| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Citoplasma
Citoplasma celulei bacteriene este un sistem coloidal complex format din proteine, glucide, apa si substante minerale. Electrolitii sunt in concentratie mare si sunt retinuti in citoplasma datorita permeabilitatii selective a membranei. Concentratia mare a substantelor dizolvate - organice si anorganice, genereaza o presiune hidrostatica sau presiune de turgor, ce se manifesta in raport cu fata interna a membranei.
In citoplasma coexista starile de coacervare, de emulsie si de solutie, intr-o continua modificare a raportului lor cantitativ, care in ansamblu, datorita lipsei curentilor citoplasmatici confera caracterul unui gel fluid.
Caracterul de gel este consecinta starii apei. Molecula de apa este polara. Proprietatea de polaritate a apei este foarte importanta, deoarece multe molecule polare se dizolva usor in apa. Apa formeaza o retea tridimensionala atat cu ea insasi, cat si cu macromoleculele. Polaritatea inalta a moleculei de apa face ca moleculele nepolare sa formeze agregate stabile. Faptul ca moleculele de apa sunt legate in retea conditioneaza proprietatile sale de solvent, tensiunea inalta de suprafata si caldura specifica inalta. In celula, apa se gaseste in doua stari:
apa libera este cea care se deplaseaza liber in celula si care, la cresterea presiunii osmotice a mediului extracelular trece in afara celulei;
apa legata (de hidratare) in structura gelului citoplasmatic.
In celula bacteriana vegetativa, apa libera (in proportie de circa 70% din totalul apei) creeaza un mediu de suspensie pentru macromolecule.
Apa libera este foarte mobila, iar cea din structura gelului este practic imobila. Starea de gel a citoplasmei pastreaza componentele celulare separate spatial.
Absenta curentilor citoplasmatici are ca rezultat imobilitatea continutului celular.
La celulele bacteriene tinere, aflate in faza logaritmica de crestere, citoplasma este fin granulara, intens bazofila, datorita continutului ridicat de acizi nucleici, este densa si omogena. ARN citoplasmatic este reprezentat de ARNr (80%), ARNt (l0-20%) si ARNm (2%).
In citoplasma celulelor tinere, proteina dominanta cantitativ este EFTu (Elongation Factor Termo-unstable), care reprezinta circa 5% din totalul proteinelor. Acest factor participa la formarea legaturilor dipeptidice, la nivelul ribosomilor. Fiecare complex molecular ARNt-aminoacid ce asteapta recunoasterea codonului este asociat cu EFTu. Abundenta EFTu este reflectarea directa a ratei inalte a sintezei proteice.
In citoplasma celulelor imbatranite apar materiale de incluzie si vacuole. Aspectul devine granular neomogen, iar bazofilia diminua datorita scaderii pana la 0 a ratei sintezei ARN, precum si datorita reducerii treptate a cantitatii existente. Afinitatea pentru colorantii bazici este neuniforma.
In mediile naturale, bacteriile sunt supuse unor variatii ample ale presiunii osmotice a mediului extern. De exemplu, bacteriile din sol supravietuiesc perioadelor de uscaciune si umiditate, bacteriile infectioase ale tractului urinar supravietuiesc variatiilor de concentratie a urinii, iar microorganismele industriale tolereaza solutii nutritive concentrate, precum si acumularea extracelulara a produselor de metabolism.
La schimbarea osmolalitatii mediului lor, bacteriile raspund prin mecanisme pasive sau active. Raspunsul este prompt, deoarece membrana citoplasmatica este liber permeabila pentru apa.
La cresterea presiunii osmotice a mediului extern, bacteriile raspund in trei faze care se suprapun partial:
deshidratarea(pierderea unei parti a apei celulei);
acumularea cosolventilor;
remodelarea celulara(a peretelui, a nucleoidului, reluarea sintezei macromoleculelor, a cresterii si a diviziunii celulare).
Raspunsul la scaderea presiunii osmotice parcurge de asemenea trei faze:
inglobarea apei
eliminarea cosolventilor
remodelarea functiilor celulare.
Cosolventii sunt molecule solubile in apa, care echilibreaza presiunea osmotica a celulei cu presiunea mediului extracelular. De cele mai multe ori, rolul de cosolvent il au cationii K+, Na+ si cosolventii organici cu sarcina pozitiva (ectoina, glicin-betaina).
Cosolventii se gasesc in interiorul celulei si la exteriorul ei: NaCl si glucidele predomina in mediul extracelular, iar K+ si cationii organici, care constituie solventii compatibili sunt intracelulari.
Modificarile presiunii osmotice a mediului extern sunt receptionate de osmosenzori, localizati in membrana citoplasmatica si in nucleoidul bacterian. Osmosenzorul este un ansamblu care detecteaza schimbarile in activitatea apei extracelulare (osmosenzor direct) sau schimbarile care rezulta in structura celulei (osmosenzor indirect). Spre deosebire de chemosenzor, care se bazeaza pe interactiunea stereospecifica dintre ligand si receptor, osmosenzorii sunt macromolecule care sufera modificari ale conformatiei si/sau oligomerizare, ca raspuns la schimbarile solventului.
Microorganismele traiesc in medii cu intregul spectru al concentratiei de sare, de la apa dulce si mediul marin, pana la mediile in care NaCl are o concentratie la saturatie. Bacteriile halofile apartin grupului moderat, adica acelea care cresc la concentratii de NaCl cuprinse intre 2-15%(0,3-2,5 M) sau sunt halofile extreme, adica necesita cel putin 15% NaCl (2,6 M) pentru crestere. Bacteriile halotolerante cresc in absenta NaCl, dar tolereaza concentratii relativ mari de sare.
Microorganismele halofile si halotolerante apartin tuturor celor trei domenii: Archaea, Bacteria si Eucarya.
Deoarece membranele biologice sunt permeabile pentru apa, celulele nu pot mentine o activitate a apei citoplasmatice, mai mare decat a mediului salin inconjurator, deoarece apa trece rapid in mediu.
Microorganismele care traiesc in medii cu salinitate foarte mare, pastreaza un echilibru izoosmotic cu mediul extracelular. Mentinerea presiunii de turgor* necesita o presiune hiperosmotica. Cu exceptia halofilelor Archaea din ordinul Halobacteriales, toate organismele halofile isi pastreaza presiunea de turgor.
Presiunea de turgor este diferenta de presiune osmotica (osmolalitate) dintre interiorul si exteriorul celulei.
Microorganismele evita stresul osmotic determinat de concentratiile saline inalte, prin doua mecanisme:
celulele pot sa pastreze concentratii saline intracelulare inalte (strategia salt-in), cel putin echivalente din punct de vedere osmotic cu concentratiile externe. Toate sistemele intracelulare vor fi adaptate prezentei concentratiei saline mari;
celulele pot sa-si mentina concentratii saline scazute in citoplasma, dar presiunea osmotica inalta a mediului extern este echilibrata de solutii compatibile organice (strategia solutiilor compatibile). Solutiile compatibile sunt compusi organici cu molecula mica, solubile la concentratie mare in apa si sunt lipsite de sarcina, sau sunt pozitive la pH fiziologic.
Strategia "salt-in" este utilizata de doua grupuri neinrudite filogenetic: halofilele extreme din Archaea (ordinul Halobacteriales) si halofilele anaerobe (ordinul Haloanaerobiales). Cu o exceptie, ele nu contin solutii osmotice organice, iar concentratia ionica intracelulara este similara celei din mediul extern. In mediul extern se gaseste NaCl, iar in celula se gasesc concentratii echimolare de KCl.
La aceste celule, toate enzimele si componentele structurale sunt adaptate la prezenta concentratiei saline mari. Enzimele lor sunt tolerante la conditiile de salinitate. Proteinele lor au aminoacizi acizi in mare exces si cantitati mici de aminoacizi hidrofobi.
Cele mai multe proteine ale Archaea halofile depind de prezenta concentratiilor mari de sare pentru mentinerea conformatiei adecvate activitatii. Sursa de energie pentru eliminarea Na+ si acumularea KCl este gradientul electrochimic de protoni prin membrana citoplasmatica, rezultat in procesul respirator.
La majoritatea organismelor halofile si halotolerante, echilibrul osmotic este asigurat de molecule organice mici, fie sintetizate de celule, fie preluate din mediu cand acestea sunt disponibile. Strategia solutiei compatibile nu necesita prezenta proteinelor adaptate la mediul salin. Solutiile organice compatibile sunt acelea care, la concentratii inalte, permit enzimelor sa functioneze eficient, deoarece, probabil interactioneaza cu proteinele celulei.
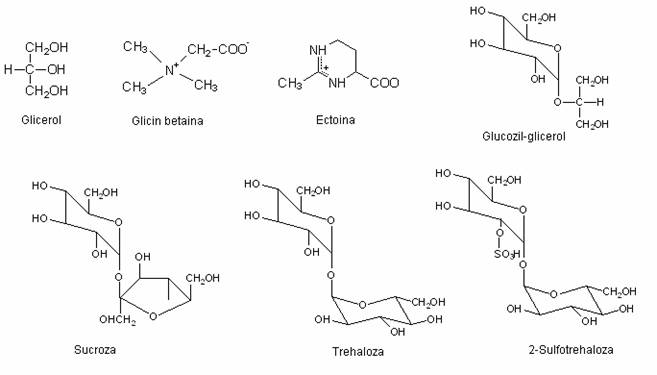
Compusi organici compatibili la microorganismele halofile si halotolerante.
Solutiile compatibile detectate la microorganismele halofile si halotolerante includ poliolii (glicerol, arabitol), glucide si derivatii lor (sucroza, trehaloza, glucozil-glicerol), aminoacizi si derivatii lor, amine quaternare (glicin-betaina). Betainele reprezinta un grup restrans de substante naturale care rezulta din aminoacizi, prin metilarea completa a gruparii aminice. Derivatii quaternari ce se formeaza sunt amfiioni, cu formula generala:
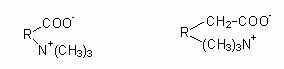
Multe bacterii care acumuleaza ori sintetizeaza substante organice pentru stabilizarea osmotica, contin si concentratii mari de Na+ si K+.
Procesele vitale in medii saline si hipersaline sunt costisitoare din punct de vedere energetic. Dintre toate solutiile organice compatibile, glicerolul este molecula cea mai simpla si cea mai ieftina pentru celula. Este miscibil cu apa in orice proportie. La Bacteria si Archaea, acumularea glicerolului nu este posibila, datorita absentei sterolilor membranari. Pentru echilibrul osmotic, ele sintetizeaza alte molecule mai costisitoare din punct de vedere energetic.
Solutiile care se sintetizeaza cu cea mai mica cheltuiala de energie se gasesc la organismele care traiesc la concentratiile saline cele mai mari si care necesita cea mai mare concentratie de solutie compatibila.
Pentru sinteza unei molecule organice compatibile sunt necesare 23-79 molecule de ATP la heterotrofe, produse in timpul respiratiei si 30-109 molecule de ATP pentru biosinteza, la autotrofe. Dar moleculele compatibile pentru solutia osmotica pot sa reprezinte si o sursa de carbon si energie, utilizabila in conditiile epuizarii resurselor energetice externe. Cand celulele trec in mediu cu salinitate scazuta (de exemplu, in lacurile sarate, cand apa de ploaie se amesteca cu apa sarata), o parte a moleculelor organice cu rol osmotic devin disponibile. Unele molecule (glicin-betaina si ectoina) nu sunt metabolizate, ci sunt excretate dupa trecerea celulei in mediul cu presiune osmotica mai mica. In mediul hipoosmotic, la Dunaliella (alga verde unicelulara), glicerolul este convertit in amidon (osmotic inactiv).
Multe microorganisme halofile si halotolerante mentin un amestec de molecule osmotice compatibile si reglarea sintezei lor este conditionata de nevoile celulei.
Bacteriile care sintetizeaza molecule organice compatibile poseda proteine membranare de transport, care le permit sa le incorporeze din mediu, daca sunt disponibile.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 1850
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved