| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Respiratia anaeroba
Majoritatea organismelor capabile de respiratie anaeroba sunt procariote. Unele dintre bacteriile heterotrofe care realizeaza respiratia anaeroba au catena de respiratie si sunt facultativ anaerobe. In prezenta O2, realizeaza metabolism oxibiotic. Transformarile chimice pe care le realizeaza in timpul generarii energiei, in absenta O2 au o deosebita importanta ecologica sau industriala.
In absenta O2, bacteriile heterotrofe facultativ-anaerobe isi obtin energia din substratul energetic, fie prin respiratia anaeroba, fie prin fermentatie. Respiratia anaeroba este mai eficienta din punct de vedere energetic, comparativ cu fermentatia.
In procesul respiratiei anaerobe, e- cedati de substratul organic oxidabil sunt preluati de catena de respiratie celulara si sunt transferati unui acceptor final, care este fie un compus oxigenat (nitrat, sulfat, carbonat), fie unui singur compus organic (fumaratul) (fig. 57). Transferul e- de-a lungul catenei respiratorii este cuplat cu fosforilarile oxidative membranare.
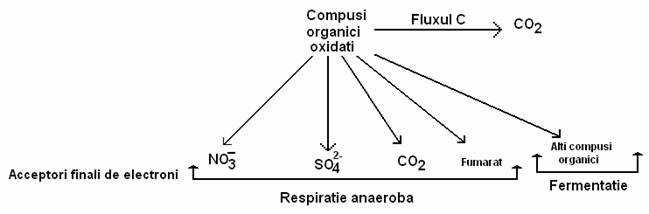
Fig. 57. Acceptorii finali de electroni in respiratia anaeroba si in fementatii. Catabolismul compusilor organici produce CO2.
Respiratia nitratilor
Unele bacterii realizeaza reducerea dezasimilatorie a nitratilor la nitriti, iar altele reduc atat nitratii la nitriti, precum si nitritii la oxizi de azot si ulterior pana la N2. Procesul fiziologic al reducerii nitratilor la N2 se numeste denitrificare si are loc atat in mediile naturale cat si in vitro. Nitratul, nitritul si respectiv oxizii de azot au rolul de acceptori de e-, care deriva din oxidarea unor compusi organici, pentru producerea energiei.
La E. coli, in conditii de anaerobioza, sistemul enzimatic al reducerii nitratului este inductibil. In prezenta lactatului (substrat nefermentabil) ca sursa de carbon si energie si a nitratului este indusa sinteza nitrat-reductazei (NAR), o enzima ce contine Mo. De aceea, procesul denitrificarii este strict anaerob. Reducerea nitratului are loc dupa reactia:
![]()
Reactia este dezasimilatorie, deoarece nitratii sunt redusi la nitriti, care, in cazul E. coli se acumuleaza in mediu si au un efect toxic, inhibitor pentru crestere, nitritul nefiind o sursa de azot asimilabil. Reducerea dezasimilatorie limitata a NO3- la NO2- se numeste respiratia nitratului.
La unele bacterii denitrificatoare, in procesul respiratiei anaerobe a nitratilor, este indus tot setul de enzime reducatoare, astfel incat reducerea nitratilor este totala, pana la N2, in urmatoarele etape:
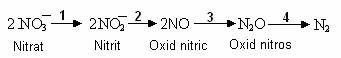
Reactia (1) este catalizata de NAR(A), treapta a 2-a, de nitrit-reductaza, iar reactiile 3 si 4, de enzime necunoscute.
Fiecare treapta de reducere este cuplata cu sinteza ATP. Unele procariote catalizeaza numai o parte a reactiilor caii reducerii nitratilor. Calea incompleta este utilizata in doua situatii:
cand bacteriile poseda echipamentul enzimatic necesar celor 4 trepte, dar in mediu se gasesc numai intermediari mai redusi decat nitratul;
cand bacteriile au incapacitati genetice de a cataliza una sau alta din treptele reducerii.
In functie de incapacitatile genetice de sinteza, variantele biochimice ale denitrificarii sunt urmatoarele:
organisme capabile sa reduca nitratul la nitrit, carora le lipsesc enzimele pentru treptele 2, 3, 4;
organisme capabile sa reduca nitratul pana la N2O (oxid nitros), carora le lipseste enzima pentru catalizarea treptei a 4-a;
organisme capabile sa reduca nitritul, la N2, dar nu au capacitatea de a cataliza reactia treptei l;
organisme capabile sa reduca nitratul la nitrit si oxidul nitric la N2O, carora le lipsesc enzimele ce catalizeaza reactiile 2 si 4.
Majoritatea bacteriilor apartin primei categorii. Nu sunt denitrificatoare, deoarece activitatea lor nu este insotita de producerea N2 (gazos).
Functia respiratorie a NAR (A) permite bacteriilor sa creasca in anaerobioza in prezenta nitratului si a unei surse oxidabile, dar nefermentabila, de energie. Intr-un mediu fara substrat fermentabil si fara nitrat, E. coli creste numai in aerobioza.
In procesul respiratiei anaerobe, oxidarea substratului energetic este completa, deoarece ciclul Krebs este functional, ca si in respiratia aeroba. Oxidarea completa a unei molecule de glucoza are loc dupa urmatoarele reactii globale:
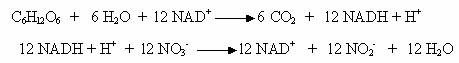
Pentru o molecula de glucoza oxidata se sintetizeaza 24 molecule de ATP, mult mai mult decat in fermentatie, dar mai putin decat in oxidarea aeroba. Oxidarea substratului organic in conditiile respiratiei anaerobe a nitratului este completa, singurul produs final fiind CO2. De aceea, bacteriile facultativ anaerobe, in prezenta nitratului, au un randament superior de crestere, in raport cu cresterea in conditii fermentative.
Unele bacterii(Shigella, Salmonella, Escherichia, Proteus, Brucella, Clostridium) reduc nitratul numai in absenta substratului care in mod obisnuit are rolul de acceptor de e-. In mediu se acumuleaza nitriti toxici care limiteaza cresterea. Alte bacterii sunt "specializate" pentru reducerea dezasimilatorie a NO3- si a NO2- pana la NO sau N2O, care ulterior sunt redusi la N2: Pseudomonas, Alcaligenes etc.
Cel mai adesea, bacteriile nitrat-reducatoare sunt heterotrofe si in prezenta O2 sunt aerobe. Ele prefera respiratia aeroba, deoarece au catena respiratorie. Catena functioneaza si in anaerobioza, dar la capatul ei, acceptorul final de e- este nitratul.
Nitrat-reductazele dezasimilatorii sunt proteine legate de membrana. In prezenta O2, sinteza lor este represata, dar este indusa in conditii anaerobe.
Reducerea dezasimilatorie a NO3- este limitata la bacterii, dar cele care catalizeaza acest proces sunt foarte diverse sub raport fiziologic.
Reducerea nitratului este o modalitate alternativa de respiratie si pentru bacteriile care oxideaza H2. Oxidarea H2 este cuplata cu reducerea NO3-, pana la N2.
Bacteriile sulfoxidante (Thiobacillus) oxideaza So, reactia fiind cuplata de asemenea cu reducerea NO3- la N2, dupa urmatoarea reactie globala:
![]()
Importanta ecologica. Bacteriile denitrificatoare sunt foarte raspandite in mediile naturale. Procesul denitrificarii este foarte important, deoarece constituie una dintre treptele circuitului azotului in natura (astfel se formeaza aproape tot N2 atmosferic), dar are efect negativ deoarece diminua randamentul productiv al fertilizatorilor cu azot, obtinuti pe cale industriala. Astfel se pierde 5-80% din cantitatea de fertilizatori cu azot.
Bacteriile denitrificatoare realizeaza scaderea concentratiei compusilor azotati din statiile de epurare a apelor uzate, inlaturand pericolul eutrofizarii bazinelor in care se deverseaza.
Denitrificarea este calea prin care, compusii cu azot, levigati din sol si transportati in oceanul planetar sunt reciclati la N2 atmosferic, acesta devenind din nou disponibil proceselor biologice prin fixare.
Unul dintre produsele reactiei de denitrificare - oxidul nitros (N2O) - difuzeaza spre stratosfera, unde, intr-o reactie fotochimica este convertit la oxid nitric (NO). NO reactioneaza cu O3, formand nitritul care se intoarce sub forma ploii acide. Rezultatul este distrugerea stratului de O3 care protejeaza organismele vii de radiatia UV.
Reducerea asimilatorie a nitratilor
Cele mai multe bacterii, dar si fungii, algele si plantele superioare au activitate nitrat-reductazica NAR(A), cu functie asimilatorie, adica enzima reduce nitratul la NH4+. Reducerea nitratilor la nitriti este urmata de o serie de reactii de reducere, consumatoare de energie, al caror rezultat final este formarea NH4:

Reducerea asimilatorie a nitratului nu este producatoare, ci consumatoare de energie.
Nitrat-reductaza asimilatorie este o proteina citoplasmatica ("solubila"). In prezenta NH4, sinteza ei este represata. La fiecare treapta a reactiei are loc transferul a 2 e-. Treptele reactiei reducerii asimilatorii au loc in conditii de aerobioza.
NH4 este sursa de azot asimilabil pentru toate bacteriile chimioorganotrofe.
La bacterii, NH4 este asimilat pe doua cai majore: calea glutamin-sintazei (GS) si glutamat-sintazei (GOGAT). O cale alternativa de asimilare, la multe bacterii, inclusiv enterobacterii este calea glutamat-dehidrogenazei (GDH), mai putin eficienta energetic decat caile GS/GOGAT. GDH are afinitate mai mica pentru amoniu si este ineficienta in celulele care cresc in conditii de limitare a sursei de azot.
Reducerea asimilatorie si cea dezasimilatorie a NO3- (denitrificarea) nu par a fi procese fiziologice corelate in mod obligatoriu. Unele bacterii catalizeaza ambele tipuri de reactii. Altele fac numai reactiile reducerii dezasimilatorii ale NO3 si nu pot sa-l asimileze, iar o alta categorie asimileaza NO3, dar nu fac reactia de denitrificare. La organismele capabile sa catalizeze ambele tipuri de reactii, distinctia dintre cele doua procese se face prin expunerea culturii la O2. De exemplu, P. aeruginosa poate asimila NO3 in prezenta sau in absenta O2, dar denitrificarea are loc numai in conditii de anaerobioza.
Respiratia sulfatului
Sulfatul, anionul major in apa marii, este cel mai oxidat compus al sulfului si are rol de acceptor final de e- in procesul oxidarii anaerobe a unor compusi organici de catre organismele unui grup fiziologic special, al bacteriilor sulfat-reducatoare.
SO42- si So au rol de acceptori de e- in conditii anoxice pentru un grup larg de bacterii chimioorganotrofe sau chimiolitotrofe hidrogen-oxidante. Ele poseda echipamentul enzimatic care catalizeaza reducerea dezasimilatorie a sulfatului. Produsul final al reducerii SO42- este H2S, care se poate acumula in cantitati mari in mediile naturale.
Bacteriile sulfat-reducatoare sunt strict anaerobe si oxideaza diferite substraturi organice. Ele constituie un ansamblu fiziologic si ecologic de tipuri morfologice diferite de bacterii anaerobe, care au in comun capacitatea de a activa SO42- si de a-l reduce la H2S.
Bacteriile din grupul I (Desulfovibrio, Desulfomonas, Desulfotomaculum) utilizeaza lactatul, piruvatul, etanolul, malatul etc., dar si H2 si reduc SO42- la H2S.
Substraturile organice sunt oxidate la acetat, deoarece bacteriile sulfat-reducatoare (Desulfovibrio, Desulfotomaculum) nu au echipamentul enzimatic al ciclului Krebs. Acetatul este secretat ca produs final.
Bacteriile din grupul al II-lea (Desulfobacter, Desulfococcus, Desulfonema) oxideaza acizii grasi si in special acetatul, pana la CO2 si reduc SO42- la So.
Desulfosarcina, Desulfococcus, Desulfobacterium, Desulfotomaculum sunt unice printre sulfat-reducatori prin capacitatea lor de a creste chimioautotrof cu H2 ca donor de e-, CO2 ca singura sursa de C si SO42- ca acceptor de e-.
Toate bacteriile sulfat-reducatoare sunt strict anaerobe. Reducerea dezasimilatorie a sulfatului este un proces respirator obligatoriu pentru acest grup fiziologic si nu o cale alternativa a respiratiei, asa cum este denitrificarea. Sulfatul este redus la H2S dupa urmatoarea reactie;
![]()
Bacteriile sulfat-reducatoare reprezinta una dintre formele vechi de viata ale planetei si participa la producerea si transformarea depozitelor minerale in natura.
Alti compusi oxidati ai sulfului (sulfitul - SO32-, tiosulfatul - S2O32-, tetrationatul - S4O62-) pot fi utilizati ca acceptori de e- si sunt redusi la H2S in procese dezasimilatorii. Reducerea lor este catalizata nu numai de bacteriile sulfat-reducatoare, ci si de alte grupe fiziologice.
Reducerea bacteriana a sulfatului este un proces important pentru mineralizarea materiei organice in mediile anoxice, in special in sistemele marine (cu salinitate de 1-4%, in care cresc Desulfovibrio, Desulfobacter, Desulfococcus) si in mediile hipersaline.
Oxidarea completa a materiei organice la CO2 cu reducerea simultana a SO42- la H2S nu depinde de fermentatiile sintrofice.
Pe langa capacitatea de a folosi SO42- ca acceptor de e-, multe bacterii sulfat-reducatoare pot folosi NO3- ca acceptor de e-, pe care-l reduc la NH3 sau fermenteaza anumiti compusi organici pentru producerea E, in absenta acceptorilor finali de e- (de exemplu, piruvatul este fermentat la acetat, CO2 si H2).
SO42- este un acceptor de e- mai putin favorabil cresterii bacteriene, comparativ cu O2 sau NO3-, dar se elibereaza suficienta E pentru sinteza ATP.
Bacteriile sulfat-reducatoare si metanogenele sunt doua populatii care intra in competitie metabolica pentru aceleasi substraturi. Ambele tipuri catalizeaza stadiile finale ale mineralizarii anaerobe a materiei organice si ambele depind de microorganismele fermentative care convertesc materia organica complexa, la compusi mai simpli (H2, CO2 si acetat).
Se accepta ca cele doua grupe se exclud reciproc. Mediile bogate in sulfat selecteaza bacteriile sulfat-reducatoare, iar cele fara sulfat selecteaza metanogenele. Totusi, cele doua grupe coexista in medii cu sulfat.
Reducerea So. Bacteriile care folosesc So ca acceptor final de e- genereaza H2S, dar nu reduc SO42- la H2S.
Reducerea asimilatorie a sulfatului
Bacteriile si organismele superioare (plante si animale) preiau sulful necesar din sulfat. Sulful are numarul de oxidare -2 in compusii organici si +6 in SO42-, o diferenta de 8e-. Asimilarea S implica reducerea SO42- la H2S, inainte de incorporarea sa in compusi organici. Aceiasi reactie de reducere are loc in procesul fiziologic al respiratiei anaerobe a SO42-, dar mecanismele enzimatice sunt diferite. Reducerea SO42- pentru a fi folosit ca sursa de H2S se numeste reducerea asimilatorie a sulfatului.
SO42- este activat de ATP. ATP-sulfurilaza catalizeaza legarea ionului SO42-, de un fosfat al ATP si rezulta adenozin-fosfo-sulfatul (APS). O alta grupare fosfat este adaugata la APS si se formeaza fosfo-adenozin-fosfo-sulfatul (PAPS), dupa care ionul SO42- este redus. Ca si in reducerea dezasimilatorie, primul produs al reducerii SO42- este sulfitul (S2O32-).
In reactiile de reducere asimilatorie a SO42-, H2S format este convertit la sulf organic, iar in cele de reducere dezasimilatorie, H2S este excretat.
H2S este incorporat in substratul organic, ca atom de S al cisteinei. La bacterii, substratul organic acceptor al S este O-acetilserina, dupa reactia;
![]()
Alti acceptori finali de e-
Fe3+ are rol de acceptor de e- in metabolismul energetic la o varietate de bacterii chimioorganotrofe si chimiolitotrofe pentru ca este abundent in mediile naturale. Potentialul reducator al Fe3+/Fe2+ este foarte electropozitiv (+ 0,77 V la pH 2). Reducerea Fe3+ poate fi cuplata cu oxidarea unor donori de e-, organici sau anorganici.
Geobacter metallireducens oxideaza acetatul, odata cu reducerea Fe3+ la Fe2+. Fe3+ este unul dintre cele mai comune metale in sol si roci. Reducerea Fe3+ la Fe2+ este importanta deoarece furnizeaza o forma mai solubila a Fe.
Mn metalic are mai multe stari de oxidare, Mn4+ si Mn2+ fiind cei mai stabili si cei mai relevanti. Bacteriile chimioorganotrofe fac reducerea anoxica a Mn4+ la Mn2+. Potentialul redox al cuplului Mn4+/Mn2+ este foarte mare, astfel ca unii compusi organici pot dona e- pentru reducerea Mn4+.
Alti compusi anorganici care pot functiona ca acceptori de e- in respiratia anaeroba sunt Se si As. In natura nu se gasesc in cantitati mari, dar sunt poluanti si pot suporta cresterea anoxica a unor bacterii. Reducerea SeO42- (selenat)la SeO3- (selenit)si chiar la Seo (metalic) este o metoda importanta pentru indepartarea Se din apa (bioremediere).
Respiratia fumaratului
In procesul respiratiei anaerobe, rolul de acceptor final de e- il au nu numai compusii anorganici (nitrati, nitriti, sulfatul), ci si fumaratul, un compus organic.
n respiratia fumaratului, transferul de e- se face, ca si in procesul respiratiei aerobe, pe calea unei dehidrogenaze, a unei lipoquinone, unul sau mai multi citocromi b si a unei fumarat-reductaze (o feredoxina), legata covalent cu grupul prostetic FAD. Fumarat-reductaza catalizeaza urmatoarea reactie:

Fumarat-reductaza oxideaza NADH, lactatul, formiatul, glicerolul. Sursele de fumarat ale celulei sunt multiple: fumaratul se formeaza din malat, aspartat, piruvat. Reducerea unei molecule de fumarat produce o molecula de ATP. E. coli creste intr-un amestec de H2 sau formiat si fumarat ca sursa de carbon si energie.
Trimetil-amin-oxidul (TMAO) este un acceptor organic de e-, fiind si o substanta de echilibru osmotic la pestii marini, unde are rolul de a excreta excesul de N.
O varietate de bacterii reduc TMAO la TMA (trimetil-amina). TMA are un miros puternic si o parte din mirosul degajat de pestele marin alterat se datoreaza TMA formata prin actiunea bacteriilor facultative care pot folosi TMAO ca acceptor de e-. Un analog al TMAO este DMSO (dimetil-sulfoxid), care este redus de unele bacterii la DMS (dimetil-sulfit).
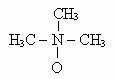
Trimetil amin-oxid
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 4422
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved