| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Mecanismul de actiune al hormonilor
Hormonii isi produc efectele la nivel celular fie prin reglarea functiei nucleare (reglare cantitativa) fie prin modificarea activitatii unor enzime existente in celule (reglare calitativa).
1. Mecanismul de actiune al hormonilor hidrofili
Hormonii hidrosolubili nu pot traversa membrana celulelor tinta si interactioneaza cu receptorii membranari.
Fixarea hormonului pe receptor activeaza un sistem transductor care transforma semnalul extern (mesagerul prim) intr-unul intracelular (mesager secund). Mesagerul secund reprezinta hormonul in celula. Mesagerul secund este un compus chimic ce interactioneaza in interiorul celulei, initiind evenimente care duc la activarea sau inactivarea enzimelor, secretiei, contractiei,
etc. Cu toate ca hormonii sunt intr-un numar mare totusi mesagerii secunzi, intracelulari sunt intr-un numar redus.
Formarea mesagerilor secunzi implica existenta unor sisteme transductoare a mesajelor externe. Pe ansamblu acest sistem mesagerial cuprinde:
- receptorul hormonal
- sistemul de cuplare a complexului hormon-receptor cu sistemul efector
- sistemul efector care genereaza mesageri secunzi
- actiunea mesagerului secund asupra unor proteine tinta, care determina raspunsul biologic.
Sistemele de cuplare sunt reprezentate de proteinele G.
Proteinele G sunt proteine membranare ce cupleza functional receptorii hormonali de sistemul efector generator de mesageri secunzi.


O situatie patologica in care este incriminata proteina G este holera. Toxina holerica este o enzima produsa de vibrionul holeric. Toxina are proprietatea ca anuleaza activitatea GTP-hidrolazica mentinand astfel subunitatea as in stare permanent activa, care genereaza continuu mesager secund (AMPc), activeaza permanent pompa de Na+ din celulele mucoasei intestinale determinand pierderi masive de ioni de Na+ si apa.
Clinic se manifesta prin scaune diareice, colici abdominale si deshidratare.
Netratata este letala. Modul de actiune al toxinei holerice a permis elucidarea mecanismului de actiune al proteinelor G.
Sistemele efectoare generatoare de mesageri secunzi sunt:
-sistemul adenilat ciclazei
-sistemul fosfolipazei C
-sistemul guanilat ciclazei
- sistemul tirozin-kinazei
Sistemul efector al adenilat ciclazei cuprinde: receptorii hormonali, proteinele Gs si Gi, adenilat ciclaza si fosfodiesteraza. Adenilat ciclaza este o enzima situata in membrana celulara a celulelor eucariote, fiind activata de Gs si inhibata de Gi. Sub actiunea adenilat ciclazei din ATP se sintetizeaza AMPc (acid adenozin monofosforic ciclic). Mesagerii secunzi au capacitatea de a modifica activitatea unor proteine intracelulare.
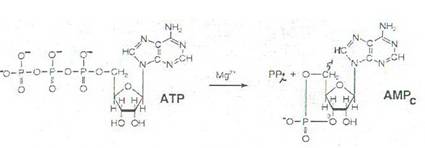
Hormonii care activeaza adenilat ciclaza sunt: glucagonul, ACTH, parathormonul, adrenalina prin receptori P -adrenergici, etc.
Hormonii care inhiba activitatea adenilat ciclazei sunt: catecolaminele prin receptori a2-adrenergici, angiotensina II, somatostatina, opiaceele, etc.
AMPc are o viata scurta deoarece este descompus rapid prin hidroliza la AMP. Fosfodiesteraza este enzima ce catalizeaza aceasta reactie de hidroliza:
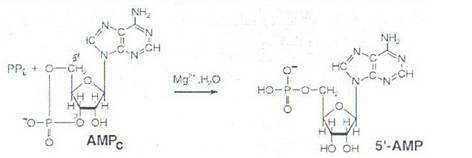
Fosfodiesteraza este activata de: Ca +, prostaglandine, insulina, IGF, etc si este inhibata de hormonii steroizi, hormonii tiroidieni, metilxantine (cafeina, teofilina, etc). Teofilina din ceai si cafeina din cafea ca inhibitori ai fosfodiesterazei, explica efectul stimulator al acestor produsi, in sensul ca pentru AMPc ele mimeaza in celula efectul hormonului prelungind actiunea acestuia.
Durata de viata a AMPc depinde atat de activitatea adenilat ciclazei cat si de cea a fosfodiesterazelor. Rolul AMPc este de a controla exprimarea unor gene, prin legarea sa la o proteina reglatoare (la procariote) sau activeaza protein-kinaze specifice, de exemplu proteinkinaza A (la eucariote)
AMPc-ul a fost descoperit in 1950 de catre Earl Sutherland si a fost primul compus cu rol de mesager secundar.
Hormoni care utilizeaza AMPc ca mesager secund sunt :
|
Hormoni care Hormoni care |
|
Semnal extern Tesut |
Raspuns |
|
|
Glucagon |
Ficat |
Glicogenolizii, gluconeogencza |
|
Catecolamine (prin receptori β) |
Ficat |
Glicogenoliza |
|
Catecolamine (prin receptori α2) Parathormon TSH Gonadotropine (LH, FSH) ACTH |
Tesut adipos Os Tiroida Gonade Corticosuprarenala |
Lipoliza Hipercalcemie Secretie de T4 si T3 Secretie de hormoni sexuali Secretie de cortisol |
Protein kinazele AMPc dependente:
Protein kinazele activate de AMPc sunt cunoscute ca proteinkinaze A (I si II), avand specificitate pentru resturile de serina sau treonina din proteina ce o regleaza. Proteinkinaza A este un
reglatoare (R)
Tetramerul R2C2 este inactiv, centrul catalitic fiind mascat de subunitatea R. Subunitatea reglatoare prin legarea a doua molecule de AMPc sufera o tranzitie alosterica si se disociaza din complex:

Desi diversi hormoni determina formarea aceluiasi mesager secund, totusi raspunsul celular poate fi mai mult sau mai putin specific, depinzand de natura proteinelor care sunt substrat al proteinkinazei A.
De exemplu, prin fosforilare se mareste sau se micsoreaza activitatea catalitica a unei enzime. Daca proteina fosforilata este es insasi o kinaza are loc o amplificare in cascada a semnalului initial. Fosforilarea poate modifica concentratia unor metaboliti reglatori (exemplu: fructozo 2,6 bisfosfat, reglator al glicolizei si gluconeogenezei).
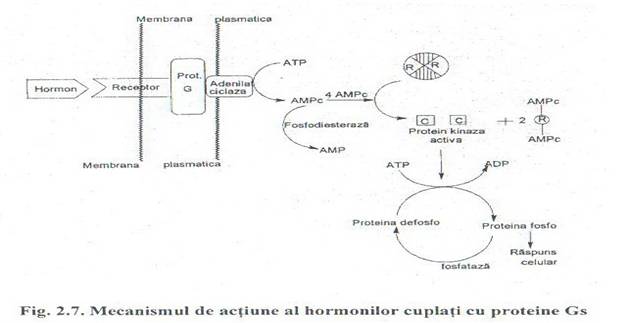
Fig. 2.7. Mecanismul de actiune al hormonilor cuplati cu proteine Gs
Fosfoproteinfosfatazele
Aceste enzime catalizeaza defosforilarea proteinelor fosforilate de catre proteinkinaza A. Prin aceasta transformare se stinge semnalul hormonal, proteinele revenind la forma anterioara receptarii mesajului hormonal.
Proteina-O-(P) +H2 O - -> Protema-OH + Pi
Stingerea semnalului hormonal care determina cresterea AMPc se poate realiza si pe alte cai, astfel:
autohidroliza GTP din ocs-GTP si inactivarea proteinei G
interventia fosfodiesterazei care hidrolizeaza AMPc la AMP
Ciclul fosforilare-defosforilare a proteinelor, intervine si in procesele de diviziune celulara, transcriere si traducerea unor gene, etc
Sistemul efector al fosfolipazei C
Acest sistem efector este alcatuit din:
-receptori hormonali,
-proteine Gp,
-fosfolipaza C
- si proteinkinaza C.
Fosfolipaza C (PLC), este o enziina din membrana plasmatica care catalizeaza reactia de hidroliza a fosfatidil inozitol 4,5-bisfosfat (PIP2) ce este un fosfolipid membranar:
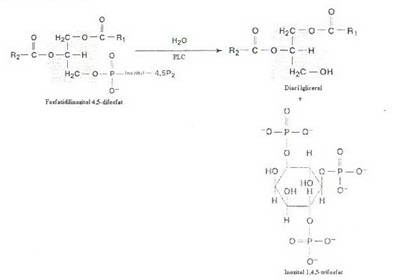
Cei doi produsi de reactie IP} si DAG actioneaza ca mesageri secunzi pentru hormonii ai caror receptori sunt cuplati cu proteinele Gp.
IP3 este la pH-ul fiziologic un polianion, puternic hidrofil, difuzeaza in citosol unde prin deschiderea unor canale ionice determina eliberarea de Ca2+ din reticulul endoplasmatic in citosol.
Diacilglicerolul (DAG) este un compus liposolubil, ramane ancorat de membrana plasmatica de unde activeaza proteinkinaza C.
Hormonii ce au ca mesageri secunzi IP3, DAG, Ca2+ sunt: catecolaminele prin receptori oti-adrcncrgici, acetilcolina prin receptori muscarinici, gastrine, colecistokinina, hormonul eliberator al gonadotropinelor, etc.
Proteinkinaza C este o proteina ce se gaseste in membrana plasmaticA. Prezinta mai multe izoforme. Unele sunt activate numai de DAG, altele necesita pentru activare si ioni de Ca2+. Are specificitate pentru resturile de serina sau treonina, dar diferite de cele fosforilate de proteinkinaza A.
Ca2+ mediaza efectele intracelulare ale multor hormoni sau a unor semnale nervoase. Concentratia citosolica este foarte mica, aproximativ l 0-7- l O -6M, iar concentratia calciului extracelular este de 10 -3 M.
Concentratia calciului intracelular este mentinuta mica prin functionarea unor pompe de Ca2+, care scot calciul din celula. Deasemenea in aceelasi scop functioneaza mecanisme ce depoziteaza Ca2+ in mitocondrii si in reticulul endoplasmic si nu trebuie neglijat faptul ca in mod natural membranele biologice au permeabilitate foarte mica pentru cationi. Rolurile reglatoare ale Ca2+sunt mediate de proteine specifice.
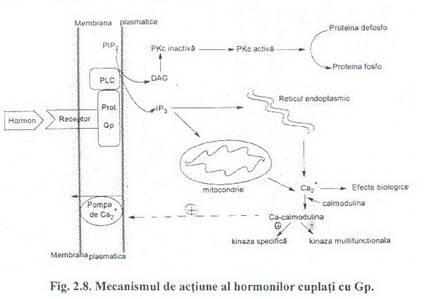
Fig. 2.8. Mecanismul de actiune al hormonilor cuplati cu Gp.
Calmodulina, este o proteina mica ce contine 148 aminoacizi, fiind prezenta in toate celulele eucariote. Poate fixa 4Ca2+/molecula. Fixarea celor 4Ca2+ de catre calmodulina determina trecerea acesteia intr-o conformatie mai compacta, activa, capabila sa interactioneze cu o alta proteina in care induce o tranzitie conformationala si astfel se modifica functia proteinei.
Complexul Ca2+-calmodulina regleaza: contractia musculaturii netede, activeaza pompa de Ca2+ din membrana plasmatica, regleaza activitatea unor proteinkinaze, interfera in celelalte sisteme mesageriale prin reglarea activitatii adenilat ciclazei, guanilat ciclazei, fosfodiesterazelor, etc.
Sistemul efector al guanilat ciclazei, este alcatuit din guanilat ciclaza, proteinkinaze -GMPc dependente. Guanilat ciclaza este o enzima ce prezinta doua izoforme: una membranara si alta citoplasmatica.
Guanilat ciclaza membranara, are atat functie de receptor (domeniu cxtracelular) cat si functie de sitem efector ce genereaza mesagerul secund GMPc.
Factorul atrial natriuretic (un hormon ce contine 28 de aminoacizi) este secretat de celulele cardiace atriale la diverse semnale si actioneaza la nivelul rinichiului si a zonei glomerulare a medulosuprarenalelor. Raspunsul celular ca urmare a actiunii hormonale este reprezentat de cresterea volumului urinar, cresterea eliminarii de Na+, scaderea secretiei de renina si aldosteron.
Guanilat ciclaza citosolica , este solubila si reprezinta sistemul efector pentru monoxidul de azot (NO). NO a fost identificat cu factorul de relaxare derivat din endoteliu (endothelium derived relaxing factor, EDRF). Acesta este hormon paracrin si actioneaza asupra multor celule, fiind un factor reglator al presiunii sanguine, mediaza citotoxicitatea si este implicat in neurotransmisie.
Monoxidul de azot se sintetizeaza de catre un sistem enzimatic cu functie oxidazica (NO-sintetaza), folosind ca precursor arginina. Monoxidul de azot activeaza in celulele tinta guanilat ciclaza citoplasmatica cu formare de GMPc, care mediaza relaxarea muschilor netezi.
GMPc, este sintetizat prin reactia catalizata de guanilat ciclaza din GTP si este descompus prin reactia catalizata de GMPc-fosfodiesteraza. Efectele GMPc in multe cazuri antagonizeaza cu cele produse de AMPc.
GMPc este mediatorul semnalelor luminoase in celulele retiniene (cu cornuri si bastonase). Semnalul luminos receptat de rodopsina prin intermediul unei proteine din familia G (transducina) determina activarea GMPc-fosfodiesterazei si are loc scaderea concentratiei intracelulare de GMPc. in urma acestui proces are loc inchiderea canalelor pentru Na+, creste concentratia acestora intracelulara si are loc o hiperpolarizare a membranei care se va constitui in senzatia de lumina.
2. Mecanismul de actiune al hormonilor hidrofobi
Receptorii acestor hormoni sunt intracelulari, deci hormonii hidrofobi patrund in celula tinta prin difuzie si in citoplasma sau nucleu isi gasesc receptorii specifici cu care formeaza complexul hormon-receptor.
Legarea hormonului la receptor modifica conformatia receptorului, facandu-1 apt sa interactioneze cu o portiune din cromatina nucleara, in nucleu complexul hormon-receptor este recunoscut si legat specific de o anumita portiune a ADN-ului, numita element de raspuns hormonal.
Interactiunea hormon-receptor-element de raspuns hormonal are repercursiuni asupra promotorului care va modula initierea transcrierii unor gene specifice, fie stimuland initierea transcrierii, fie blocand-o. Astfel, daca initierea transcrierii a fost favorizata, gene specifice din ADN vor fi transcrise sub forma de ARNm care eliberati in citoplasma vor genera prin traducere, la nivelul ribozomilor, proteine cu functii specifice.
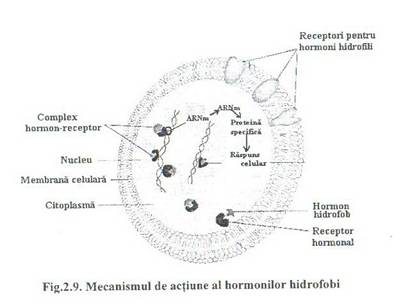
Fig.2.9. Mecanismul de actiune al hormonilor hidrofobi
Dupa acest mecanism actioneaza hormonii steroizi si tiroidieni.
Complexul hormon-receptor dupa interactia cu receptorul si obtinerea raspunsului celular, sufera un proces de disociere, hormonul trece in sange de unde este luat de ficat si catabolizat, iar receptorul se recicleaza.
Prin actiunea la nivelul membranei celulare hormonii tiroidieni stimuleaza patrunderea aminoacizilor cu radical hidrofob si a glucozei in celule.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 6866
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved