| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Replicarea ADN bacterian
Legaturile de H nu sunt singurele forte stabilizatoare ale moleculei de ADN. Bazele sunt hidrofobe, se stivuiesc si tind sa se separe de mediul apos. Rasucirea helixului aduce bazele mai aproape si apa este exclusa mai eficient. Structura dublu catenara este stabilizata prin interactiunile hidrofobe dintre bazele celor 2 catene.
Desi bazele sunt hidrofobe si foarte putin solubile in apa, acizii nucleici sunt destul de solubili, datorita hidrofiliei axei glucid-fosfat si in special, datorita concentratiei mari de grupari fosfat incarcate negativ. Acestea pot avea efect invers, de respingere electrica a celor doua catene, dar prezenta sarurilor in mediu, creeaza un nor de ioni pozitivi care anuleaza respingerea.
Replicarea cromosomului bacterian este un proces strict reglat si are loc intr-o etapa bine definita a ciclului celular.
Replicarea ADN bacterian este de tip semiconservativ: fiecare din cele doua molecule fiice contine o catena sintetizata de novo si o catena complementara, conservata din molecula parentala (fig. 27).
Pentru ca replicarea ADN sa aiba loc, cele doua catene ale moleculei parentale trebuie sa se separe fizic, cel putin pe o secventa scurta, pentru a permite ADN-polimerazei si proteinelor asociate sa citeasca fiecare catena individuala cu rol de matrita. Reasocierea catenelor separate este blocata prin legarea SSB(single stranded DNA-binding protein).
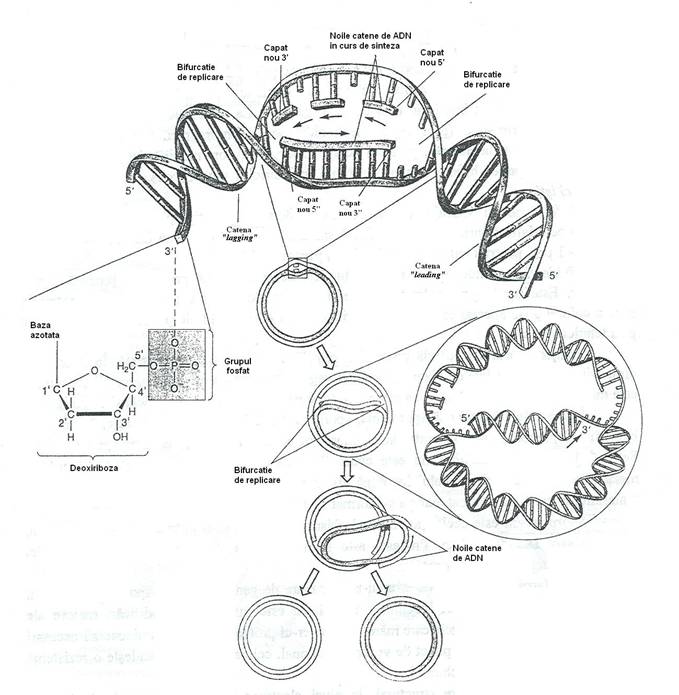
Fig. 27. Pentru replicarea ADN la
procariote, catenele se separa la punctul de origine a replicarii, de
unde bifurcatia de replicare se deplaseaza simetric. Fiecare dintre
cele doua catene are rol de matrita pentru sinteza unei catene
complementare. Sinteza noilor catene de ADN are loc in directia 5'- 3'. De aceea, sinteza
unei noi catene este discontinua, sub forma segmentelor scurte (
Cromosomul bacterian este un singur replicon*, deoarece se replica de la o singura origine. Procesul replicarii moleculei de ADN este initiat la un punct denumit originea replicarii. Aceasta este o secventa de 250-300 de baze, recunoscuta de complexul de initiere. La punctul de origine a replicarii, dubla catena de ADN este sectionata si replicarea este initiata pe cele doua catene izolate. Situsul de replicare, denumit bifurcatie de replicare se deplaseaza bidirectional, pe molecula de ADN.
Complexul proteic de replicare este format din proteina A (cu rol in legarea cromosomului pe situsul membranar), helicaza B - o topoizomeraza I (cu rol de despiralizare a ADN), proteina SSB, cu rolul de a stabiliza cele doua catene separate, ADN giraza (topoizomeraza II), ce limiteaza derularea extensiva a ADN sub actiunea topoizomerazei I si induce supraspiralizarea negativa a ADN nou replicat). Aceste proteine,
Cromosomii celulei eucariote au fiecare mai multi repliconi, ceea ce face ca replicarea ADN sa se faca intr-un interval scurt.
impreuna cu G-primaza (cea care sintetizeaza ARN primer) formeaza un primosom (complexul primar care initiaza replicarea). Sunt necesare doua complexe de replicare: unul se deplaseaza in directia 3'-5' pe un lant, iar celalalt, in directia 5' - 3', pe catena opusa.
Deoarece replicarea moleculei de ADN este bidirectionala, exista doua puncte de sinteza, denumite bifurcatii de replicare, care se deplaseaza in directii opuse, incepand de la punctul de origine si progreseaza cu 500-1000 nucleotide/sec., pana se intalnesc. Pe imaginile autoradiografice se observa structuri de forma literei theta.
Replicarea ADN are caracter cooperant, adica complexul de proteine implicate in replicare formeaza un ansamblu stabil care se deplaseaza de-a lungul fiecarei catene, fara disociere si reasociere la fiecare treapta, ceea ce contribuie decisiv la viteza foarte mare a replicarii ADN.
Enzima care catalizeaza adaugarea nucleotidelor este ADN-polimeraza III.
Replicarea ADN este semidiscontinua: o catena a ADN este sintetizata continuu, iar cealalta se sintetizeaza sub forma unor fragmente scurte, care ulterior sunt reunite cap la cap. Cele doua catene ale moleculei parentale nu se replica simultan, deoarece sunt antiparalele (una are directie 5'- 3', iar cealalta, 3'- 5'). Directia de polimerizare a catenei noi este 5'- 3', nici una dintre polimeraze neputand sa adauge dezoxiribonucleotide in directia 3'- 5', la catenele nascente.
Catena care creste in directia 5'- 3' (catena leading) se sintetizeaza in mod continuu, deoarece, permanent exista un capat 3'OH liber la bifurcatia de replicare, la care se adauga o noua nucleotida. ADN-polimeraza III catalizeaza adaugarea unei nucleotide la catena de ADN preexistenta, dar nu initiaza sinteza ADN de novo, dintr-un amestec de nucleotide. Totdeauna este necesar un primer de ARN. La deschiderea dublei catene de ADN, o ARN-polimeraza catalizeaza sinteza unui primer de ARN. ARN-polimeraza specifica se numeste ARN-primaza.
Catena care creste in directia 3'- 5' se sintetizeaza discontinuu, deoarece la bifurcatia de replicare nu exista un capat 3'OH liber. Mici fragmente de ARN primer furnizeaza capatul 3'OH. Aceasta este catena succesoare sau intarziata (lagging). Ea se sintetizeaza sub forma unor fragmente scurte in directia opusa bifurcatiei de replicare.
Dupa ce ADN este sintetizat, primerii ARN sunt indepartati (probabil sub actiunea RN-azei H). Fragmentele scurte rezultate din sinteza discontinua sunt completate de ADN-polimeraza I si reunite cap la cap sub actiunea catalitica a polinucleotid-ligazei.
Deoarece o catena este sintetizata continuu, iar cealalta discontinuu, intregul proces al replicarii ADN are caracter discontinuu.
Pentru un scurt interval dupa replicare, catena parentala si cea progena difera din punct de vedere chimic, deoarece catena parentala este metilata. Aceasta diferenta de ordin chimic este utila pentru corectarea fidelitatii copierii catenei matrita (functia de proof-reading). Astfel, catena noua este controlata pentru eventualele imperecheri gresite ale nucleotidelor. Eventualele erori de imperechere sunt depistate de sistemul enzimatic de corectare, sunt clivate, dupa care o enzima de reparare incorporeaza nucleotidele corecte.
ADN-metilazele metileaza adenina si citozina si inhiba astfel actiunea enzimelor de restrictie.
Cele mai importante schimbari ale ADN in cursul replicarii sunt acelea ale gradului de spiralizare, produsa sub actiunea topoizomerazelor. ADN suprahelical se despiralizeaza mai usor decat moleculele cu un grad inferior de spiralizare.
Deoarece topoizomerazele regleaza nivelul de supraspiralizare, ele sunt de fapt reglatoare ale proceselor de replicare si transcriere.
Transcrierea. Spre deosebire de ADN, care se sintetizeaza intr-o perioada definita a ciclului celular, sinteza ARNr se desfasoara continuu, pe tot parcursul ciclului de crestere a celulei. La eucariote, sinteza ARN in timpul mitozei este stopata, deoarece, cromosomii fiind condensati, transcrierea nu poate avea loc.
Informatia genetica este convertita la ARNm. Procesul este catalizat de ARN-polimeraza si este foarte asemanator cu cel al replicarii ADN, pentru ca ADN are rol de matrita si secventa de baze in noua molecula sintetizata reflecta pe aceea a matritei. Intre transcriere si replicare sunt doua diferente majore: a)se sintetizeaza molecule scurte; b)este transcrisa numai o catena a ADN.
Unele gene sunt transcrise de pe o catena, iar altele de pe catena opusa, dar in general, o secventa a ADN este transcrisa dintr-o singura catena.
ARN-polimeraza initiaza transcrierea la secventele de start denumite promotori, iar la capatul mesajului se gasesc semnalele care stopeaza polimerizarea, cand blocul de gene a fost transcris. ARN-polimeraza este alcatuita din 5 catene polipeptidice: doua catene α identice, β si β' si s (sigma). Primele 4 catene formeaza regiunea centrala a enzimei, dar specificitatea legarii de promotor se datoreaza celei de a 5-a subunitati - factorul sigma. Cele 4 catene, impreuna cu factorul sigma, formeaza holoenzima.
ARN-polimeraza este mare si intra in contact simultan, cu mai multe baze. Moleculele de ARN-polimeraza interactioneaza aleatoriu cu ADN cromosomal, aluneca de-a lungul ei, dar au afinitate mica fata de majoritatea secventelor. Polimeraza se leaga foarte strans de secventa specifica denumita promotor, ce contine situsul de start pentru initierea sintezei ARN.
Dupa ce ARN-polimeraza se leaga de regiunea promotor, structura initiala dublu catenara inchisa a ADN este convertita la forma deschisa, in care cele doua catene se separa pe o secventa localizata. Deschiderea dublei catene expune bazele lantului codificator, permitand imperecherea bazelor ribonucleozid-trifosfati pentru sinteza ARN. Se formeaza prima legatura fosfodiesterica si factorul s se disociaza din complex. Pentru extensia catenei de ARN este necesara numai regiunea centrala a ARN-polimerazei. Transcrierea ARN nu necesita primer si se face in directia 5'- 3', prin adaugarea secventiala a nucleotidelor la capatul 3'. Transcrierea progreseaza pana la un semnal terminal, cand ARNm si ARN-polimeraza sunt eliberate.
Semnalele de terminare a transcrierii. Transcrierea incepe la situsul de start (promotor) si se termina la sfarsitul genei, marcata de o secventa terminator (stop), caracterizata prin prezenta unei regiuni cu secvente repetate invers (fig 38).
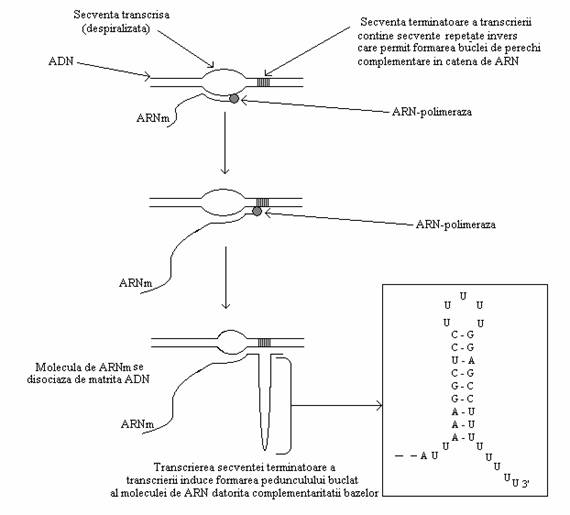
Fig. 28. Ilustrarea mecanismului molecular al terminarii transcrierii ARNm (modificat dupa Dale, 1996).
Secventa terminala va fi transcrisa in ARN, cu formarea structurii stem-loop (peduncul cu bucla) ceea ce va permite reasocierea celor 2 catene de ADN. Astfel, transcrierea se intrerupe si ARNm se disociaza.
Molecula de ARN contine secventa complementara a secventei genice, adica secventa catenei care nu a avut rol de matrita.
La bacterii, promotorii puternici sunt asociati cu gene ce sunt transcrise intr-un numar mare de molecule de ARN.
In faza de crestere logaritmica a culturii bacteriene, la temperatura optima (37 oC), cromosomul este transcris cu o rata de 60 nucleotide/secunda. In orice moment, in celula se gasesc 400-800 molecule identice de ARNm, circa l00 molecule diferite de ARNt si circa 700 molecule precursoare ale ARNr. Transcrierea fiecarei molecule de ARN este catalizata de o molecula de ARN-polimeraza. Cele circa l600 molecule de ARN-polimeraza reprezinta l% din cantitatea de proteine celulare. La procariote, toate tipurile de ARN sunt transcrise de un singur tip de ARN-polimeraza.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 3240
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved