia Tn1 au pana la 20 kb, dar au fost descrisi altii mult mai mari (80 kb). Se disting de transpozonii simpli prin aceea ca atat secventele de transpozitie cat si secventele specifice sunt flancate de SRI scurte (40 pb).
Interesul pentru studiul transpozonilor s-a amplificat odata cu descoperirea faptului ca genele de rezistenta la antibiotice pot sa treaca de la un replicon la altul. De exemplu, gena pentru β-lactamaza este foarte mobila, trecand cu usurinta de pe o plasmida pe alta, ceea ce explica raspandirea larga a rezistentei bacteriene la antibioticele β-lactamice.
La multe bacterii, patogenitatea si virulenta sunt conditionate de gene care fac parte din structura EGT: Bacillus, Bordetella, Brucella, Campylobacter, Clostridium, Escherichia, Haemophilus, Neisseria, Vibrio, Yersinia, dar ]i la patogeni ai plantelor: Agrobacterium, Erwinia, Pseudomonas, la simbiontul Rhizobium.
Alt domeniu de importanta practica este bioremedierea: Tn poarta gene care sunt parti ale cailor degradative ori catabolice (Tn ce poarta gene pentru utilizarea citratului).
Bacteriofagi transpozanti
Fenomenul transpozitiei este cunoscut pentru fagul Mu-1 (Mutator), care infecteaza celulele de E. coli K12. Capul este icozaedric, iar coada cu simetrie helicala, este prevazuta cu cele 6 fibre ale placii bazale. Molecula de ADN are 39 kbp, dar numai 37,2 kbp formeaza genomul, deoarece ambele capete ale moleculei de ADN, contin secvente ale cromosomului bacterian
Dupa infectie, Mu-l poate urma o cale lizogena sau calea litica.
Evenimentul comun pentru evolutia litica sau lizogena este insertia sa ca profag in cromosomul celulei. Insertia se face aleatoriu, producand mutatii cu o frecventa foarte mare. Genomul fagic integrat (profagul) se deplaseaza continuu prin transpozitie, catalizata de o transpozaza specifica, in diferite situsuri cromosomale, reintegrandu-se in oricare din cele doua orientari posibile. Genomul fagic se replica prin transpozitii succesive. Situsul transpozitiei nu are specificitate, genomul fagic integrandu-se la diferite situsuri ale cromosomului bacterian.
Fagul Mu-1 produce o varietate de rearanjari ale cromosomului bacterian. Mutatiile sunt rezultatul efectului polar intens, ce consta in inactivarea tuturor genelor operonului situate in aval fata de punctul de insertie, in raport cu secventa promotor.
Particularitatile ciclului de replicare a fagului Mu-1 au fost clarificate numai dupa descoperirea EGT.
Pentru evolutia litica, genomul ADN Mu-1 este excizat sub actiunea unei nucleaze fagice. Excizia este incorecta: cu l0-l50 de baze la stanga fata de capatul stang al genomului, iar la capatul din dreapta, 500-3000 de nucleotide isi au originea in cromosomul gazdei. Se sintetizeaza proteine virale si se asambleaza virioni progeni. Secventele de nucleotide care flancheaza genomul fagic se pierd totdeauna prin clivare, inainte de integrarea sa in cromosomul bacterian.
Fagul Mu-1 este considerat a fi un transpozon de tip special, de dimensiuni mari, ce codifica nu numai enzima de transpozitie (transpozaza), ci si toate proteinele necesare asamblarii si morfogenezei capsidei.
Transpozitia
Transpozitia semnifica excizia EGT din situsul sau de integrare intr-un replicon si insertia intr-un alt situs al aceluiasi sau al altui replicon. Integrarea EGT se realizeaza prin procese de recombinare nelegitima, datorita absentei omologiei genetice cel putin pe o lungime considerata ca minima, intre cele doua secvente ale ADN recombinant. Recombinarea are loc independent de produsul genei rec A, mediatorul recombinarii genetice intre secvente omologe de baze.
Transpozitia este un fenomen rar, datorita concentratiei foarte mici de transpozaza in celula. De exemplu, Tn1o codifica sinteza unei singure molecule/celula/generatie. Transpozitia sa are loc, in medie, o data
la l07 generatii celulare.
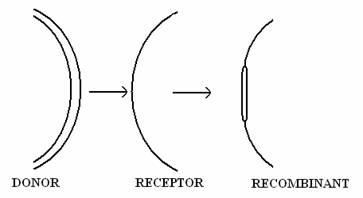
Fig. 126. Reprezentarea schematica a procesului de transpozitie. Secventa din molecula donor este reprezentata prin linia dubla, iar molecula receptor, prin linie simpla.
Mecanismul molecular al transpozitiei este ipotetic. Transpozaza se sintetizeaza imediat dupa replicarea ADN. Transpozitia este asociata in timp, cu momentul sintezei transpozazei. Unele EGT (de exemplu, genomul fagului lambda) se integreaza numai la situsuri cromosomale bine determinate, asa numitele "zone calde" ale transpozitiei. Altele (de exemplu, IS1) se insera aproape la orice situs cromosmal, desi prefera anumite secvente. Ca si enzimele de restrictie, transpozonii recunosc secvente palindromice de 4-6 perechi de baze.
Transpozonii sunt excizati cu precizie la extremitatile insertiei lor in replicon, deoarece transpozaza recunoaste ambele extremitati ale situsului de insertie. Moleculele de transpozaza leaga ambele extremitati ale transpozonului, precum si secventa de baze a repliconului receptor. Ambele catene ale moleculei acceptoare sunt clivate decalat, dupa mecanismul de actiune al enzimelor de restrictie. Simultan se produce clivajul monocatenar al extremitatilor transpozonului, pe catenele opuse, astfel incat transpozonul are o catena libera la fiecare capat. Dupa legarea monocatenara a fiecarei extremitati a EGT la situsul receptor, fenomenul transpozitiei poate urma una din urmatoarele doua cai:
a. Modelul transpozitiei conservative sau simple presupune ca transpozaza produce al II-lea set de sectiuni la cele doua capete ale transpozonului. Rezultatul este transferul transpozonului intact la noul situs. Continuitatea catenelor repliconului receptor este restabilita de ADN-ligaza. Discontinuitatea catenelor repliconului nu mai este reparata si molecula este degradata de nucleaze. Transpozitia simpla implica numai ruperea si reunirea catenelor repliconului receptor si deplasarea EGT la un nou situs;
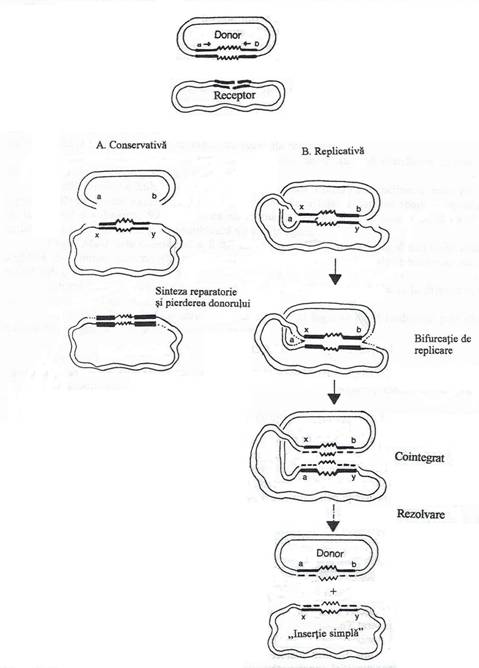
Fig. 127. Modelul molecular al transpozitiei. A. Transpozitia conservativa (simpla). B. Transpozitia replicativa, cu formarea unei structuri intermediare cointegrate. Unii transpozoni se separa din structura cointegrata, sub actuinea ''rezolvazei''', enzima care sectioneaza si leaga capetele celor doi repliconi. Alteori, repliconii fuzioneaza intr-o structura unica. Liniile groase intre a si b delimiteaza transpozonul, in repliconul donor ; linia in zig-zag semnifica secventele terminare repetate ; x, y = situsurile de clivare in cei doi repliconi. Alte amanunte, in text (dupa Davis, 1990).
b. Modelul transpozitiei replicative presupune ca in momentul transpozitiei, EGT este replicat: o copie a sa este transpozata la situsul receptor, iar cea de a II-a ramane la situsul original al repliconului donor. Cea de a II-a serie de sectiuni a catenelor EGT, nu mai are loc, astfel incat cei doi repliconi fuzioneaza si formeaza o structura cointegrata.
In procesul fuziunii, EGT este copiata integral si rezulta doua copii. Ulterior, cei doi repliconi se separa, dar fiecare poarta cate o copie a EGT, sau raman fuzionati intr-o structura unica. Prin procesul transpozitiei replicative a unui EGT dintr-o plasmida intr-o alta plasmida, urmat de fuziunea celor doi repliconi, se formeaza o plasmida mai mare, stabila, cu cate o copie a EGT la ambele puncte de jonctiune.
Modelul transpozitiei replicative pare a fi cel mai probabil. Transpozitia nu consta intr-un schimb de material genetic, ci in dobandirea unui EGT.
Desi transpozitia EGT este rezultatul unor procese de recombinare nelegitima, totusi fenomenul are loc intre secvente cu un grad variabil de omologie a bazelor, intre slaba si moderata. Face exceptie fagul λ, care se insera cu specificitate inalta, la situsuri omologe, existente atat in cromosomul de E. coli (att B), cat si in cel fagic (attP).
La bacterii, cele mai multe EGT se deplaseaza (transpozeaza) foarte rar (odata la l05 - l07 generatii celulare) si de aceea se diferentiaza greu de componenta imobila a cromosomului.
La levuri, la plante, la protistele eucariote, la Drosophyla si la procariote s-au gasit EGT, a caror transpozitie se face dupa un mecanism similar ciclului de multiplicare a retravirusurilor. Prima treapta a transpozitiei este transcrierea EGT intr-o copie de ARN, de peste 5000 de baze. Aceasta copie codifica sinteza enzimei reverstranscriptaza, care catalizeaza sinteza unei copii de ADN dublu catenara, circulara, pe matrita de ARN. ADN circular se integreaza intr-un situs cromosomal, cu specificitate de secventa a bazelor. Integrarea ADN este catalizata de o transpozaza, codificata de copia de ARN. De aici deriva denumirea de retrotranspozoni sau retropozoni pentru aceste EGT. Ele au, in special, importanta teoretica, deoarece sugereaza posibila origine si evolutie a retravirusurilor. Singura deosebire intre retravirusuri si retropozoni pare sa conste in absenta invelisului viral.
Consecintele transpozitiei
Procesele de transpozitie produc o multitudine de rearanjari genetice. Nu numai ca deplaseaza diferite gene de la un situs la altul, dar transpozitia produce diferite macroleziuni: mutatii, deletii, inversii etc. Situsurile insertiei EGT sunt "zone calde" pentru astfel de modificari genetice. Insertia elementelor transpozabile perturba invariabil integritatea genei in interiorul careia s-a facut transpozitia si efectul este mutatia. Daca mutatia se produce intr-o gena proximala (in raport cu secventa operatoare) a operonului, ea exercita efecte polare intense asupra genelor situate distal fata de gena mutanta. Noul tip de mutatie s-a numit mutatie polara. Concluzia a fost ca mutatiile cu efect polar intens, rezulta prin insertia unei secvente de ADN, la situsul mutatiei.
Insertia unui Tn intr-un situs nou, prin efectele sale polare, produce frecvent o mutatie. Efectele polare se produc pentru ca EGT sunt, in general, entitati genetice de sine statatoare si adeseori semnifica terminarea transcrierii. De aceea, transcrierea operonului rareori depaseste situsul de insertie a EGT, in absenta unui promotor la capatul terminal, care sa reinitieze transcrierea dincolo de secventa EGT. Restul genelor operonului, situate in aval de situsul de insertie, vor ramane netranscrise (Bennett, l998).
Mutatiile prin deletie survin ca urmare a exciziei incorecte a transpozonului, impreuna cu secvente cromosomale.
Cea mai cunoscuta mutatie datorata unei inversii a secventei cromosomale este cea care determina fenomenul variatiei de faza, descris la Salmonella. Flagelii contin proteina H1 sau H2, cu specificitate antigenica distincta, niciodata ambele. Schimbarile compozitiei chimice, de la H1 la H2 sau invers, se produc cu o frecventa de circa l la l000 diviziuni celulare, cu mult peste rata mutatiei spontane. Fenomenul se datoreaza inversiilor cromosomale, al caror rezultat este exprimarea alternanta a genei h, in cele doua orientari (h1 sau h2). Cand segmentul de ADN codificator are orientare directa, se sintetizeaza proteina H1, iar sinteza proteinei H2 este
represata. Dupa inversiunea segmentului de ADN codificator, se sintetizeaza proteina H2, iar sinteza proteinei Hl este represata.
Transpozonii au rolul unor "comutatori" pentru genele adiacente situsului de insertie: in raport cu situsul integrarii, intr-o directie se initiaza transcrierea, iar in directia opusa, ori o inhiba, ori produc terminarea ei prematura.
Transpozonii pot avea consecinte functionale favorabile pentru celula, prin introducerea genelor noi.
Un alt efect al transpozitiei este fuziunea repliconilor. Un transpozon inserat intr-o plasmida, va usura integrarea ei ulterioara in cromosomul celulei, flancata fiind de doua copii ale transpozonului. Un exemplu in acest sens, este generarea liniilor Hfr (High frequency of recombination) la E. coli, dupa integrarea factorului F in cromosom. Fuziunea repliconilor poate sa fie rezultatul transpozitiei sau al unui crossing-over intre EGT cu secvente omologe, existente in ambii repliconi.
Datorita existentei elementelor genetice transpozabile, genomul bacterian a dobandit potentialul unei remarcabile plasticitati genetice.
Descoperirea plasmidelor a condus la ideia "genomului bacterian colectiv", suprapus celui individual, reprezentat de cromosom, decurgand din caracterul transmisibil al acestora cu o frecventa mare, de la o celula la alta.
Anterior descoperirii EGT, se considera ca mutatiile si recombinarile genetice omologe ("legitime") erau suficiente pentru a genera diversitatea lumii bacteriene in mediile naturale. Descoperirea lor a evidentiat faptul ca prin mobilitatea foarte accentuata, EGT sunt promotorii recombinarilor "nelegitime". Acestea se produc cu o frecventa mult mai mare decat recombinarile pe baza omologiei genetice, sporind astfel diversitatea mecanismelor de control genetic al proceselor fiziologice si permitand o mai eficienta adaptare a celulei bacteriene la conditiile variabile ale mediului de viata.
Integronii sunt secvente de ADN ce contin determinanti genetici ai unui sistem de recombinare cu specificitate de situs. Ei recunosc, inglobeaza si astfel mediaza deplasarea unor scurte secvente mobile de ADN, denumite casete genice (Recchia si Hall, l995). Cele mai multe casete genice cuprind o singura gena de rezistenta la antibiotice. Un integron contine o secventa ce codifica integraza. Integraza este o enzima ce catalizeaza recombinarea cu specificitate de situs. Integronul contine o secventa specifica de ADN, la care integraza poate functiona pentru integrarea casetelor genice si un promotor ce poate exprima caseta genica nou integrata. Intr-un integron pot fi grupate casete genice multiple.
Frecvent, integronii fac parte din structura unor transpozoni, dar au o distributie mai larga.
O caseta genica poate exista in doua forme:
ca o secventa lineara intr-un integron
ca o molecula circulara de ADN, dublu catenara, ce poarta l-2 gene de rezistenta si un situs de recombinare (att) de 59 de nucleotide, cu rolul de a integra caseta genica la situsul specific al unui replicon.
Secventele 5' si 3' care flancheaza caseta genica lineara, sunt bine conservate. Regiunea 5' contine gena int, codificatoare a integrazei, o enzima de recombinare la situs specific, iar la capatul 3' se gaseste gena de rezistenta la un antibiotic (fig. 128).
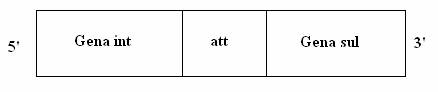
Fig. 128. Structura genica a unui integron. Secventa att mediaza recombinarea la situs specific. Gena sul codifica rezistenta la sulfonamide (dupa Reccchia si colab., 1995).
Integrarea unei casete genice intr-un replicon este reversibila. Caseta excizata se integreaza in alt replicon. Prin intermediul casetelor genice, complementul genelor de rezistenta poate fi mereu rearanjat si deplasat de pe un replicon pe altul.
Integronii sunt sisteme de diseminare a genelor bacteriene, distincte de plasmide si de EGT.
Integronii se gasesc aproape exclusiv la bacteriile Gram negative si mediaza rezistenta multipla la antibiotice (MDR): aminoglicozide, peniciline, cefalosporine, trimetoprim, tetraciclina, eritromicina, cloramfenicol.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 3764
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved