| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Originea ARNm la ribovirusuri
La ribovirusuri, ARNm are origini diferite in functie de tipul de virus, de natura genomului sau (ARN mono- sau dublu catenar), dar in special in functie de polaritatea materialului genetic.
Polaritatea acizilor nucleici este stabilita conventional, in raport cu posibilitatea traducerii lor directe sau indirecte, in proteine. ARN genomic, tradus direct in proteine, fara sa necesite o etapa prealabila de transcriere este considerat a avea polaritate (complementaritate) pozitiva. ARN genomic, care este mai intai transcris intr-o molecula de ARNm complementar are polaritate negativa (fig. 87).
a
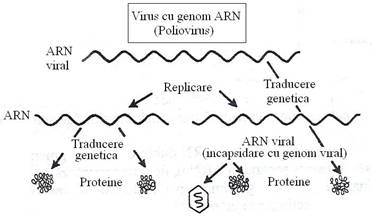
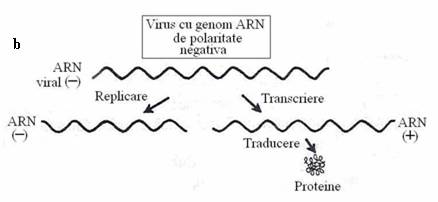
Fig. 87. Transcrierea si traducerea informatiei genetice la virusuri: (a) cu genom de polaritate pozitiva ; (b) cu genom de polaritate negativa
In ciclul de multiplicare a ribovirusurilor nu se face distinctia functionala intre ARNm timpuriu si tardiv, deoarece genomul viral functioneaza alternativ ca matrita pentru sinteza ARNm si a ARN genomic.
Dupa natura ARN genomic si dupa raportul sau cu ARNm, ribovirusurile infectioase pentru celulele animale, pot fi impartite in 6 clase:
clasa I cuprinde familiile Picornaviridae si Togaviridae, la care ARN genomic are rol de ARNm si codifica sinteza proteinelor virale structurale si functionale. Genomul acestor virusuri este infectios, pentru ca functioneaza direct ca ARNm pentru sinteza intregului set de proteine virale, inclusiv a ARN-polimerazei, care catalizeaza replicarea genomului viral.
Coronavirusurile au particularitatea ca genomul lor, cu polaritate pozitiva, este transcris initial de ARN-polimeraza virala, intr-o copie de sens negativ, de aceiasi lungime. Catena negativa, la randul ei, este transcrisa in ARNm monocistronic, de diferite lungimi;
virusurile clasei a II-a (Paramyxo-, Rhabdo-, Filoviridae, virusuri viscerotrope - Marburg, Ebola) au genom cu polaritate negativa. Genomul lor antimesager nu functioneaza ca ARNm si nu este transcris de celula. Virionul contine o transcriptaza proprie, ARN-polimeraza dependenta de ARN, care transcrie genomul de sens negativ, in ARNm monocistronic. Transcrierea este initiata la un singur promotor*, dar ARN-polimeraza recunoaste secventele semnal si la sfarsitul fiecarei gene, este eliberata copia de ARNm monocistronic.
Promotorii sunt secvente de ADN care regleaza expresia regiunilor codificatoare adiacente.
virusurile clasei a III-a (Myxoviridae) au genom segmentat, monocatenar, cu polaritate negativa. Fiecare segment genomic este transcris in propriul sau mesager. Fiind de polaritate negativa, transcrierea catenei genomice in ARNm este catalizata de o ARN-polimeraza dependenta de ARN, existenta in virion, analoga functional cu enzima de transcriere a altor ribovirusuri cu catena genomica ARN de polaritate negativa. ARN-polimeraza virusului influenza este defectiva din punct de vedere functional: nu initiaza sinteza ARNm si nici nu-l modifica prin bonetare sau metilare interna. Aceste functii sunt suplinite de ARN-polimeraza II a celulei, cea care catalizeaza initierea sintezei si metilarea ARNm celular;
clasa a IV-a (Arenaviridae), au genom ARN monocatenar, format din doua segmente. Particularitatea sa functionala consta in aceea ca jumatatea 3' a fiecarui segment genomic are o polaritate negativa si este transcrisa in ARNm complementar, de o transcriptaza virionica, iar jumatatea 5' a fiecarui segment are polaritate pozitiva. Pentru jumatatea 5', ARNm rezulta dupa o transcriere dubla: mai intai se sintetizeaza o copie a genomului si ulterior, aceasta este transcrisa in ARNm. Deoarece informatia este inscrisa in directii opuse in cele doua jumatati ale fiecarui fragment, strategia de codificare a genomului este ambisens.
La Bunyaviridae, genomul este de asemenea ambisens, deoarece atat ARN genomic
(de sens negativ), cat si catena complementara transcrisa ulterior (de sens pozitiv),
functioneaza ca ARNm;
clasa a V-a (Reoviridae), au genomul ARN dublu catenar, segmentat. Fiecare din cele l0 segmente genomice este transcris separat la ARNm, de o transcriptaza a virionului. Transcrierea este conservativa, deoarece catenele genomice parentale nu se separa si numai catena de complementaritate negativa functioneaza ca matrita. ARNm este monocistronic. ARN genomic ramane in regiunea centrala a virionului, iar copiile complementare sunt eliminate prin capsida partial dezintegrata.
clasa VI-a (Retraviridae), prezinta particularitatea ca genomul lor se replica printr-o copie intermediara de ADN. ARN genomic are complementaritate pozitiva, dar nu functioneaza ca ARNm. ARN pur nu este infectios pentru ca replicarea sa este dependenta de revers-transcriptaza asociata virionului, care transcrie ARN genomic intr-o molecula de ADN. ARNm este transcris din molecula de ADN, de ARN-polimeraza II celulara.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 2114
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2026 . All rights reserved