| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Fungii
Fungii sunt organisme eucariote, heterotrofe, care au evoluat dintr-un stramos comun cu al metazoarelor. Sunt mai inruditi cu metazoarele decat cu plantele. Asemenea majoritatii bacteriilor si organismelor animale, fungii folosesc substantele organice, ca sursa de energie. Nu contin pigmenti fotosintetizanti, ci pentru producerea energiei sunt dependente de sisteme enzimatice specifice.
Fungii pot fi organisme saprobionte, adica cresc pe materia organica in descompunere, de origine vegetala si animala. Altii sunt paraziti si preiau nutrientii din tesuturile organismelor vii, plante si animale, prin intermediul unor hife specializate denumite haustori. In ambele cazuri, enzimele hidrolitice sunt eliberate din celula fungica in mediul inconjurator, unde moleculele nutritive sunt simplificate prin digestie enzimatica, iar nutrientii trec in celula fungica, sub forma unei solutii apoase.
Organizare celulara si fiziologie
Celula fungica este
asemanatoare, din punct de vedere structural, cu cea vegetala
sau animala. Nucleul celulei fungice este foarte mic. Invelisul
nuclear dublu-membranar, adeseori, prezinta
In diviziunea meiotica, atat membrana nucleara, cat si nucleolul se dezorganizeaza.
Vacuolele, totdeauna prezente in celula fungica, niciodata nu ating dimensiunea celor din celula vegetala.
Celula fungica depoziteaza glicogen sau lipide.
Peretele celular consta din microfibrile de chitina sau de celuloza. Majoritatea fungilor au perete chitinos. Uneori, chitina si celuloza coexista in compozitia aceluiasi perete.
Intre plasmalema si peretele celular, apar structuri speciale denumite lomasomi, alcatuiti din agregate de tubuli sau din vezicule. Functia lor nu este cunoscuta.
Compozitia chimica a peretelui celular al levurilor este heterogena. El contine cantitati aproximativ egale de glucan (un polizaharid cu grad inalt de ramificare) si manan (un polizaharid solubil). Alti compusi chimici ai peretelui fungic sunt lipidele, proteinele si glucozamina (un glucid aminat).
Studiul suprafetei fungilor este important, deoarece la acest nivel se gasesc structurile moleculare care realizeaza interactiunile celulare in cursul reproducerii sexuate, in procesul flocularii levurilor de fermentatie, in inglobarea si excretia moleculelor. Studiile ultrastructurale au evidentiat, pe suprafata diferitelor levuri, prezenta fimbriilor, carora li se atribuie un rol important in etapa initiala a procesului de reproducere sexuata.
Unitatea structurala a aparatului vegetativ al fungilor este, de regula, hifa. Pe masura ce creste, hifa se ramifica intr-o retea densa de hife, care constituie miceliul vizibil ca o pasla fina. Fungii macroscopici sunt alcatuiti din hife impachetate intr-o structura denumita corp de fructificare. Miceliul este analog coloniei bacteriene.
Hifele sunt alcatuite din celule. Celulele fungice pot avea dimensiuni diferite: uneori o singura celula formeaza intregul aparat vegetativ al fungilor. In aceasta categorie intra levurile monocelulare.
Celulele fungice pot fi alungite
si filiforme, mononucleate sau binucleate. La Zygomycetes, hifele sunt coenocitice
(neseptate), adica dupa diviziunea nucleului, peretii
despartitori nu se formeaza si continutul hifei se
poate deplasa liber pe lungimea filamentului. La alti fungi, hifele
miceliene sunt divizate prin pereti transversali (septe). Adeseori,
septurile transversale sunt perforate de unul sau mai multi
Miceliul prezinta diferentieri functionale. Aproape toti fungii prezinta miceliul vegetativ, care creste in grosimea substratului si are rol absorbant al substantelor nutritive. Unele specii prezinta structuri specializate denumite rizoizi, care absorb nutrientii, dar au si rol de fixare. Miceliul care creste deasupra substratului este miceliul aerian, pe care se dezvolta structurile cu rol reproducator. Cele doua tipuri de miceliu pot sa se dezvolte simultan, formand un aparat vegetativ unitar, sau aparitia lor este decalata in timp. Miceliul asimilator este bine dezvoltat si patrunde adanc in substrat. Structurile reproducatoare sunt totdeauna aeriene si pot fi microscopice sau macroscopice. La levuri, aceiasi celula indeplinesse ambele functii.
Dimorfismul fungic. Cativa fungi patogeni prezinta un dimorfism evident, adica doua aspecte morfologice distincte, in conditii diferite de mediu. Pe medii nutritive, aceste organisme manifesta tipul morfologic normal, caracteristic nutritiei saprobionte, dar in tesuturile animale sau pe medii foarte bogate in substante nutritive, la temperaturi superioare, aparatul vegetativ are caracteristicile unei levuri. Denumirea de "saprobionta" se foloseste pentru varianta morfologica filamentoasa, iar cea de "parazitara" se foloseste pentru stadiul de levura.
S. cerevisiae sufera o tranzitie dimorfica, de la morfologia de levura, la forma filamentoasa, ca raspuns la semnalele nutritionale din mediu, in special limitarea sursei de azot si excesul sursei de carbon fermentescibil. Celulele diploide sufera tranzitia dimorfica la forma filamentoasa, denumita diferentiere pseudohifala. Cresterea filamentoasa reprezinta o schimbare ampla a morfologiei cresterii celulare. Celulele devin alungite, se comuta la modul de inmugurire unipolara, raman atasate una de alta si invadeaza substratul de crestere, ceea ce ii permite utilizarea nutrientilor in conditii defavorabile
Cresterea pseudohifala invaziva se produce ca raspuns al tulpinilor
diploide, la abundenta sursei de C si limitarea sursei d
In mediile naturale, cele mai multe tulpini de S. cerevisiae sunt diploide, iar starea haploida a ciclului este reprezentata de gameti
Fungii se reproduc pe cale sexuata si asexuata. Cele doua cai nu sunt comune tuturor fungilor, deoarece unele grupe se reproduc numai pe cale asexuata.
Unitatea structurala de reproducere, tipica pentru fungi este sporul cu unul sau mai multi nuclei. Formarea sporilor este o modalitate comuna pentru reproducerea, dispersia si supravietuirea in conditii nefavorabile. Pentru fungii patogeni, sporii sunt sursa majora de infectie a gazdelor.
Sporii, sexuati sau asexuati sunt produsi pe hife, in interiorul lor sau in structuri specializate. Formarea sporilor sexuati implica imperecherea hifelor de polaritate sexuala opusa si diviziunea meiotica, iar sporii asexuati se formeaza prin diviziuni mitotice.
La fungii macroscopici se formeaza structuri reproducatoare multicelulare ce poarta spori, denumite sporocarpi.
Sporul este o structura ce poate fi separata de talul parental si astfel este diseminat.
Tipul de spori produsi si modalitatea de sporulare sunt criterii importante pentru clasificarea fungilor. In general, sporii sexuati nu sunt mai termorezistenti decat celulele vegetative.
Reproducerea sexuata poate sa fie rezultatul unirii hifelor, a celulelor sexuale diferentiate (gameti), a unor structuri multinucleate cu polaritate sexuala denumite gametangii sau a unui gametangiu femel cu un gamet mascul imobil.
Toate aceste structuri reproducatoare au nuclei cu un set haploid de cromosomi. Unirea elementelor reproducatoare mascule si femele realizeaza intr-o prima etapa, fenomenul de plasmogamie, ce semnifica unirea maselor citoplasmatice, dar cei doi nuclei haploizi raman separati si constituie stadiul de celula dicariotica. Faza dicariotica este, probabil, prezenta la toate organismele care se reproduc sexuat, dar uneori are o durata foarte scurta.
La majoritatea fungilor superiori (Bazidiomicete), faza dicariotica se extinde pe cea mai mare parte a ciclului de viata si se prelungeste mai multe generatii celulare, timp in care reproducerea se face numai pe cale asexuata.
Cand cei doi nuclei (mascul si femel) fuzioneaza in procesul de cariogamie, rezulta un zigot diploid, la care prima diviziune este meiotica si se formeaza sporii sexuati haploizi. Asadar, formarea sporilor sexuati este precedata de fuziunea nucleara si de diviziunea meiotica. Celulele sporale formate pe cale sexuata se numesc ascospori, bazidiospori, oospori si zigospori. Sporii sexuati se formeaza cu o frecventa mai mica decat cei asexuati.
Sporularea sexuata este indusa in conditii speciale de mediu.
Ascosporii se formeaza in celule saciforme, denumite asce, dupa fuziunea citoplasmatica si nucleara a celulelor polarizate sexual. Sporii sunt structuri individualizate, de obicei in numar de 8 pentru o asca, sferici sau ovali, nerezistenti la factorii de mediu.
Bazidiosporii se formeaza la capatul unor structuri dilatate, denumite bazidii. Sunt solitari, sferici sau ovali, in numar de 4, fara proprietati de rezistenta (de exemplu, la Amanita, Agaricus, Coprinus).
Oosporii se dezvolta intr-o celula ou, denumita oogonie. Sunt sferici, in numar de 1-20 intr-o oogonie, mai rezistenti decat majoritatea sporilor asexuati (de exemplu, la Saprolegnia).
Zigosporii sunt mari, cu pereti grosi, sferici sau ovali, nerezistenti. Se formeaza un singur zigospor (de exemplu, la Rhizopus).
Sporii asexuati. Unele grupe de fungi produc atat spori sexuati cat si asexuati, iar altele produc numai spori asexuati. Formarea sporilor asexuati poate avea loc in faza haploida, in faza dicariotica sau in faza diploida a ciclului sexual. Spre deosebire de sporii sexuati, sporii asexuati nu sunt niciodata precedati de diviziunea meiotica.
Sporii asexuati includ o diversitate de structuri, cu denumiri diferite in functie de modalitatea de formare: artrospori, blastospori, clamidospori, conidii, sporangiospori, zoospori.
Unele grupe de fungi au capacitatea de a forma doua tipuri de spori asexuati. Sporii asexuati difera prin aspectul morfologic si dimensiuni, culoare, prin numarul de nuclei.
Artrosporii se formeaza prin fragmentarea hifelor si au forma cilindrica sau sferica. Nu sunt rezistenti la actiunea factorilor de mediu. Sunt definitorii pentru genurile Geotrichum, Trichosporon etc.
Blastosporii se formeaza prin inmugurirea celulei principale. Au forma sferica sau ovalara si sunt nerezistenti. Sunt caracteristici pentru genurile Candida, Saccharomyces etc.
Clamidosporii se formeaza prin dilatarea celulelor terminale sau intercalare ale hifei. Sunt solitari, sferici, au pereti grosi si foarte rezistenti la uscaciune si caldura. Se diferentiaza la Candida, Mucor etc.
Conidia se formeaza ca o structura solitara sau in siruri moniliforme pe hife specializate, denumite conidiofori. Au forma sferica sau ovoida si nu sunt rezistenti la factorul termic. Se diferentiaza la Aspergillus, Penicillium (si se numesc microconidii) sau sunt multicelulari (macroconidii), lungi si ascutiti, nerezistenti (de exemplu, la Alternaria, Trichophyton).
Fialosporul (o conidie speciala) se formeaza pe ramuri specializate ale conidioforilor, denumite fialide. Fialosporii sunt solitari sferici sau ovalari, nerezistenti. Se diferentiaza la Phialospora.
Sporangiosporii se formeaza in interiorul unor celule dilatate, la capatul hifelor, denumite sporangi. Sunt sferici, nerezistenti (de exemplu, la Mucor, Rhizopus etc.).
Zoosporii se formeaza in interiorul zoosporangilor, diferentiati la capatul hifelor principale sau al ramificatiilor laterale. Sunt sferici, nerezistenti, dar caracterul lor distinctiv este mobilitatea prin flageli, de unde isi iau si denumirea. Se formeaza la fungii inferiori (de exemplu, la Saprolegnia).
Sporii produsi de fungii aquatici sunt mobili, iar cei produsi de fungii terestri sunt inconjurati de pereti mai grosi decat ai hifei. Peretele sporal este alcatuit din epispor si endospor, care acopera protoplastul sporal. Intreaga structura poate fi acoperita de perispor.
In conditii favorabile de mediu (substante nutritive, umiditate, pH, temperatura), sporii fungici germineaza si produc una sau mai multe structuri filamentoase denumite tuburi de germinare (fig. 146). Ele emerg in zonele mai subtiri ale invelisului sporal. Apoi cresc, se alungesc si se ramifica pentru a forma hifele.
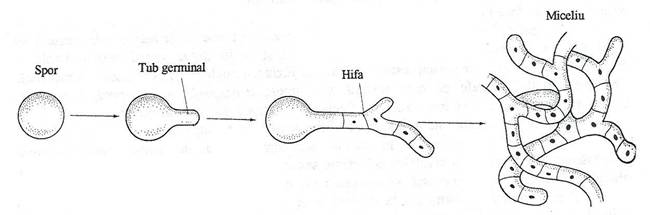
Fig. 146. Germinarea sporilor fungici. Initial se formeaza tubul de germinare, care creste si rezulta o hifa. Hifa creste, se ramifica si formeaza o panza hifala, denumita miceliu.
Criteriile majore de clasificare a fungilor sunt urmatoarele:
modul de reproducere;
ciclul de viata
morfologia organelor de reproducere si a sporilor formati; particularitati de ordin biochimic, fiziologic, determinari de ordin calitativ a asemanarilor si deosebirilor dintre diferiti reprezentanti.
Fungii sunt considerati ca unitate taxonomica de sine statatoare, cu rangul de regn: regnul Fungi (Mycetae), propus de Ainsworth (l973).
Fungii inferiori formeaza zoospori si includ clasele Oomycetes si Chytridiomycetes.
Fungii din diviziunea Amastigomycota nu produc zoospori si sunt reprezentati de clasele: Zygomycetes, Ascomycetes, Basidiomycetes, Deuteromycetes.
Regnul Fungi cuprinde si un grup de organisme cu particularitati comune fungilor si protozoarelor: Mixomycete (Gymnomyxa).
Fungii inferiori constituie un grup heterogen, care au fost reuniti in doua clase neinrudite: Oomycetes si Chytridiomycetes.
Majoritatea fungilor inferiori traiesc in mediul aquatic si intr-un anumit stadiu al ciclului de viata formeaza o celula mobila, denumita zoospor. La originea sa poate fi un proces sexuat sau unul asexuat. Zoosporul participant la un proces sexuat are functia unui gamet.
Zoosporii sunt mobili prin unul sau doi flageli. Flagelul este un apendice filiform, alcatuit din doua fibre centrale si 9 fibre periferice, contractile.
Zoosporii si gametii mobili se formeaza intr-o structura in forma de sac, denumita zoosporangiu. Daca zoosporul are rol de gamet, sacul se cheama gametangiu.
Oomycetes
Oomycetes sau fungii filamentosi de apa se deosebesc de fungii propriu-zisi, deoarece au pereti celulozici. Cromosomii lor contin proteine histonice, in timp ce fungii adevarati au histone putine sau deloc. Un alt caracter distinctiv fata de fungii propriu-zisi, consta in prezenta centriolului, absent la fungi. Reprezentantii lor traiesc mai ales in apa dulce. Unii sunt terestri. Cel mai reprezentativ este Plasmopara viticola, agentul care produce mana vitei de vie. Amestecul Bordeaux, format din CuSO4 si Ca(OH)2 este foarte eficient pentru controlul acestei boli cu un potential distructiv urias.
Ciclul de reproducere include doua stadii: sexuat si asexuat. Sporii asexuati sunt biflagelati si mobili. Gametii sunt imobili si prin fuziune genereaza zigotul 2n, denumit oospor.
Zygomycetes
Zygomycetes se caracterizeaza printr-un tip de reproducere sexuata, in care doua gametangii fuzioneaza si formeaza un zigot (zigospor). Hifele au putini pereti transversali sau acestia lipsesc complet, miceliul fiind de tip coenocitic(fig. 147).
Un reprezentant comun al grupului este Rhizopus nigricans, care creste pe resturi alimentare pe care le acopera cu un miceliu de culoare inchisa (mucegaiul negru). Unele hife se raspandesc orizontal pe suprafata substratului si se numesc stolonifere. Din acestea se dezvolta hife care penetreaza substratul nutritiv, avand rol in absorbtia nutrientilor si de fixare. La extremitatea unor hife care cresc vertical se formeaza o dilatare in forma de sac, denumita sporangiu. In fiecare sporangiu se dezvolta o aglomerare de spori asexuati, care se elibereaza prin ruperea peretelui sporangial.
Cresterea acestui mucegai in si pe produsele de panificatie este stopata prin adaugarea agentilor inhibitori, la aluatul de crestere. Unele zigomycete produc infectii la om si animalele vertebrate. Altele paraziteaza viermii si artropodele.
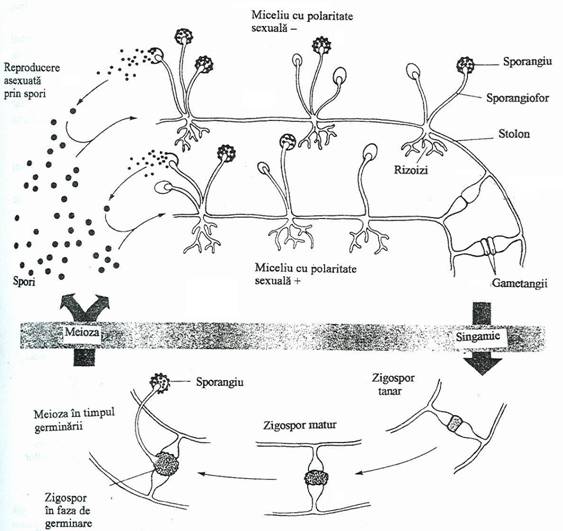
Fig. 147. Ciclul reproducerii sexuate si asexuate la Rhizopus stolonifer (mucegaiul negru al painii).
Ascomycetes
Ascomicetele formeaza cel mai mare si diversificat grup de fungi, cu circa 2000 de genuri. Grupul include levurile, fungii filamentosi care produc boli caracteristice la plante denumite fainari si fungi cu palarie: Morchella (zbarciogul) si Tuber (trufele).
Majoritatea sunt specii terestre, dar un numar considerabil, populeaza apele dulci sau sarate. Majoritatea sunt saprobionte pe resturile vegetale in descompunere, iar unele sunt parazite sau traiesc in asociatii reciproc benefice, cu alte microorganisme sau cu unele plante.
Multe ascomicete saprobionte sunt foarte specializate si cresc numai pe anumite parti in descompunere ale unor specii de plante, iar cele parazite sunt restranse la anumite tesuturi ale gazdei (de exemplu, petiolul frunzei). Un numar important de reprezentanti ai grupului sunt paraziti ai plantelor si mai putini ai insectelor sau altor animale.
Ascomicetele se reproduc pe cale sexuata si asexuata. Caracteristica acestui grup de fungi este ca sporii sexuati, denumiti ascospori se formeaza intr-o structura in forma de sac, denumita asca.
Asca este o celula care se formeaza prin procesul de fuziune a unor structuri tubulare a doua celule invecinate haploide, polarizate sexual. Initial fuzioneaza numai masele citoplasmatice, prin procesul de plasmogamie. Plasmogamia poate fi rezultatul unirii a doua celule similare sau hife, ori prin unirea heterogena a doua celule diferite, diferentiate, denumite gametangii, unul mascul si unul femel sau a unui gametangiu femel cu o celula masculina detasata (spermatie) (fig. 148).
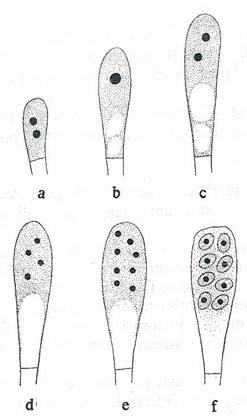
Fig. 148. Stadiile succesive ale formarii ascei. a. Celula binucleata rezultata prin fuziunea celulelor haploide. b. Fuziunea nucleara. c, d, e. Diviziuni nucleare. f. Formarea ascosporilor.
Dupa plasmogamie, celula are doua cai de evolutie:
cariogamia poate avea loc imediat dupa plasmogamie si rezulta zigotul. Zigotul sau celulele rezultate prin diviziunea sa, se transforma in asca;
cariogamia intarzie si starea dicariota, initiata prin procesul plasmogamiei, se prelungeste pentru mai multe generatii.
In interiorul ascei, procesul generarii ascosporilor este asemanator la toti fungii. Dupa cariogamia imediata sau tardiva, nucleul diploid al ascei, sufera diviziunea meiotica. Dupa prima diviziune meiotica, se formeaza doi nuclei, iar dupa a II-a diviziune rezulta 4 nuclei. La cele mai multe ascomicete, urmeaza o a II-a diviziune mitotica si se formeaza 8 nuclei. Cei 8 nuclei sunt incorporati in ascospori, delimitati de o membrana dubla. Membrana este originara in reticulul endoplasmic sau in invelisul nuclear. Ea se invagineaza pentru a inconjura fiecare nucleu. Peretele sporal nou, este depus intre cele doua lame membranare. Membrana interna devine plasmalema sporului. Numarul ascosporilor poate fi numai de 4 daca cea de a II-a diviziune mitotica nu mai are loc, sau de l6, 32 sau alt multiplu de 8, in functie de numarul diviziunilor mitotice. La unele ascomicete, ascele se pot forma in corpi de fructificatie, denumiti ascocarpi. Se cunosc patru tipuri de astfel de structuri: apotecia, cleistotecia, loculotecia si peritecia.
Reproducerea asexuata este comuna majoritatii ascomicetelor. Pe medii copiotrofe, celulele de levuri se reproduc asexuat prin inmugurire, iar pe medii oligotrofe, celulele diploide rezultate prin fuziune, sporuleaza (fig. 149).
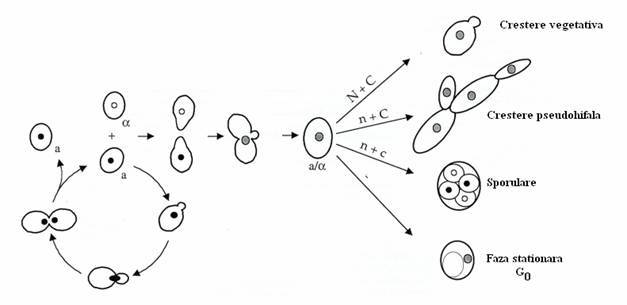
Fig. 149. Ciclul de viata la S. cerevisiae. Pe mediul bogat in nutrienti, celulele haploide de polaritate sexuala opusa, se imperecheaza si fuzioneaza. Rezulta celule diploide, cu evolutie diferita in functie de disponibilitatea nutrientilor: N + C = mediu nutritiv bogat in surse de N si de C fermentescibile; n + C = sursa de N este limitata, iar cea de C fermentescibila este abundenta; n + c = surse limitate de N si de C nefermentabil; - mediu fara nutrienti( dupa K. Lengeler, 2000).
In acest proces, nucleul se divide prin mitoza, fara dezorganizarea membranei nucleare. Unul dintre cei doi nuclei, se deplaseaza intr-o mica proiectie a citoplasmei, delimitata de membrana si perete, denumita mugure.
Reproducerea asexuata prin conidii este comuna. Ele se pot forma libere pe miceliu sau sunt organizate in corpi de fructificatie, asemanatori ascocarpilor. Adeseori, stadiul conidial si cel de asca, apar pe micelii separate. Miceliul formator de conidii se mai numeste stadiul imperfect, datorita absentei modalitatii de reproducere sexuata, iar miceliul formator de asce se numeste stadiul perfect, pentru ca este sediul proceselor sexuale.
Formarea sporilor asexuati la Asp. nidulans. Ciclul de sporulare asexuata la A. nidulans poate fi impartit in 2 faze (fig. 150):
o faza de crestere, in care celulele devin competente pentru a raspunde la semnalele inductoare ale sporularii:
o faza de reproducere asexuata, in care se initiaza sporularea si se formeaza structurile sporifere.
Aceasta ciuperca creste formand o retea ordonata de hife ce constituie un miceliu. Fiecare hifa este formata din celule vegetative haploide, multinucleate si separate de septuri perforate, ce permit migrarea nucleilor. Hifa creste prin extensie apicala si se ramifica, formand o colonie cu simetrie radiala.
Hifa are rolul de a absorbi nutrientii din mediul de crestere, facilitata de secretia enzimelor degradative. Dupa o perioada fixa de timp, ce urmeaza cresterii vegetative, unele celule hifale din centrul miceliului produc ramificatii aeriene ce initiaza sporularea asexuata. Se formeaza structuri multicelulare denumite conidiofori, ce poarta lanturi de spori denumite conidii. Este o trecere de la cresterea hifala, inalt polarizata, la cresterea prin inmugurire si tranzitia de la starea multinucleata la cea uninucleata.
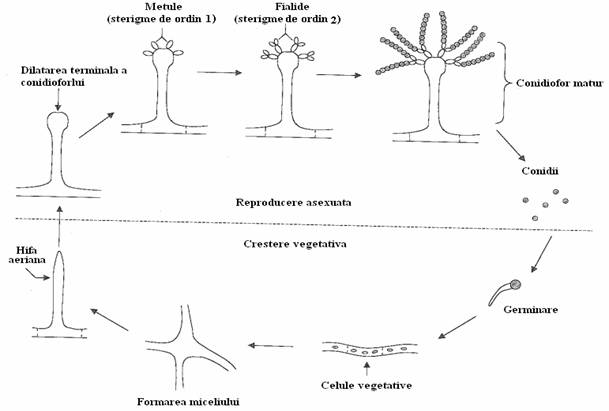
Fig. 150. Ciclul formarii sporilor asexuati la Asp. nidulans. In faza de crestere vegetativa, sporii germineaza si se formeaza hife miceliene. In faza reproductiva, celule hifale specializate din miceliul primar dau nastere hifelor aeriene specializate denumite conidiofori. Hifele aeriene se termina cu o vezicula, ce inmugureste si se ramifica, rezultand sterigme de ordinul 1 si 2. Prin diviziuni mitotice succesive, sterigmele de ordinul 2 (fialide) formeaza lanturi de conidii care pot germina pe un mediu favorabil si reiau ciclul (dupa Lengeler, mmbr 2000).
Ciclul sexuat. In conditii de deficit nutritiv, hifele multinucleate si ramificate formeaza un miceliu multicelular. In ciclul vegetativ, N. crassa produce doua tipuri de spori vegetativi: macro- si microconidii. Macroconidiile sunt multinucleate si se diferentiaza din hife aeriene specifice denumite conidiofori, ce cresc perpendicular pe suprafata miceliului. Dupa o perioada de crestere aeriana apicala, hifele aeriene se comuta la modul de crestere prin inmugurire, ce duc la formarea lanturilor de proconidii in interiorul conidioforului. Pe masura ce se matureaza, se separa si sunt dispersate in primul rand de curentii de aer. In conditii favorabile, germineaza rapid (fig. 151).
Microconidiile se diferentiaza din miceliul vegetativ primar (de substrat), sunt mai mici, uninucleate si au capacitate redusa de germinare.
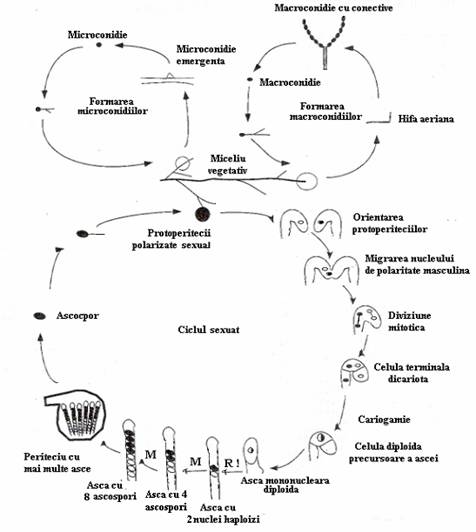
Fig. 151. Ciclul de viata la N. crassa. Ca raspuns la conditiile de mediu, miceliul vegetativ produce conidii (macro si microconidii) si protoperitecii. Macroconidiile se formeaza din conidiofori ce se diferentiaza din hifele aeriene, iar microconidiile se formeaza direct pe hifele vegetative. Dupa fertilizarea protoperiteciei de polaritate feminina cu un nucleu al protoperiteciei de polaritate masculina, este initiat ciclul sexual in care se produc ascospori (adaptat dupa Lengeler, mmbr, 2000).
Ciclul sexual. La fungi, meioza este strans asociata cu sporularea. Dezvoltarea sexuata a ascei se caracterizeaza unui corp de fructificare ce contine asce cu celule precursoare sporale. In contrast, levurile formeaza asca pe cale directa, dintr-o singura celula diploida, fara sa formeze corpul de fructificare.
N. crassa este heterotalica si are doua tipuri de hife miceliene, polarizate sexual: A si a. Ciclul sexual se initiaza ca raspuns la epuizarea sursei de N. Reproducerea sexuata are loc numai intre tulpini de polaritate sexuala opusa (mat A si mat a).
Celulele vegetative A, in conditiile limitarii sursei de N, produc precursori ai corpilor de fructificare, denumite protoperitecii. Din protoperitecii cresc hife specializate polarizate sexual (gameti feminini), care manifesta crestere chemotropa spre celule de polaritate masculina (condii sau hife), ca raspuns posibil la feromoni. Contactul dintre trichogin si celula de polaritate masculina duce la intrarea nucleului masculin in trichogin sI transportul sau ulterior in ascogonul protoperiteciului. Odata ce nucleul unui gamet mascul intra in protoperiteciu, acesta devine periteciu- o hifa ascogena dicariota, pentru ca cei doi nuclei de polaritate sexuala opusa nu fuzioneaza imediat, dar se divid mitotic pentru a produce o masa hifala ascogena. Urmeaza fuziunea celor 2 nuclei de polaritate opusa. Se formeaza celule diploide, care intra in diviziune meiotica urmata de 1-2 diviziuni mitotice, rezultand asce cu 4- 8 ascospori.
Bazidiomicetele sunt fungii cei mai evoluati. Miceliul este pluricelular si se distinge prin organul sporifer denumit bazidie, in care se formeaza bazidiosporii.
Grupul bazidiomicetelor include 20-30 000 de specii, cu morfologie diferita a talului, in functie de modul de viata: saprobionte sau parazite. Din acest grup fac parte fungii gelatinosi, fungii care produc boli la plante (rugini, taciuni, maluri), fungii consola, care se dezvolta pe scoarta copacilor, fungii macroscopici comestibili si necomestibili.
Denumirile comune se refera la partea vizibila a ciupercii, denumita bazidiocarp, sustinut de un miceliu asimilator bine dezvoltat, care penetreaza substratul (solul sau materialele vegetale), pe care ciuperca fructifica.
Reproducerea sexuata. Structura purtatoare de spori sexuati a bazidiomicetelor este o celula specializata, cu aspect maciucat, care se numeste bazidie. La varful ei se dezvolta 4 bazidiospori, care proemina la exteriorul bazidiei. Sporii sunt eliberati si dupa germinare se dezvolta un miceliu. De cele mai multe ori, bazidiile se formeaza pe corpuri de fructificare (bazidiocarp).
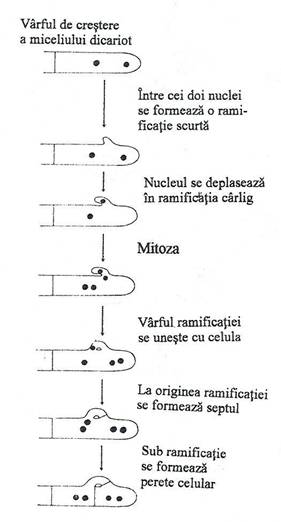
Fig. 152. Modalitatea diviziunii celulare intr-o hifa dicariotica de bazidiomicete. In acest fel, fiecare dintre cei doi nuclei distincti se distribuie in cele doua celule. Structura caracteristica are forma de carlig.
Cele mai multe bazidiomicete produc bazidii, organe de reproducere omologe ascei si probabil derivate din ea. Bazidia se formeaza intr-o structura hifala binucleata, sediul evenimentelor definitorii ale reproducerii sexuate: cariogamia si meioza. Spre deosebire de asca, bazidia poarta bazidiosporii la exteriorul ei.
Bazidiosporul contine un nucleu haploid. Dupa germinare, din bazidiospor se dezvolta un miceliu haploid monocariotic, septat, format din celule uninucleate. Faza miceliului uninucleat haploid este scurta, deoarece curand se produce plasmogamia, care semnifica fuziunea a doua hife monocariote. La bazidiomicete, organele sexuale lipsesc.
Celula dicariotica, formata prin plasmogamie, continua sa se divida, dar isi pastreaza starea binucleata. Se formeaza un miceliu asimilator care penetreaza in substrat.
Cand conditiile de mediu sunt favorabile procesului de reproducere, miceliul dicariotic sufera o morfogeneza complexa, pentru a forma un bazidiocarp. Unele dintre celulele bazidiocarpului se transforma in bazidii. Celulele din care se formeaza bazidiile (probazidii) sunt dicariote, avand doi nuclei haploizi. Cei doi nuclei ai probazidiei fuzioneaza si formeaza un nucleu diploid, care se divide meiotic. De obicei, bazidia dilatata formeaza la extremitatea ei libera, 4 proiectii alungite denumite sterigme. Varful fiecarei sterigme se dilata pentru a forma bazidiosporul. In sterigma, migreaza un singur nucleu haploid si intreaga structura devine bazidiospor. Bazidiosporii, de obicei, contin un singur nucleu (fig. 153).
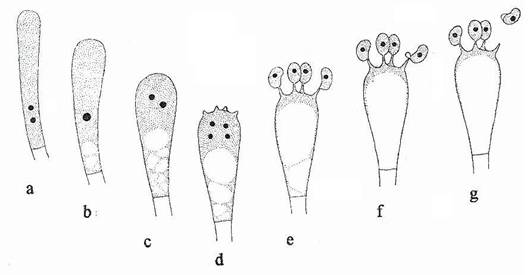
Fig. 153. Stadiile succesive ale formarii bazidiei si eliberarii bazidiosporilor. a. O celula binucleata. b. Fuziunea nucleilor. c, d. Diviziunea nucleara. e. Formarea bazidiosporilor. f, g. Eliberarea bazidiosporilor.
Rareori, in bazidiospori sunt incorporati doi nuclei si se formeaza o bazidie care contine numai doi bazidiospori binucleati.
Corpii de fructificare (bazidiocarpi) sunt caracteristici tuturor fungilor macroscopici, dar lipsesc la cei microscopici (taciuni, maluri, rugini).
Bazidiosporii formati pe corpul de fructificare, se disperseaza si initiaza cresterea miceliului. Miceliul haploid poate sa creasca extensiv, dar uneori, prin fuziunea a doua micelii haploide se formeaza miceliul dicariotic, care formeaza corpul de fructificatie.
In anumite conditii de mediu, hifele dicariote cresc si formeaza structuri butonate submerse. Acestea sunt primordiile corpilor de fructificare. Butonii pot sa ramana perioade lungi de timp, in stare latenta. In conditii de umiditate optima, butonii cresc rapid in cateva ore sau 1-2 zile, pe seama acumularii apei si se formeaza corpi de fructificare. De multe ori, pe o suprafata de sol bine delimitata, se produce o adevarata "explozie" a formarii corpilor de fructificare.
Reproducerea asexuata este comuna la bazidiomicete, desi este mai putin frecventa decat la fungii inferiori si ascomicete. Structurile reproducatoare asexuate la bazidiomicete sunt mugurii (ca si la levuri), conidiile, clamidosporii si oidiile. Oidiile se formeaza prin fragmentarea miceliului monocariotic: o hifa se fragmenteaza in celulele sale componente. Oidiile sunt haploide si prin germinare se formeaza un miceliu haploid. Miceliile derivate din oidii, fuzioneaza si formeaza un miceliu dicariotic.
Ciclul de viata la Ustilago maydis. Celula asexuata are aspect usor alungit este saprobionta pe materia organica si creste prin inmugurire. Doua celule cu polaritate sexuala opusa, dezvolta protruzii filamentoase, denumite tuburi de conjugare, fuzioneaza si formeaza o hifa dicariota infectioasa. Hifa este patogena si infecteaza tesutul meristematic al oricarei parti aeriene a plantei. Proliferarea hifala in tesuturile plantei induce cresterea neoplazica si formarea galelor. Hifele fungice, in interiorul acestor structuri tumorale, se rotunjesc si se produce cariogamia, dand nastere la teliosporii diploizi uninucleati. Prin spargerea tesutului tumor-like, sporii se disemineaza in mediu. Ca raspuns la conditiile favorabile de mediu, are loc meioza si teliosporii germineaza, formand un promiceliu ce consta din 4 celule haploide, care include si teliosporul original. In final, bazidiosporii inmuguresc din promiceliu si germineaza, reluand ciclul (fig. 154).
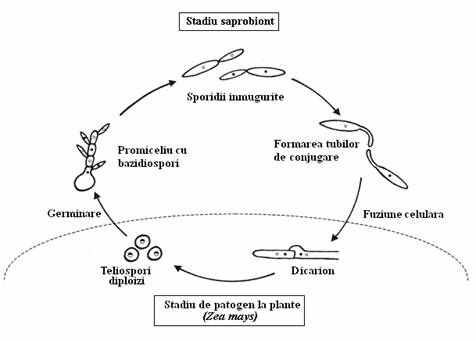
Fig. 154. Ciclul de
viata
Deuteromycetes (Fungi Imperfecti)
Grupul include circa l5 000 de specii de fungi, carora, aparent le lipseste stadiul sexuat de reproducere si din aceasta cauza se numesc fungi imperfecti. Se presupune ca majoritatea organismelor cuprinse in acest grup sunt, stadiile asexuate ale fungilor care se reproduc sexuat, ce apartin grupelor Ascomycetes si Bazidiomycetes.
Ca grup, deuteromicetele sunt heterogene, speciile componente neavand origine comuna si nici raporturi filogenetice.
La unii reprezentanti ai grupului s-a identificat existenta unei faze sexuate a ciclului de viata. In astfel de situatii, ambele stadii de dezvoltare a ciupercii (sexuat sau perfect si asexuat sau imperfect) sunt transferate la grupul caruia ii apartine stadiul perfect, in acord cu conceptia ca, impreuna, cele doua stadii formeaza ciclul complet de viata al ciupercii.
Membrii grupului cresc pe orice tip de substrat. Unele sunt saprobionte pe materia organica vegetala in descompunere, iar altele sunt parazite la plante si animale.
In acest grup sunt inclusi fungii filamentosi usor cultivabili pe medii artificiale. Unii produc substante utile pe care se bazeaza industria micologica: antibiotice, acizi organici sau sunt folositi pentru obtinerea unor specialitati de branzeturi.
Miceliul poate fi de tipul ascomicetelor, cu septuri intercelulare perforate, sau de tipul bazidiomicetelor, la care se formeaza septuri complexe si conexiuni "carlig".
Reproducerea se face prin fragmente miceliene sau prin spori. Unii membri ai grupului nu formeaza spori. Sporii pot fi oidii, clamidospori sau conidii. Conidiile se formeaza pe hife modificate si specializate, denumite conidiofori, diferentiate din miceliul vegetativ. Conidioforii pot fi neramificati sau ramificati, cu grosime uniforma sau dilatati la varf. Conidiile se pot forma la varf (pentru cei neramificati) sau in diferite puncte pe traseul hifei specializate. Conidiile sunt colorate ca si conidioforii. Ele sunt sferice, eliptice, curbate, rasucite, in forma de secera. De cele mai multe ori, conidiile sunt unicelulare, dar uneori pot avea doua sau mai multe celule.
Myxomycetes (Gymnomyxa)
Grupul cuprinde o varietate larga de organisme, care au fost clasificate ca fungi sau protozoare, caracterizate prin mai multe proprietati comune, dar care se deosebesc de fungii adevarati. Aparatul lor vegetativ este o masa amoeboida, asemanatoare cu un protozoar, dar corpul de fructificare se aseamana cu al fungilor si aici se formeaza spori cu perete celular.
Aparatul vegetativ al mixomicetelor este o masa celulara nuda, denumita plasmodiu, mobila pe suprafata unui substrat solid. Plasmodiul este alcatuit din celule amoebiene (denumite mixamoebe), lipsite de perete celular. Structurile reproducatoare formeaza corpi de fructificare, asemanatori cu ai fungilor.
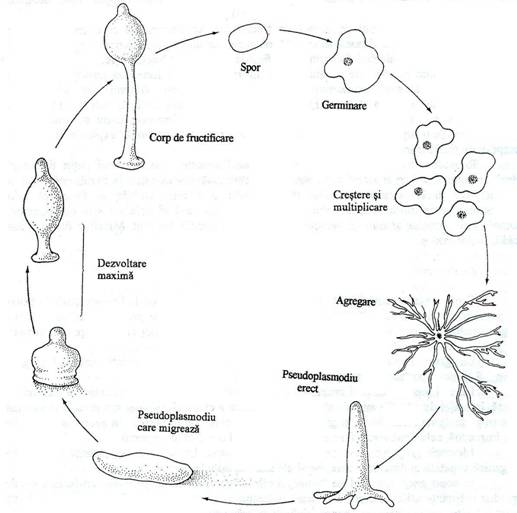
Fig. 155. Ciclul de viata la Dictiostelum discoideum.
Se cunosc circa 500 de specii de mixomicete. Sunt organisme heterotrofe si se hranesc cu materia vegetala in descompunere. Corpii de fructificare sunt viu colorati si apar frecvent pe lemnul in descompunere din padurile dense, cu grad mare de umiditate.
Unele sunt parazite. De exemplu, Plasmodiophora brassicae produce boli la conopida, ridiche, nap, caracterizate prin deformarea radacinilor.
Mixomicetele sunt divizate in doua grupe: celulare si plasmodiale.
Cele cu organizare celulara au un aparat vegetativ alcatuit din celule individualizate, asemanatoare amoebelor. Aparatul vegetativ al mixomicetelor plasmodiale este format dintr-o masa de protoplasma nediferentiata, de diferite aspecte si dimensiuni, comparabil cu o amoeba giganta plurinucleata. Se deplaseaza amoeboidal.
Mixomicetele se caracterizeaza printr-un ciclu de viata neobisnuit, cu alternanta unor structuri de aspect morfologic foarte variat. Cele de tip celular traiesc si se multiplica in sol, sub forma unor celule amoebiene, care se hranesc cu bacterii pe care le fagociteaza. Dupa epuizarea sursei de hrana, celulele se agrega (dar nu fuzioneaza) in structuri multicelulare (pseudoplasmodii).
Cel mai cunoscut reprezentant este Dictyostelium discoideum. Agregarea celulelor este produsa de acrasina, o substanta identificata ca AMPc, cu efect chimiotactic pentru mixomicete. Pseudoplasmodiul manifesta fototactism si se deplaseaza spre sursa de lumina. Dupa incetarea deplasarii, pseudoplasmodiul formeaza corpul de fructificare (fig. 155).
Agregatul multicelular (pseudoplasmodiul), prin procese morfogenetice complexe formeaza un corp de fructificare colorat. Acesta este un sporocarp, format dintr-un disc bazal, un peduncul si o capsula plina cu spori, la extremitatea pedunculului. Sporii sunt celule reproducatoare asexuate. In conditii de umiditate, sporii germineaza si rezulta celule individualizate denumite mixamoebe, care in privinta modului de hranire si locomotie sunt asemanatoare cu amoebele. Se divid prin fisiune simpla. In conditii nefavorabile de mediu, ele se agrega in pseudoplasmodiu si ciclul se reia.
Fungii au un rol deosebit de important in natura, deoarece, fiind descompunatori ai materiei organice, redau elementele biogene, circuitului natural. Fungii descompun toate categoriile de macromolecule caracteristice lumii vii: polizaharide, acizi organici, lignine, hidrocarburi aromatice si alifatice, zaharuri, alcooli, aminoacizi, purine, lipide, proteine, acizi nucleici. O specie fungica oarecare poate sa descompuna numai anumite componente ale tesuturilor vegetale si animale. Descompunerea substratului necesita actiunea cooperanta a mai multor specii de microorganisme. Actiunea degradativa a fungilor produce mari pierderi ale fructelor si altor produse vegetale in depozite.
Unii reprezentanti ai fungilor au o importanta economica de prim ordin in bioindustria micologica (producerea antibioticelor, a acizilor organici etc.), iar fungii paraziti prezinta atat interes medical, dar in special pentru productia agricola.
Ascomicetele au o importanta economica deosebita, atat prin potentialul lor distructiv, cat si prin efectele benefice. Levurile sunt utilizate in producerea industriala a bauturilor alcoolice de fermentatie si in industria panificatiei, iar Claviceps purpurea (cornul secarei) este folosita ca sursa de medicamente (ergotina).
Morchella (zbarciogul) si Tuber (trufe) sunt ascomicete comestibile. Tuber este o ciuperca sferica subterana, care creste in jurul anumitor specii de stejar. Savoarea unica a acestor doua specii, le face sa fie foarte cautate de gastronomi, desi valoarea lor nutritiva este scazuta.
Claviceps purpurea este parazita si creste, in special, pe plantele de secara, dar si pe alte plante cerealiere. Din ergotina (un alcaloid toxic), se extrag medicamente utilizate in tratamentul afectiunilor vasculare, precum si cunoscutul halucinogen LSD (dietil-amida acidului lisergic).
Dintre macromicete, Agaricus bisporus este cultivata in amenajari speciale.
Dintre deuteromicete, cei mai importanti sunt membrii g. Penicillium. Unele specii de Penicillium sunt producatoare de penicilina. P. roqueforti si P. camemberti confera savoarea distincta si consistenta moale a branzeturilor de Roquefort si Camembert, datorita activitatilor enzimatice ale fungilor specifici. Miceliul se dezvolta preponderent la suprafata, dar patrunde si in substrat si prin actiunea sa enzimatica ii diminua consistenta si ii confera o savoare unica.
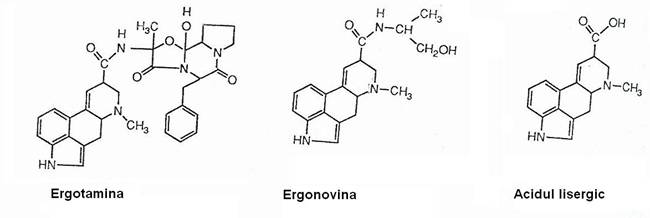
Alcaloizi cu importanta medicala: ergotamina si ergonovina. Acidul lisergic produs de microorganisme este modificat chimic pentru prepararea la scara comerciala a ergonovinei.
Fungii patogeni. Circa 50 de specii de fungi sunt patogene pentru om si animale. Majoritatea fungilor patogeni sunt agenti infectiosi oportunisti, care invadeaza tesuturile organismelor cu rezistenta scazuta la infectie.
Infectia fungica a tesuturilor umane si animale se produce pe fondul unor conditii locale favorizante. Din aceasta cauza, transmiterea infectiei de la un organism la altul este rara. In general, maladiile fungice nu sunt contagioase.
Infectiile produse de deuteromicete se numesc micoze si pot fi impartite in doua categorii mari:
infectii sistemice sau profunde, in care agentul patogen este larg diseminat in organism si creste in diferite organe si tesuturi;
micoze superficiale, care includ infectii ale pielii, parului, unghiilor. Unii fungi produc numai un tip de infectie, iar altele pe amandoua.
Pentru majoritatea fungilor patogeni nu se cunosc stadii de inmultire sexuata si de aceea pozitia lor sistematica este incerta. Unele micoze sistemice sunt produse de levuri. Altele sunt cauzate de fungi dimorfici, care cresc ca levuri in vivo (la 37o) si ca miceliu la temperatura mai mica, in vitro pe medii artificiale.
Fungii patogeni traiesc in sol si patrund in organism pe cale respiratorie, fiind inhalati odata cu aerul. De la poarta de intrare, agentul infectios se disemineaza in organism si se instituie o infectie cronica, cu simptome foarte variate. Infectiile fatale sunt rare.
Infectiile fungice apar frecvent, ca o consecinta a eliminarii microbiotei normale, prin terapie orala cu antibiotice. Cativa fungi (Cryptococcus neoformans, Coccidioides imitis, Histoplasma capsulatum, Blastomyces dermatitides) produc infectii pulmonare foarte asemanatoare prin manifestarile clinice, cu tuberculoza.
Micozele superficiale sunt cauzate, in primul rand, de fungi micelieni (Microsporum, Trichophyton, Epidermophyton). Ele au afinitate pentru tesuturile care contin keratina (piele, par, unghii) si produc enzima necesara degradarii acestei proteine. Unele cresc pe suprafata firului de par, iar altele la interior. Varsta este un factor important care conditioneaza sensibilitatea la infectie: impetigo al pielii capului este frecventa la copii si rara la adulti, iar infectia interdigitala este frecventa la adulti si rara la copii. Acizii grasi nesaturati din scretia glandelor sebacee ale adultului, au actiune antifungica, ceea ce explica rezistenta mai mare a adultului la infectiile fungice superficiale. Una dintre substantele antifungice, acidul undecilenic, se adauga in compozitia multor unguente, folosite in controlul infectiilor fungice ale pielii.
Infectia interdigitala ste favorizata de umiditatea locala crescuta si este produsa de fungi din g. Epidermophyton.
Fungii care produc infectii superficiale se numesc dermatofite. Majoritatea infectiilor cu fungii dermatofitici se pot trata eficient cu griseofulvina, un antibiotic antifungic produs de Penicillium griseofulvum. Griseofulvina se concentreaza in celulele epidermice, la valori inhibitoare pentru cresterea dermatofitului.
Micozele intermediare sunt considerate acelea care produc leziuni epidermice, subcutanate, ale mucoaselor, plamanului. Infectia se poate extinde si in alte tesuturi, la diferite profunzimi.
Infectia este realizata de inhalarea sporilor, prin contaminarea ranilor sau prin ingestie. Cele mai tipice infectii intermediare sunt candidoza si aspergiloza.
Candidoza este produsa de variate specii de Candida, in special C. albicans, din grupul fungilor imperfecti. In procesul infectios se prezinta sub forma unei levuri, dar formeaza si structuri miceliene. Infectia se localizeaza, in primul rand, la nivelul membranelor mucoase, dar si in piele sau plaman. Infectiile mucoasei bucale a nou-nascutului sunt comune, iar la adult produce infectii bucale, tonsilare, vaginale.
Aspergiloza, cauzata de Aspergillus fumigatus este frecventa la puii de gaina, in primele zile dupa ecloziune, la care produce infectii de amploarea unei epidemii. Miceliul se dezvolta abundent in plaman si respiratia devine imposibila. Produce avortul la vaca, oaie etc. Omul prezinta rezistenta naturala la infectia cu A. fumigatus. Cazurile de imbolnavire sunt rare.
Fungi patogeni pentru plante. Cei mai importanti agenti patogeni ai plantelor, atat in ceea ce priveste numarul, cat si a pierderilor economice produse, sunt fungii. Dintre fungii inferiori (Plasmopara, Phytophtora, Peronospora etc.) au un potential distructiv foarte mare, iar fungii care produc taciunii, malura, ruginile, apartin bazidiomicetelor.
Fungii patogeni infecteaza si produc boli la toate tipurile de plante, de la cele ierboase, pana la cei mai mari copaci. S-au studiat in special agentii patogeni ai plantelor de cultura. Toate partile aparatului vegetativ sunt sensibile la atacul fungilor. In general, o specie fungica are specificitate evidenta pentru anumite parti ale plantei. Leziunile tesuturilor gazdei se datoreaza producerii de toxine, blocarii vaselor ce transporta apa si nutrientii sau inducerii malformatiilor.
Fungii produc o larga varietate de metaboliti secundari, a caror semnificatie adeseori nu este cunoscuta, dar au importanta practica medicala, industriala sau agricola. Unele sunt vatamatoare (micotoxine), altele sunt benefice (antibiotice) pentru om.
Cele mai multe produse secundare sunt sintetizate de organisme cu crestere filamentoasa si au morfologie relativ complexa.
Sinteza metabolitilor secundari, la microorganisme este asociata cu procesele de sporulare.
Metabolitii secundari asociati cu sporularea sunt de 3 categorii:
metaboliti care activeaza sporularea (acidul linoleic, la A. nidulans);
pigmentii structurilor de sporulare (melaninele necesare formarii sau integritatii sporilor sexuati si asexuati);
metaboliti toxici secretati la timpul sporularii (micotoxinele).
Producerea metabolitilor secundari incepe tarziu in timpul de crestere, la intrarea in faza stationara. Conditiile de mediu necesare sporularii si secretiei metabolitilor secundari sunt adeseori asemanatoare si chiar mai stringente decat acelea necesare cresterii vegetative. S-a crezut ca sinteza metabolitilor este obligatorie pentru sporulare, dar unele tulpini sporuleaza chiar in absenta producerii metabolitilor secundari.
Melaninele sunt pigmenti de culoare inchisa care se formeaza prin polimerizarea oxidativa a compusilor fenolici. Se sintetizeaza in timpul sporularii si sunt depozitati in peretele celular.
Pigmentii sunt protectori fata de UV, dar sunt si factori de virulenta.
Micotoxine
Contaminarea produselor alimentare, a cerealelor si chiar a alimentelor preparate, cu fungi din sol este aproape inevitabila. Depozitarea in conditii improprii, favorabile cresterii fungilor, altereaza caracteristicile de gust si miros ale produsului. Cei mai multi fungi filamentosi care cresc pe alimente sunt inofensivi. Dar cresterea unor fungi pe substraturi alimentare este insotita de sinteza si eliminarea unor toxine puternice, denumite micotoxine, care produc imbolnavirea sau chiar moartea omului si animalelor.
Cele mai cunoscute micotoxine sunt aflatoxinele, produse de A. flavus, toxice pentru om si pentru multe specii de animale.
Din punct de vedere chimic, aflatoxinele sunt inele aromatice complexe. In doze mari sunt letale, iar in doze mici, induc tumori hepatice.
Alte micotoxine: ochratoxina (produsa de A. ochraceus), rubratoxina (P. rubrum), patulina (A. clavatus), stachibotriotoxina (Stachybotrys atra).
Factorul esential favorizant al cresterii fungilor in produsele alimentare, in cereale si alimente este umiditatea. Producerea micotoxinelor are cea mai mare frecventa in depozitele de cereale, arahide si nuci, la umiditate mai mare de l5%. In regiunile tropicale, datorita umiditatii permanent crescute, dezvoltarea fungilor in produsele depozitate este greu de controlat. Este posibil ca aparitia endemica a neoplaziilor hepatice sa se datoreze cresterii fungilor si producerii de aflatoxine in produsele depozitate. Data fiind importanta lor pentru inducerea unor stari patologice, multe micotoxine au fost caracterizate chimic si s-au elaborat metode pentru depistarea lor in produsele alimentare.
Intoxicatia cu toxine fungice are o semnificatie deosebita, in contextul comunitatilor care sufera de malnutritie. In timpul tratamentului pentru malnutritie, indivizii afectati sufera leziuni hepatice severe, datorate micotoxinelor. Intoxicatia se produce din cauza ca sursele proteice folosite pentru corectarea malnutritiei (arahide, cereale) pot fi contaminate cu fungi producatori de micotoxine.
Micotoxinele sunt un factor de mediu cu semnificatie majora, care influenteaza starea de sanatate. Dintre produsele naturale, micotoxinele sunt cele care pericliteaza cel mai grav starea de sanatate. OMS atrage atentia asupra pericolelor pentru sanatate, pe care le implica consumul alimentelor contaminate cu fungi.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 8822
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved