| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Neuronul - o celula secretorie
1. Neuronii si produsii de secretie
Neuronul sintetizeaza diferiti neurotransmitatori si neuromodulatori. El este astfel o celula specializata cu functie secretorie. Membrana secretorie (membrana presinaptica) poate fi la o distanta de peste 1 m de locul unde are loc secretia. Conexiunea dintre doua regiuni este realizata desigur de catre axon. Axonul trebuie sa asigure pasajul produsilor de secretie de la nucleu catre butonul sinaptic, unde vor fi eliberati. Mai mult, transportul nu se realizeaza intr-o singura directie.
Cativa neuroni si-au dezvoltat functia de secretie nu pentru a o elibera in fanta finaptica, ci direct in sistemul vascular. Aceste celule sunt cunoscute ca celule neurosecretorii (fig. 2-1). Asemenea celule sunt intalnite in hipotalamus si glanda pituitara. In momentul de fata, exista un interes major in difuzia neurotransmitatorilor, in special a particulelor mici in spatiul intercelular.
Sinteza in corpul celular
Mesajul genetic pastrat in structura ADN-ului este transcris in ARNm si apoi translatat in robozomi in secventele specifice de aminoacizi (AA) ce alcatuiesc proteinele. Acest proces este foarte complex. El implica multe tipuri de enzime si acizii ribonucleici ARNt, ARNr, si ARNm. In celulele secretorii, proteina trebuie sa fie impachetata si transportata pana la locul unde este eliberata.
Corpul celular al celulelor nervoase implicate in biosinteza proteinelor prezinta un nucleu mare cu unul/mai multi nucleoli. Citoplasma este bogata in RE rugos si prezinta intotdeauna un aparat Golgi (fig. 2-2).
1. Insertia co-translationala
Lantul polipeptidic elaborat de catre ribozomi este inserat direct in lumenul RE. Insa nu toate proteinele pe care le fabrica un neuron sunt destinate exportului. Cateva dintre ele sunt pastrate pentru activitati "gospodaresti" ale nucleului.
Primii 30 AA din lantul polipeptidic ce se formeaza din ribozom, alcatuiesc o secventa semnal ce este recunoscuta de catre o "particula de recunoastere a semnalului" (signal recognition particle - SRP) (fig. 2-3). Aceasta particula prezinta doua functii. In primul rand previne orice translatie ulterioara, asigurand celula ca proteina de export nu este pur si simplu abandonata in citoplasma. In al doilea rand, secventa semnal este recunoscuta de un "receptor al particulei de recunoastere al semnalului" sau "docking protein" (DP) din membrana RE.
S-a demonstrat ca daca nucleoproteina SRP ramane atasata la secventa semnal, translatia nu mai are loc. Totusi, in mod normal, odata ce ribozomul se ancoreaza la membrana, SRP este inlaturat pentru a fi reciclat. Acest lucru permite ca secventele semnal de AA, ce sunt in general hidrofobi, sa se ataseze la un complex proteic mare al membranei RE cunoscut drept translocon. Transloconul are un por central hidrofobic care este de obicei inchis, dar care este deschis de secventa semnal. Ribozomul este pozitionat deasupra porului de catre lantul polipeptidic, in timp ce capatul sau N-terminal patrunde prin por in lumenul RE. Polipeptidul este astfel sintetizat si inserat in lumenul RE in acelasi timp. Nu este expus deloc in citosol si nu se impacheteaza decat atunci cand este in RE. Intregul proces este denumit insertie co-translationala. In final, secventa semnal N-terminala este taiata de catre enzime cand intalneste un semnal de oprire (UAA) pe ARNm, apoi se disociaza si este reciclata (fig. 2-4).
Exista 2 modalitati posibile de terminare a acestui proces de co-translatie (fig. 2-5). Intr-unul din cazuri, polipeptidul este secretat complet in lumenul RE. Acesta este cazul proteinelor destinate exportului - neurotransmitatori. In al 2-lea caz, polipeptidul nu patrunde deloc in lumenul RE. Acesta este cazul proteinelor care intra in alcatuirea membranei plasmatice (neurilema).
Aparatul Golgi si modificarile post-translationale
Primii pasi in procesul post-translational au loc in lumenul RE. Aici apar legaturi disulfurice intre rezidurile de cisteina si este initiata glicozilarea.
Pasii initiali ai glicozilarii se numesc glicozilare centrala (core glycosylation) pentru a o diferi de modificarile de final din aparatul Golgi (fig. 2-6). Aceste etape implica frecvent adaugarea unei oligozaharide catre capatul aminic al asparaginazei. Aceasta oligozaharida prezinta intotdeauna 2 unitati de monozaharide acetilate - N-acetilglucozamina (NAG) si un numar de unitati de manoza si glucoza.
Etapa urmatoare a procesului post-translational are loc in aparatul Golgi. Celulele active in sinteza de proteine pentru export au de obicei un numar mare de complexe Golgi. Acest proces are loc si in cazul multor neuroni. Figura 2-7 prezinta structura complexa a aparatului Golgi. El prezinta numeroase cisterne sau saci membranosi. Acesti saci se formeaza la nivelul uneia din fete - cis, prin fuziunea veziculelor ce inmuguresc din RE si apoi se desprind de la nivelul celeilalte fete - trans, pentru a constitui veziculele secretorii. Astfel se poate spune ca aparatul Golgi prezinta o fata cis (de formare) si o fata trans (de maturare).
Veziculele de transport si cele secretorii prezinta un element comun. Ele sunt inconjurate de molecule de clatrina. Fiecare molecula prezinta o structura cu 3 brate = trischelion (160kDa)(fig. 2-8). Un anumit numar de trischelioni se aduna in jurul unei membrane lipoproteice a unei vezicule si cu ajutorul catorva proteine mici mentin vezicula "invelita". Clatrina se poate asambla si dezasambla foarte usor.
Odata ce veziculele de transport provenite din RE au fuzionat cu fata cis a aparatului Golgi, proteinele pe care le contin sufera mai departe procesul co-translational (fig. 2-6). Sacii aparatului Golgi contin multe enzime ce continua procesul de glicozilare = glicozilare terminala. Alte enzime adauga un semnal, probabil prin fosforilare sau printr-o glicozilare speciala, ce serveste proteinei pentru a ajunge la destinatia corecta. In final, aparatul Golgi are rolul important de a impacheta proteinele destinate pentru export.
Astfel, proteinele sunt sintetizate, impachetate si pregatite pentru export (fig. 2-9).
3. Transportul de-a lungul axonului
Veziculele secretorii ce pleaca de pe fata trans a aparatului Golgi trebuie sa se deplaseze in continuare.
Exista 2 sisteme de transport distincte: unul functioneaza rapid pentru a transporta produsii de secretie de la corpul celular catre capatul sinaptic (anterograd), iar celalalt sistem functioneaza lent in directie inversa (retrograd).
Transportul rapid axoplasmic este responsabil de deplasarea mitocondriilor si a veziculelor secretorii. Aceste vezicule contin neurotransmitatori si modulatori, glicoproteine si enzime necesare metabolismului. Sistemul lent este implicat in transportul elementelor citoscheletului (tubulina, actina, neurofilamente). Sistemul retrograd transporta materiale uzate, factori neurotrofici (NGF).
Exista 3 tipuri majore de fibre citoscheletice - microfilamentele (cu un diametru de 8nm), filamentele intermediare (Ifs) (7-11nm) si microtubulii (25nm). Toate aceste 3 tipuri de fibre sunt intalnite in neuroni, in special in axon.
3.1. Microfilamentele
Sunt alcatuite in principal din actina. Actina este desigur intalnita si in muschi, unde este implicata in mecanismul de contractie. Fibrele de actina (F-actina) sunt alcatuite din subunitati globulare (G-actina). Doua subunitati de F-actina sunt rasucite una in jurul celeilalte pentru a forma o "franghie" (fig. 2-10).
3. Filamentele intermediare
Spre deosebire de microfilamente sau microtubuli, filamentele intermediare sunt d.p.d.v biochimic extrem de heterogene. Exista 5 clase majore de filamente intermediare, fiecare clasa fiind intalnita intr-un anumit tip de celula. In sistemul nervos, celulele gliale contin filamente gliale (o singura proteina cu 51kDa), in timp ce neuronii prezinta NF-L (63kDa), NF-M (169kDa) si NF-H (200kDa) (cu greutate moleculara joasa, medie si inalta).
Chiar daca filamentele intermediare sunt extrem de variate, ele prezinta un domeniu central de 310 AA (fig. 2-11). Acest domeniu prezinta 2 α-helixuri rasucite unul in jurul celuilalt. Catre fiecare capat al domeniului exista 2 regiuni hipervariabile - "capul" si "coada".
3.3. Microtubulii (MTs)
Microtubulii sunt alcatuiti din tubulina. Precum actina, tubulina prezinta subunitati globulare. Insa subunitatile tubulinice nu se aseamana intre ele si sunt de doua feluri: α si β tubulina; fiecare subunitate (monomer) prezinta 3 domenii. Subunitatile α si β se asociaza pentru a forma dimerii si acestia se unesc cap la cap pentru a forma protofilamentul. In final 13 protofilamente se aliniaza paralel si in cerc pentru a forma un tub. (fig. 2-12). Tubul este polarizat: prezinta un cap (+) si o coada (-). Polimerizarea are loc prin adaugarea de noi subunitati la capatul (+). In axon, microtubulii sunt aliniati astfel incat capatul (-) este indreptat catre corpul celular. In dendrite totusi, sunt aranjati in ambele directii.
Polimerizarea necesita un anumit numar de factori accesori si GTP. Doi din cei mai importanti factori sunt proteinele accesorii microtubulului (MAPs) si proteinele tau. Acestea sunt implicate atat in procesul de polimerizare, cat si in stabilizarea tubului.
3.4. Citoscheletul axonal
Sistemele in vitro au permis izolarea si analiza a doua clase de proteine motorii: kinezinele si dineinele citoplasmatice.
Kinezina
Kinezinele (KIFs) sunt de asemenea prezente si in alte celule, unde sunt implicate in transportul veziculelor si materialelor de la o celula la alta si in formarea fusului in timpul diviziunii celulare. Ele prezinta 2 domenii globulare mari ce au activitate ATP-azica, conectate la un complex de transport. (fig. 2-13A). Kinezinele sunt proteine motorii asociate microtubulilor: KIF1A, KIF1B, KIF2, etc.
Dineina citoplasmatica
Dineinele citoplasmatice sunt strans legate de dineinele din cili si flageli. Ca si kinezinele, ele sunt implicate in formarea de fusului in timpul diviziunii celulare si de asemenea prezinta doua domenii globulare mari. Ele sunt totusi molecule foarte mari si mai putin variate. Cele doua domenii globulare contin fiecare 4 situsuri de legare a ATP si furnizeaza energia pentru miscarea de-a lungul microtubulilor.
Transportul
Dineinele citoplasmatice si kinezinele sunt implicate in transport in cadrul celulelor eukariote, precum si in miscarea din timpul diviziunii celulare mitotice si meiotice. Ambele proteine motorii sunt ATP-aze dependente de microtubuli, activitatea lor fiind localizata la nivelul celor 2 capete globulare (fig. 2-13). Capetele lor se ataseaza la un microtubul, iar coada lor se ataseaza la veziculele translocate.
Veziculele imbracate in kinezina si dineina citoplasmatica se misca de-a lungul microtubulului urmand niste linii de ghidare. S-a demonstrat ca veziculele invelite in kinezina se misca catre capatul (+) al microtubulului (anterograd), cele invelite in dineina citoplasmatica se misca catre capatul (-) (retrograd) (fig. 2-14).
Atasarea veziculelor
Capatul complexului kinezina-dineina citoplasmatica este de o importanta deosebita. Deoarece cele 2 proteine motorii se misca in directii opuse de-a lungul microtubulului, capatul complexului trebuie sa se ataseze de incarcatura (cargo) potrivita. Intradevar, dineina citoplasmatica trebuie sa fie transportata mai intai intr-o forma inactiva de catre kinezina catre capatul terminal al axonului si apoi sa isi preia incarcatura de material folosit (uzat), si NGF pentru transportul catre corpul celular.
Au fost identificate 2 proteine ce fac legatura intre elementele membranare si lanturile usoare de kinezina. Acestea sunt denumite intr-un mod hazliu "conducatorul de duminica" ("Sunday driver")(Syd) si proteina precursorului amiloid (APP). Ambele proteine au un rol important in atasarea cargoului impachetat inauntrul veziculelor membranoase la kinezina.
Dineina citoplasmatica este o molecula mult mai mare si se presupune ca isi ataseaza incarcatura printr-un numar de legaturi polipeptidice in lanturile ei usoare.
4. Exocitoza si endocitoza la capatul sinaptic
Dupa zile sau chiar saptamani, veziculele sinaptice ajung in sfarsit la destinatia lor finala - butonul sinaptic. Acum asteapta scurtul lor moment de actiune. Acesta depinde de potentialul de actiune. Cand acesta are loc, depolarizarea membranei deschide canalele de Ca2+. Influxul de Ca2+ declanseaza eliberarea continutului veziculei in fanta sinaptica.
Examinarea terminatiilor sinaptice la ME (microscopul electronic) arata ca ele contin numeroase vezicule sinaptice. Aceste vezicule variaza in dimensiuni si forma, in functie de continutul lor. Veziculele mici, sferice contin acetilcolina, glutamat, etc. Alte vezicule mici au o forma elipsoidala si contin neurotransmitatori inhibitori cum ar fi glicina. Veziculele mai mari contin catecolamine, in timp ce veziculele si mai mari contin peptide.
In figura 2-15 este prezentata o jonctiune neuromusculara. Aceasta foloseste acetilcolina drept neurotransmitator.
Fig. 2-15C arata o imagine de ME a unei jonctiuni neuromusculare de broasca. Se observa foarte usor veziculele. Fiecare dintre ele contin 5000-10000 molecule de acetilcolina; la nivelul terminatiei presinaptice pot exista mai mult de 1 milion de vezucule. Axoplasma din spatele veziculelor contine acumulari dense de elemente citoscheletice. Acest lucru este denumit reteaua presinaptica.
Veziculele se ciocnesc continuu de membrana presinaptica si isi elibereaza continutul in fanta sinaptica. Aceste pachete de 5000-10000 molecule de acetilcolina reprezinta un "cuantum". Ele cauzeaza o depolarizare mica in membrana subsinaptica. Totusi majoritatea veziculelor fuzioneaza cu membrana presinaptica si isi elibereaza continutul doar atunci cand terminalul presinaptic este depolarizat de un potential de actiune.
In figura 2-16 se prezinta membrana presinaptica a unei jonctiuni neuromusculere pe sectiuni inghetate.
4.1. Adunarea veziculelor
Am vazut mai devreme ca atunci cand veziculele au fost impachetate in pericarion, ele au fost apoi invelite de catre clatrina. Se pare ca anumite vezicule sunt atasate de bratele kinezinei prin intermediul clatrinei. Clatrina nu este desigur specifica neuronului, ea se intalneste in majoritatea celulelor secretorii. O alta proteina este totusi specifica neuronului. Aceasta este synapsina 1. Aceasta proteina formeaza un invelis proteic mai ales in cazul veziculelor mici (fig. 2-17).
4. Ca2+
Calciul liber este in cantitati foarte mici in celule. Astfel, orice influx de Ca2+
din exterior poate avea un efect dramatic. Unul din aceste efecte este activarea catorva kinaze. Doua dintre aceste kinaze dependente de Ca2+ - protein kinaza A (PKA) si protein kinaza 2 dependenta de Ca2+- calmodulina, fosforileaza synapsina 1 (fig. 2-18).
Secventa reactiilor biochimice determina influxul de Ca2+ in butonul presinaptic, ceea ce duce la eliberarea veziculelor cu synapsina.
4.3. Andocarea veziculelor
Proteine de fuziune specifice sunt necesare pentru a uni membrana veziculelor sinaptice cu membrana presinaptica.
Cateva tipuri de proteine prezinta un rol destul de complicat, insa coordonat. Cele mai evidente sunt proteinele SNARE.
La inceput acestea au fost clasificate in 2 grupe: vSNARE asociate cu veziculele de secretie, si tSNARE asociate cu membranele tinta.
Pe langa proteinele SNARE, si alte proteine joaca roluri importante in cadrul sinapsei. Aici putem include sinaptotagminele si sinaptofizinele din membrana veziculei; neurexina si canalele de Ca, de pe fata P a membranei presinaptice. In plus, cateva proteine non-membranare joaca roluri esentiale: n-sec1, rab3A, α-SNAP. Acest complex este prezentat in figura 2-18B.
Nu trebuie uitat ca semnalul pentru asamblarea tuturor factorilor este sosirea potentialului de actiune si in consecinta deschiderea canalelor de Ca. Exista dovezi ca sinaptotagmina functioneaza ca un senzor al Ca2+.
Sinaptofizina, dupa cum se observa in figura 2-18A, este o proteina cu 4 pasaje transmembranare (4TM) si poate forma un por ionic. Cand se formeaza complexul de fuziune (fig. 2-18B), porul sinaptofizinei se poate alinia cu canalele gemene (fizofilina) in membrana presinaptica.
Proteinele vSNARE si tSNARE sunt foarte importante pentru fuziunea veziculelor cu membrana presinaptica. Ele formeaza un complex de legatura. Proteinele non-membranare sunt de asemenea esentiale, de exemplu Rab3A este un membru al superfamiliei ras a GTP-azelor care joaca un rol important in pregatirea fuziunii veziculelor cu membrana presinaptica. Rab3A se ataseaza la complexul de legatura si daca ramane stabil o anumita perioada de timp pentru a hidroliza GTP-ul, blocheaza complexul in aceasta pozitie.
NSF si α-SNAP sunt necesare pentru a finaliza procesul de fuziune. N-sec1 este necesara in etapele initiale ale reactiei. Disocierea ei de sintaxina permite sinaptobrevinei (o proteina vSNARE) sa se lege de sintaxina si SNAP25 (tSNARE).
4.4. Eliberarea neurotransmitatorilor
Daca in loc de jonctiunea neuromusculara am analiza o sinapsa tipica, am observa ca veziculele sunt ordonate de catre o retea presinaptica. Aceasta retea este formata de proiectii dense care apar din membrana presinaptica si se indreapta catre interiorul sinapsei (fig. 2-19). Aceste proiectii sunt legate impreuna de filamente fine si formeaza o retea ordonata pe fata P a membranei presinaptice. Desenul pare sa fie hexagonal. Sase spatii triunghiulare inconjoara fiecare proiectie si acestea sunt probabil ariile membranare specializate pentru atasarea veziculelor sinaptice.
Deoarece eliberarea neurotransmitatorului din veziculele mici este foarte rapida (aproximativ 200s dupa influxul de Ca) se poate spune ca veziculele sunt deja in contact cu membrana (langa canalele de Ca), cand soseste potentialul de actiune; astfel nu ar fi timp suficient pentru ca vezicule sa se mute din locurile de atasare la citoschelet.
Se poate spune in concluzie ca exista doua populatii de vezicule mici: un grup ce se poate elibera pentru ca este deja in contact cu fata P a membranei presinaptice si un grup de rezerva atasat prin sinapsina la citoschelet.
Eliberarea continutului veziculei se realizeaza prin exocitoza.
4.5. Disocierea complexului de fuziune si recuperarea si reconstructia membranei veziculare
Se stie ca NSF si α-SNAP sunt necesare pentru a forma complexul SNARE. Acest complex se formeaza intre 2 membrane diferite (veziculare si presinaptice). Acesta este denumit complexul trans. Dar odata ce membranele au fuzionat, complexul SNARE este prezent intr-o singura membrana, in pozitia cis. In aceasta pozitie, NSF si α-SNAP in loc sa asambleze complexul, il dezasambleaza. Proteinele complexului sunt eliberate si sunt gata pentru a forma un alt complex ca raspuns la un nou influx de Ca2+.
De indata ce vezicula este indepartata de membrana presinaptica, se poate recicla (fig. 2-20B) sau poate fuziona cu un endozom din cadrul terminatiei presinaptice (fig. 2-20A). Aici componentele membranei sunt reasamblate si noi vezicule apar prin inmugurire. Mai exista o ipoteza - "kiss and go" - in care vezicula nu isi pierde niciodata integritatea si este umpluta cu neurotransmitatori si apoi refolosita (fig. 2-20C).
4.6. Realimentarea veziculelor
Daca vezicula contine neurotransmitatori mici, ea este umpluta chiar din aceasta regiune terminala. In alte cazuri, cand neurotransmitatorul nu este sintetizat aici, ci in corpul celular, vezicula este transportata de-a lungul axonului prin intermediul sistemului retrograd. Aceste vezicule de obicei fuzioneaza pentru a forma un corp multivezicular.
Acest mecanism este exemplificat in figura 2-21.
4.7. Finalizarea eliberarii neurotransmitatorilor
Eliberarea lor se incheie prin inlaturarea ionilor de Ca. Exista mai multe mecanisme de inlaturare a acestora: sechestrarea in mitocondrii, in cisternele RE si probabil cel mai important prin calmodulina.
De indata ce Ca2+ este indepartat, semnalul de eliberare a transmitatorilor dispare. Potentialul sinaptic revine la starea lui initiala.
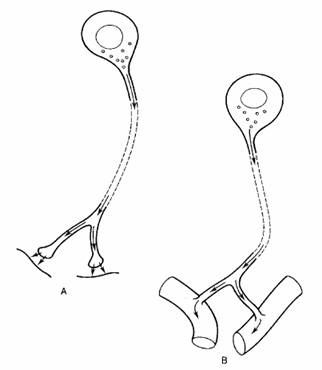
Fig. 2-1 : Neuronul - celula secretorie - (A) Neuron tipic; transmitatorul este eliberat in fanta sinaptica; (B) Celula tipica neurosecretorie; neurosecretie este eliminata intr-un vas sanguin.
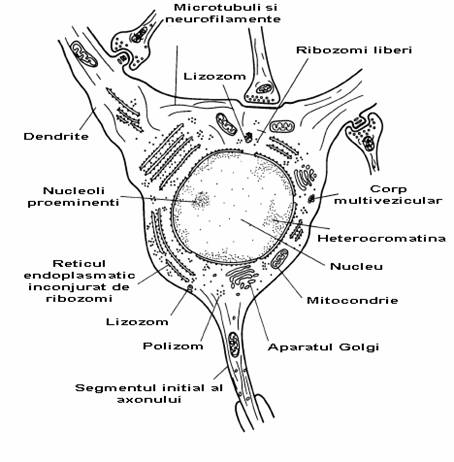 Fig. 2-2 : Corpul celular al unui neuron cortical (ME).
Fig. 2-2 : Corpul celular al unui neuron cortical (ME).
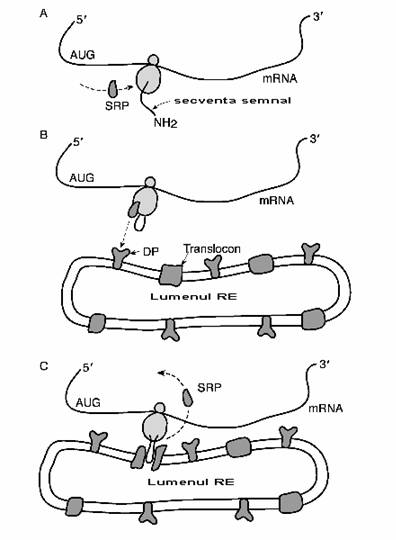
Fig. 2-3 : Ancorarea ribozomului la reticulul endoplasmatic (RE) - (A) Ribozomul se ataseaza la capatul 3' al ARNm si atunci cand intalneste secventa de start AUG incepe translatia; secventa initiala de AA se numeste secventa semnal; (B) Secventa semnal este recunoscuta de o proteina de recunoastere a semnalului (SRP) ce se ataseaza la ea si la ribozom; (C) SRP este recunoscut de catre o proteina ("docking protein")(DP) din membrana RE; translocatia continua la nivelul ribozomului si lantul polipeptidic trece in lumenul RE.
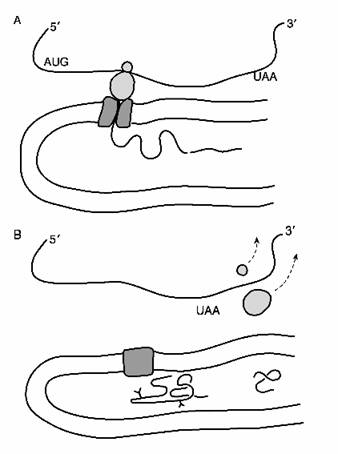 Fig. 2-4 : Insertia
co-translationala a polipeptidului in RE - (A) Lantul polipeptidic este
sintetizat la nivelul ribozomului si inserat in lumenul RE si apoi secventa
semnal este inlaturata. (B) Cand se ajunge la secventa stop (UAA) tranlsatia
inceteaza si cele 2 parti ale ribozomului se separa pentru a fi reciclate.
Fig. 2-4 : Insertia
co-translationala a polipeptidului in RE - (A) Lantul polipeptidic este
sintetizat la nivelul ribozomului si inserat in lumenul RE si apoi secventa
semnal este inlaturata. (B) Cand se ajunge la secventa stop (UAA) tranlsatia
inceteaza si cele 2 parti ale ribozomului se separa pentru a fi reciclate.
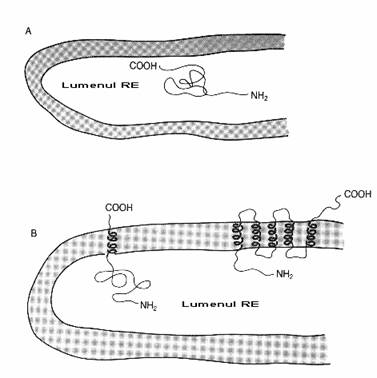
Fig. 2-5 : Doua rezultate ale procesului de co-translatie; (A) - polipeptidul este in intregime in lumenul RE; (B) - segmente hidrofobice ale polipeptidului asigura prinderea sa in membrana RE.
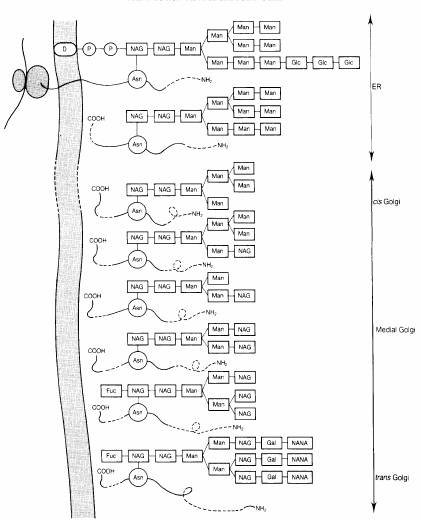 Fig. 2-6 : Glicozilarea
in RE si aparatul Golgi - N-glicozilarea; pasul initial este transferul
unui oligozaharid catre rezidul de asparaginaza ;
Fig. 2-6 : Glicozilarea
in RE si aparatul Golgi - N-glicozilarea; pasul initial este transferul
unui oligozaharid catre rezidul de asparaginaza ;
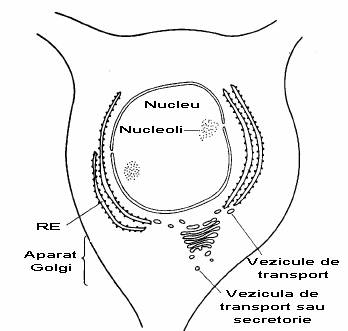 Fig. 2-7 : Structura
aparatului Golgi - Corpul celular al neuronilor contine numeroase aparate
Golgi. Fluxul este de la RE prin veziculele de transport catre fata cis a
aparatului Golgi; veziculele fetei cis formeaza o sacula si se muta catre fata
trans; in final veziculele inmuguresc si se elimina de pe aceasta fata.
Fig. 2-7 : Structura
aparatului Golgi - Corpul celular al neuronilor contine numeroase aparate
Golgi. Fluxul este de la RE prin veziculele de transport catre fata cis a
aparatului Golgi; veziculele fetei cis formeaza o sacula si se muta catre fata
trans; in final veziculele inmuguresc si se elimina de pe aceasta fata.
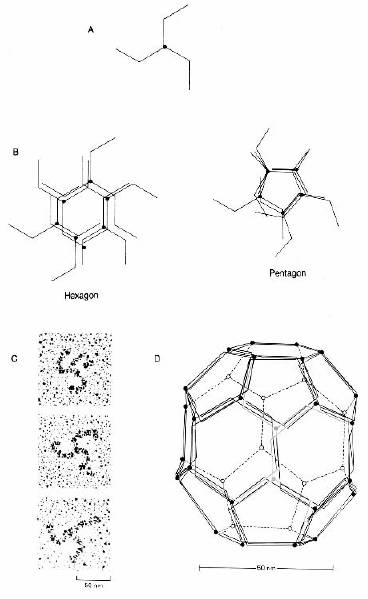 Fig. 2-8 : Trischelionul
- (A) trischelion de clatrina; (B) trischelioanele de clatrina se asambleaza
spontan pentru a forma hexagoane si pentagoane; (C) Imagine la ME; (D) 36
trischelioni organizati intr-o retea de 12 pentagoane si 8 hexagoane formeaza o
"patura" ce inconjoara veziculele.
Fig. 2-8 : Trischelionul
- (A) trischelion de clatrina; (B) trischelioanele de clatrina se asambleaza
spontan pentru a forma hexagoane si pentagoane; (C) Imagine la ME; (D) 36
trischelioni organizati intr-o retea de 12 pentagoane si 8 hexagoane formeaza o
"patura" ce inconjoara veziculele.
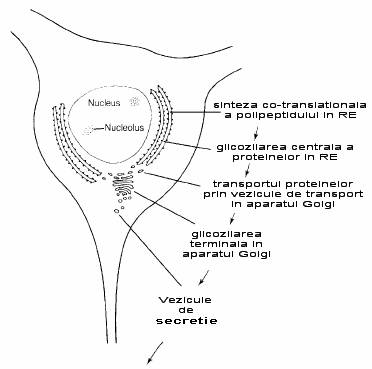
Fig. 2-9 : Diagrama caii de export a proteinelor.
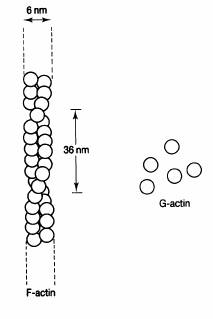 Fig. 2-10 : Structura microfilamentelor.
Fig. 2-10 : Structura microfilamentelor.
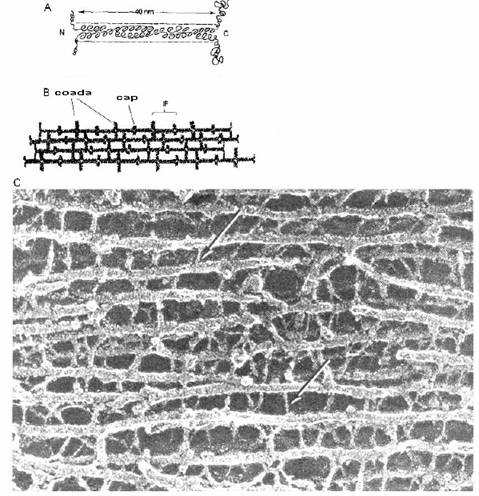
Fig. 2-11 : Structura filamentelor intermediare si a neurofilamentelor - (A) IF - 2 α-helixuri rasucite unul in jurul celuilalt; fiecare segment se termina intr-o regiune variabila; (B) capetele si cozile variabile tind sa se uneasca, formand un fascicul; (C) neurofilamente observate la ME (sagetile indica neurofilamentele).
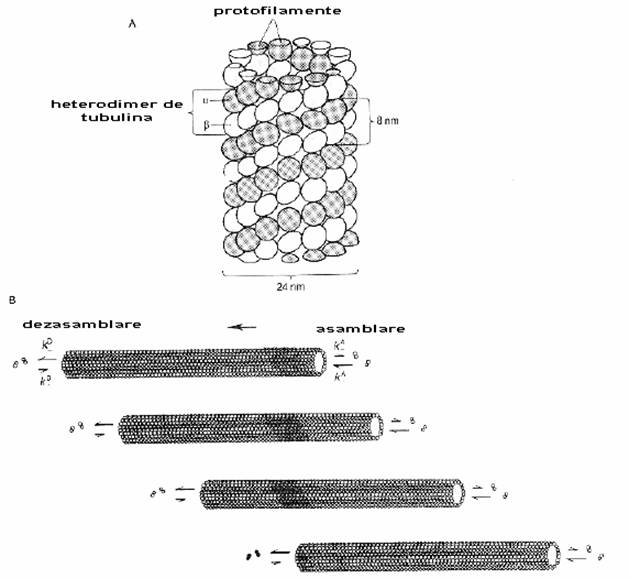
Fig. 2-12 : Structura moleculara a tubulinei - (A) structura moleculara a microtubulilor; tubul prezinta 13 randuri longitudinale de unitati α si β; aceste randuri sunt denumite protofilamente; (B) in prezenta GTP, heterodimeri α si β sunt adaugati la capatul (+) si sunt inlaturati la cel (-); incorporarea unui radioizotop ne permite vizualizarea unei pete negre ce se misca de-a lungul microtubulului.
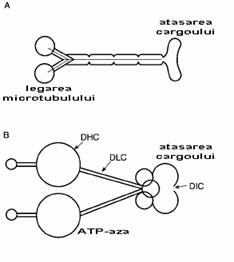
Fig. 2-13 : Structura kinezinei si dineinei citoplasmatice (A) kinezina; (B) dineina citoplasmatica - este o molecula mult mai mare decat kinezina cu o structura mult mai complexa; DHC=dynein heavy chain; DIC=dynein intermediate chain; DLC=dynein light chain.
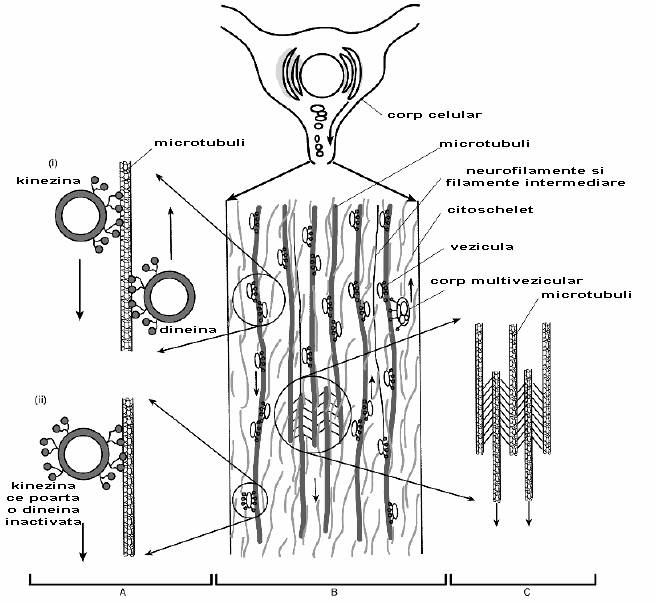
Fig. 2-14 : Transportul axoplasmic (schematic) - (A) (i) veziculele invelite in kinezina si dineina se misca de-a lungul microtubulilor in directie anterograda si retrograda; (ii) kinezina ce porta o dineina inactivata; (B) Diagrama schematica ce arata vezicule secretorii continand neurotransmitatori sau componente membranare ce se misca in directie anterograda; alte materiale, probabil membrane folosite se misca catre corpul celular in principal sub forma ce corpi multiveziculari; acestia isi varsa continutul in lizozomii corpului celular pentru digestia ulterioara; (C) legaturi incrucisate realizate de dineina / kinezina.
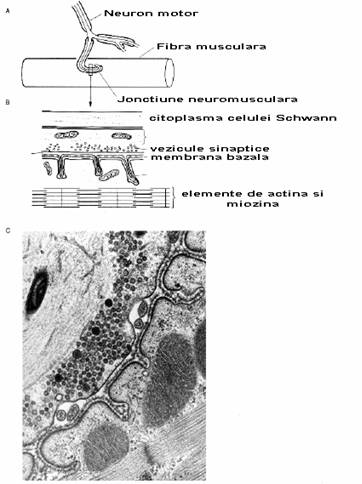 Fig. 2-15 : Jonctiunea neuromusculara (A) Fibre nervoase motorii inerveaza un
grup mic de fibre musculare (unitate motorie); (B) o portiune din unitatea
motorie; (C) jonctiune neuromusculara observata la ME - fibra musculara este in
dreapta jos, se observa membrana bazala, sarcolema fibrelor musculare.
Fig. 2-15 : Jonctiunea neuromusculara (A) Fibre nervoase motorii inerveaza un
grup mic de fibre musculare (unitate motorie); (B) o portiune din unitatea
motorie; (C) jonctiune neuromusculara observata la ME - fibra musculara este in
dreapta jos, se observa membrana bazala, sarcolema fibrelor musculare.
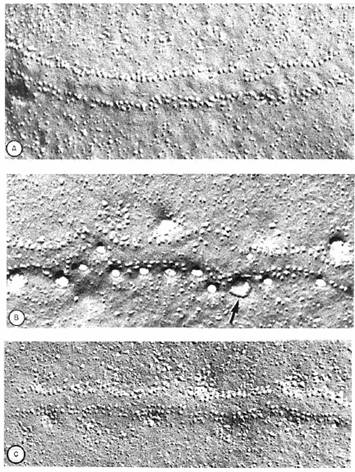
Fig. 2-16 : Membrana presinaptica a unei broaste - (A) Imagine la ME arata o membrana la 3ms dupa ce nervul a fost stimulat; (B) Membrana presinaptica la 5ms dupa stimulare; pete largi adiacente randurilor paralele de vezicule sunt vizibile; ele reprezinta locurile unde veziculele sinaptice au fuzionat cu membrana si au eliminat continutul; (C) Membrana la 50ms.
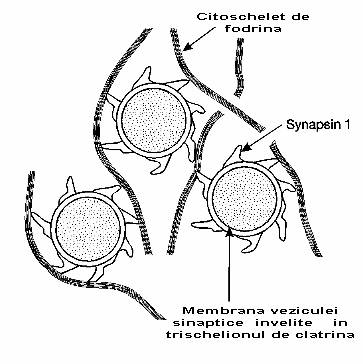
Fig. 2-17 : Atasarea veziculelor la citoscheletul de fodrina.
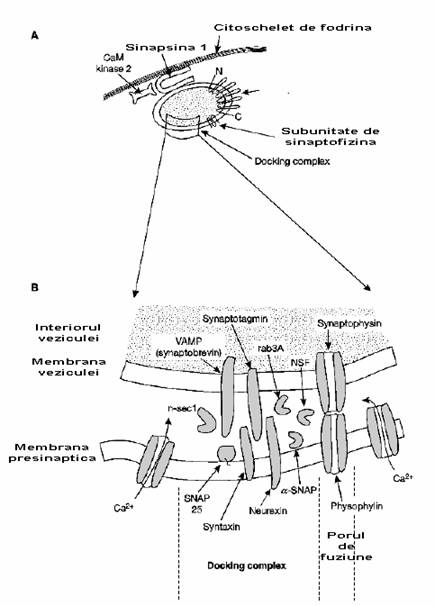
Fig. 2-18 : Veziculele ajung la nivelul membranei presinaptice - (A) O vezicula sinaptica este atasata prin intermediul sinapsinei la citoschelet; (B) Ilustrare a complexului de andocare (docking complex)
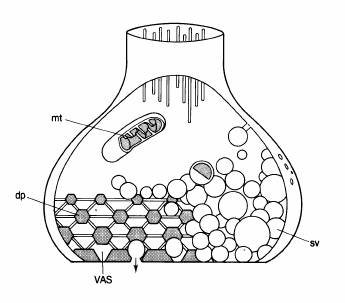 Fig. 2-19 : Organizarea unei retele presinaptice in cadrul unei sinapse centrale.
Fiecare spatiu triunghiular constituie un "loc de atasare a veziculei"(VAS)
unde veziculele sinaptice pot veni si isi pot eliberea continutul in fanta
sinaptica. dp=proiectii dense; mt=mitocondrii; sv=vezicule sinaptice.
Fig. 2-19 : Organizarea unei retele presinaptice in cadrul unei sinapse centrale.
Fiecare spatiu triunghiular constituie un "loc de atasare a veziculei"(VAS)
unde veziculele sinaptice pot veni si isi pot eliberea continutul in fanta
sinaptica. dp=proiectii dense; mt=mitocondrii; sv=vezicule sinaptice.
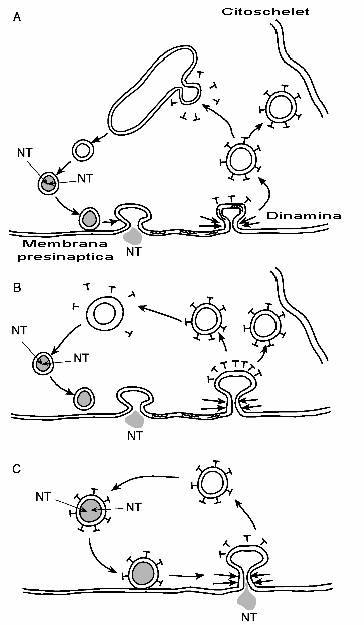 Fig. 2-20 : (A) Veziculele sinaptice fuzioneaza cu membrana presinaptica si isi
elibereaza continutul in fanta sinaptica; constituentii membranei
veziculare difuzeaza in membrana presinaptica si sunt recunoscuti de catre
clatrina sau sinapsina 1; in ambele cazuri, veziculele se formeaza prin
invaginare; moleclele de dinamina comprima vezicula si o inlatura din membrana
presinaptica; vezicule imbracate in sinapsina se ataseaza la citoschelet, iar
cele imbracate in clatrina pierd clatrina si fuzioneaza cu un endozom;
veziculele inmuguresc din endozom si sunt umplute cu neurotransmitatori. (B)
Ciclul (A) dar fara pasajul prin endozomi; (C) Ipoteza "kiss and go"
Fig. 2-20 : (A) Veziculele sinaptice fuzioneaza cu membrana presinaptica si isi
elibereaza continutul in fanta sinaptica; constituentii membranei
veziculare difuzeaza in membrana presinaptica si sunt recunoscuti de catre
clatrina sau sinapsina 1; in ambele cazuri, veziculele se formeaza prin
invaginare; moleclele de dinamina comprima vezicula si o inlatura din membrana
presinaptica; vezicule imbracate in sinapsina se ataseaza la citoschelet, iar
cele imbracate in clatrina pierd clatrina si fuzioneaza cu un endozom;
veziculele inmuguresc din endozom si sunt umplute cu neurotransmitatori. (B)
Ciclul (A) dar fara pasajul prin endozomi; (C) Ipoteza "kiss and go"
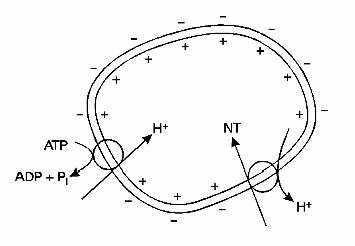
Fig. 2-21 : Umplerea veziculelor sinaptice cu transmitatori specifici - o pompa de protini inlatura H+ din vezicula; gradientul creat este folosit pentru a transport neurotransmitatorul (NT) in vezicula.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 5043
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved