| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
1.1. Sistemul radicular
Sistemul radicular variaza in functie de specie. La o serie de taxoni fixati, radacina se formeaza din radicula embrionului-radacina principala (Sagittaria, Potamogeton, Alisma, Myriophyllum, Ranunculus s.a.).
La alte specii fixate (Nymphaea, Nuphar, Vallisneria) este prezent un sistem rizomal bine dezvoltat, fixat in sol prin numeroase radacini adventive. La Nymphaeaceae, radacinile adventive au origine mezogena si provin din straturile corticale externe.
O serie de plante sunt initial slab fixate (Elodea canadensis, Trapa natans, Myriophyllum, Ceratophyllum) iar ulterior libere; astfel radacinile sunt absente la plantele mature (Fig. 3: B, D, E).
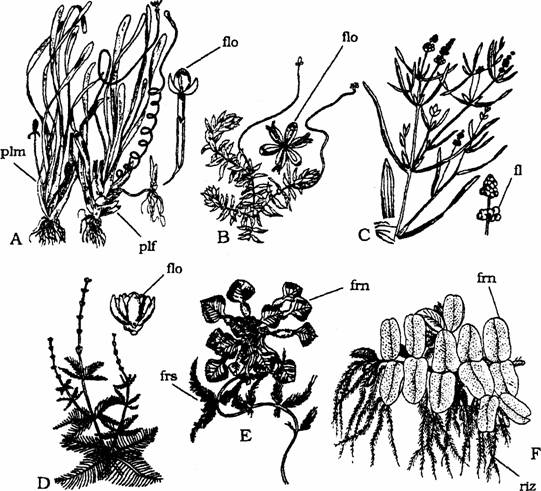
Fig. 3 - Hidrofite: A - Vallisneria spiralis, B - Elodea canadensis, C - Potamogeton pusillus, D - Myriophyllum spicatum, E - Trapa natans, F - Salvinia natans, flo - floare, frn - frunza natanta, frs - frunza submersa, plf - planta femela, plm - planta mascula, riz - rizofila (d. Sarbu, 1999)
Printre hidrofite exista si specii fara radacini (Wolffia arrhiza, Aldrovanda vesiculosa, Utricularia vulgaris, Salvinia natans). Majoritatea plantelor arhizice traiesc in apa, pe care o absorb usor prin intreaga suprafata. La Utricularia vulgaris (otratelul de balta) absorbtia se face prin frunze filiforme, iar la Salvinia natans (pestisoara) prin frunze puternic divizate, submerse, asemanatoare ca aspect cu radacina, avand functie absorbanta-rizofile.
La plantele submerse se intalnesc de asemenea radacini asimilatoare. La Lemna sp. radacina nu se ramifica si are rol asimilator.
1. Tulpina
Tulpinile sunt prezente la plantele submerse (Utricularia sp., Ceratophyllum sp., Myriophyllum sp.) sau cu frunze plutitoare (Nuphar sp., Nymphaea sp.) (Fig. 3-D).
Speciile de Salvinia, Potamogeton si Ceratophyllum au tulpini subtiri si cilindrice iar speciile de Nymphaea si Nuphar au tulpini groase si scurte, fixate pe fundul apei- rizomi (Fig.3: C, F).
Unele plante acvatice (Salvinia natans, Statiotes sp., Hydrocaharis morsus-ranae) au tulpini reduse. Recordul in aceasta directie il detine Wolffia arrhiza a carei tulpini nu depaseste marimea unui bob de mei.
Speciile de Lemna si Wolffia au tulpina taloidica, lipsita de frunze si mult redusa.
1.3. Frunza
Dispozitia frunzelor pe tulpina. La Elodea canadensis si Lysimachia vulgaris - se intalneste o dispozitie verticilata caracterizata prin dezvoltarea a trei sau mai multe frunze la fiecare nod (Fig.3-B).
Dintre adaptarile morfologice care apar la plantele care traiesc total sau partial cufundate in apa, se distinge dezvoltarea pronuntata a sistemului foliar in raport cu restul corpului.
Organele lor vegetative sunt foarte diferite de cele ale plantelor terestre. Frunzele a caror forma frapeaza ochiul nu se mai intalnesc la alte plante, ele fiind subtiri, semitransparente, moi, de un verde clar, alungite frecvent circulare sau divizate in filamente capiliforme.
Forma frunzei la plantele acvatice este foarte variata: Utricularia sp. prezinta frunze peltate transformate in vezicule; Salvinia natans prezinta limbul intreg, iar Sagittaria sp. are lamina in forma de sageata.
Frunzele pot fi subtiri si transparente (Elodea canadensis, Potamogeton lucens, Potamogeton pusillus), tubulare (Myriophyllum spicatum, Ranunculus aquatilis), in forma de benzi (Vallisneria spiralis).
O metamorfoza determinata de factorul apa este si dimorfismul foliar (heterofilia) caracteristic pentru speciile amfibii si natante care prezinta o parte a corpului cufundata in apa (submersa) si alta deasupra apei sau pe suprafata apei (emersa).
In acest sens Ranunculus aquatilis diferentieaza frunze plutitoare cordat reniforme 3-5 lobate si frunze submerse puternic sectate (Fig.4).
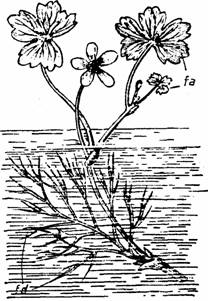
Fig. 4 - Dimorfism foliar la Ranunculus aquatilis: fa-frunze aeriene; fd-frunze dezvoltate in apa (d.Grintescu, 1985)
Trapa natans, o planta acvatica foarte raspandita in lacurile dunarene, se caracterizeaza deasemenea, prin existenta a doua categorii de frunze: unele submerse - liniare, intregi, sesile si cu o dispozitie opusa, iar cele natante au lamina triunghiulara (Fig.3-E).
Dimorfismul foliar este prezent si la Salvinia natans, feriga acvatica la care din cele trei frunze dispuse in verticil la fiecare nod al tulpinei, doua sunt intregi si inlesnesc plutirea plantelor la suprafata apei, iar a treia este filiform divizata si se prezinta morfologic si functional asemanator cu radacina, motiv pentru care a primit denumirea de rizofila (Fig.3-F).
La Sagittaria sagittifolia frunzele submerse sunt liniare, cele natante au limbul eliptic cordat, iar cele aeriene, foarte lung petiolate au limbul sagitat (Fig.5).
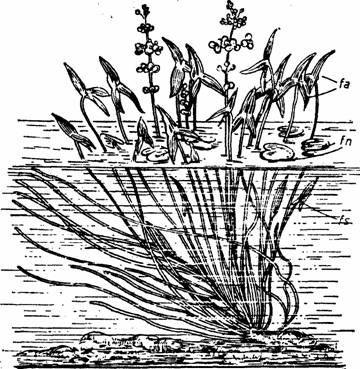
Fig. 5 - Polimorfism foliar la Sagittaria sagittifolia - fa-frunze aeriene; fn-frunze natante; fs-frunze submerse (d.Grintescu, 1985)
Alte metamorfoze care apar la nivelul limbului foliar sunt legate de nutritia heterotrofa complementara a unor taxoni: Utricularia vulgaris, Aldrovanda vesiculosa s.a.
Aldrovanda vesiculosa - planta acvatica submersa, fara radacini, are limbul usor bombat, ca o vezicula inotatoare, compus din doua jumatati mobile, indreptate una catre alta, dispuse sub un unghi de 90 de grade. Pe fiecare jumatate de limb se gasesc patru zone, fiecare cu o anumita activitate in actiunea de capturare si digerare a insectelor.
Utricularia vulgaris - planta acvatica submersa, fara radacini, are frunze alterne, sectat-divizate.Unele segmente ale frunzei sunt lacinii filiformi, altele transformate in vezicule semiglobuloase sau semiovale, comprimate lateral, cu partea ventrala mai mult sau mai putin plana, iar cea dorsala bombata. Vezicula se fixeaza pe tulpina ramificata cu ajutorul unui petiol relativ scurt. Deschiderea veziculei se afla la partea anterioara si are marginea indoita inauntru ca o palnie. Partea ei de sus, elastica si concrescuta lateral cu peretele veziculei, reprezentand un fel de supapa, iar partea de jos, opusa, a deschiderii are aspectul unei umflaturi in forma de potcoava. In jurul deschiderii se afla numerosi peri: unii setiformi, mici, simpli, altii lungi, ramificati.
Adaptari structurale
1. Sistemul radicular
In general plantele care traiesc in mediul acvatic, sufera sub actiunea factorului apa, o serie de modificari adaptative majore, ce se concretizeza prin structura lor hidromorfa.
La Lemna sp. radacina are rol de organ asimilator.
La unele plante acvatice (Lemna sp., Hydrocharis morsus-ranae, Statiotes sp.) piloriza este inlocuita de o formatiune speciala numita rizomitra, care nu se uzeaza si nu se reinoieste. Rizomitra este de alta origine decat piloriza (Fig.6).
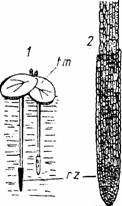
Fig. 6 - Lemna minor: 1-planta adulta; 2-rizomitra mult marita; rz-rizomitra; tm-tulpina metamorfozata (d.Grintescu,1985)
Rar la unele plante acvatice (Hydrocharis morsus-ranae s.a.) este prezenta piloriza (caliptra) tipica, formata in urma activitatii caliptrogenului (meristem primar radicular).
Formarea perilor absorbanti este legata de conditiiile de mediu. Majoritatea plantelor acvatice au radacini lipsite de peri absorbanti. Plantele acvatice absorb apa pe toata suprafata corpului.
Perisorii sugatori cresc perpendicular pe axa radacinii, prin alungirea celulelor rizodermice. La Salvinia natans ei sunt pluricelulari (7-9 celule).
La multe plante acvatice s-a observat ca nu toate celulele rizodermei dau nastere la peri absorbanti; la Sagittaria sagittifolia se transforma in peri sugatori numai celulele scurte iar la Hydrocharis morsus-ranae numai cele lungi; la Azzola caroliniana perii absorbanti sunt reuniti in tufe.
Dezvoltarea radicelelor. La unele hidrofite, primordiul radicelelor isi are originea in straturile cele mai interne ale scoartei.
La plantele acvatice submerse, cum ar fi Potamogeton lucens, de pe rizomul subteran se formeaza radacini adventive, caracterizate prin lipsa perisorilor absorbanti si a exodermei, prezenta unui sistem cortical de canale aerifere mari, reducerea elementelor mecanice si a celor xilemice din structura cilintrului central.
Cilindrul central. La Scirpus sp. periciclul este intrerupt in fata fasciculelor lemnoase iar la Potamogeton sp. si Vallisneria spiralis se intrerupe in dreptul fasciculelor liberiene.
La Hydrocharis morsus-ranae fasciculele lemnoase se reduc la un singur vas. La multe specii acvatice vasele lemnoase se resorb lasand lacune in locul lor; fasciculele liberiene se pot reduce si ele (Potamogeton sp., Valisneria spiralis, Hydrocharis morsus-ranae).
Scoarta. S-a constatat ca radacina in scoarta careia se gasesc cloroplaste, cum sunt radacinile de la Statiotes aloides, Hydrocharis morsus-ranae sunt lipsite de exoderma.
Tulpina
Tulpinile acvatice prezinta unele particularitati.
Epiderma. In general plantele acvatice submerse sunt lipsite de stomate (Vallisneria spiralis) sau daca se gasesc sunt de obicei nefunctionale.
Tulpina de la Myriophyllum sp., Elodea canadensis s.a prezinta tesutul cortical bine dezvoltat, strabatut de canale sau lacune aerifere; se observa de asemenea o reducere a numarului de vase lemnoase; cat priveste stereomul si el este de asemenea foarte redus (Fig.7).
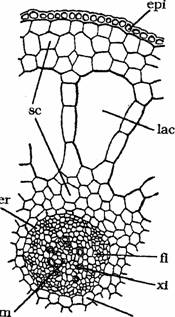
Fig. 7 - Structura tulpinii de Myriophyllum spicatum (sectiune transversala): end-endoderma; epi-epiderma; fl-floem; lac-lacuna corticala; m-maduva; per-periciclu; sc-scoarta; xi-xilem (d.Sarbu,1999)
Structura primara a tulpinii. Plantele acvatice prezinta endoderma tipica. Tulpinile plantelor acvatice ofera un alt exemplu de structura primara modificata insa sub actiunea factorului apa. Ca adaptare la viata acvatica, plantele submerse prezinta o structura hidromorfa, corpul lor fiind strabatut de canale aerifere, care inlesnesc atat schimbul de gaze cat si mentinerea in pozitie verticala.
Astfel luand ca exemplu tulpina de la Myriophyllum spicatum, se observa, de la inceput, modificarea raportului dimensional dintre scoarta si cilintrul central (Fig.7).
Scoarta alcatuita in cea mai mare parte dintr-un aerenchim prevazut cu un singur inel de canale aerifere mari se caracterizeaza printr-o dezvoltare puternica. Epiderma este unistratificata si nu se diferentieaza stomate. Endoderma este prezenta, fiind alcatuita din celule voluminoase dintre care foarte multe prezinta ingrosari Caspary in forma de banda (lenticulare in sectiune transversala). Tesutul mecanic este redus, ca de altfel si numarul vaselor lemnoase, din structura fasciculelor conducatoare.
Stelul. La Marsilea quadrifolia, stelul este de tip sifonostel amfifloic, adica intre lemn si maduva se mai formeaza un cordon de liber.
Variatii ale structurii primare. Structura scoartei este influentata de mediul in care traiesc plantele. De exemplu la plantele acvatice parenchimul cortical este lacunos; la unele se formeaza chiar canale aerifere (Ceratophyllum demersus, Myriophyllum spicatum, Nymphoides peltata). Parenchimul poate fi de origine schizogena (Trapa natans, Potamogeton sp., Ceratophyllum sp., Nymphaeaceae) sau poate fi de origine lisigena sau rexigena.
La unele plante acvatice maduva lipseste complet (Ceratophyllum sp., Myriophyllum sp., Utricularia sp., Salvinia natans). La Nymphaeaceae maduva este dezvoltata si tot atat de lacunoasa ca si scoarta.
In ceea ce priveste variatiile vaselor lemnoase este de mentionat in primul rand tendinta spre o reducere a numarului de vase la plantele acvatice.
Tesuturi secundare din tulpina. Datorita conditiilor de viata, la o serie de plante acvatice in lemnul secundar se remarca absenta traheelor.
3. Frunza
Epiderma. La plantele acvatice cu frunze plutitoare, stomatele se formeaza doar pe epiderma superioara-epistomatice (Trapa natans-300 stomate, Nymphaea alba- 648 de stomate). Plantele cu frunze submerse sunt lipsite de stomate functionale (Vallisneria spiralis).
La Nymphaeaceae stomatele sunt de tipul anomocitic si se caracterizeza prin celule stomatice inconjurate de un anumit numar de celule anexe care nu difera ca marime si forma fata de celelalte celule epidermice.
Numarul mediu de stomate pe milimetru patrat de limb foliar este cuprins intre 100-300. Numarul stomatelor pe milimetru patrat scade sau creste la aceeasi specie sau la diferite specii de plante fiind in functie de faza ontogenetica a plantei si de conditiile ecologice in care aceasta se dezvolta (Andrei, 1967).
La unele plante acvatice submerse, exista celule epidermice care se diferentiaza in hidropote (Fig.8).
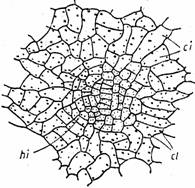
Fig. 8 - Structura unei hidropote la Nymphoides peltata: ci-celule ale epidermei inferioare; cl-cloroplaste; hi-hidropota (d.Andrei, 1976)
Hidropotele sunt structuri de natura epidermica alcatuite din una sau mai multe celule epidermice, grupate, specializate, permeabile pentru apa si saruri, de obicei mai mici decat celulele obisnuite. Celule epidermice care se diferentiaza in hidropote se numesc meristemoide ale hidropotelor. Ele se deosebesc de celelalte celule epidermice prin continutul bogat in citoplasma granulara. Meristemoidele hidropotelor se observa la Nymphaea alba si Nuphar lutea pe partea inferioara a frunzelor tinere inca din stadiul de mugure. La microscopul electronic meristemoidele hidropotelor sunt prevazute cu nucleu voluminos, vacuole putine, bogate in flavone, tanin, cloroplaste, mitocondrii, dictiozomi, reticul endoplasmatic.
In mod normal cuticula epidermei acopera si celulele diferentiate in hidropote. Analizata la microscopul electronic cuticula hidropotelor apare poroasa. Porii cuticulari corespund cu spatiile intermicrofibrilare ale peretilor tangentiali externi ai celulelor hidropotelor. La nivelul acestor pereti are loc schimbul de substanta dintre tesuturile vii ale frunzei si mediul inconjurator.
Structura limbului. Plantele acvatice cu frunze plutitoare au frunze bifaciale cu structura dorsiventrala heterofaciala (Nymphaea alba, Nuphar lutea, Trapa natans, Hydrocharis morsus-ranae s.a.). In acest caz in tesutul palisadic (3-4 straturi de celule), localizat sub epiderma superioara, se diferentiaza elemente mecanice sclerenchimatice (idioblaste), care confera frunzei rezistenta necesara. Tesutul parenchimatic lacunar prezinta lacune foarte mari, alcatuind de fapt un parenchim aerifer. In structura nervurilor, xilemul este slab reprezentat, dar existenta colenchimului angular, localizat intre nervura si epiderma inferioara, sporeste la acest tip de plante rezistenta laminei (Fig. 9; Fig. 10.2).
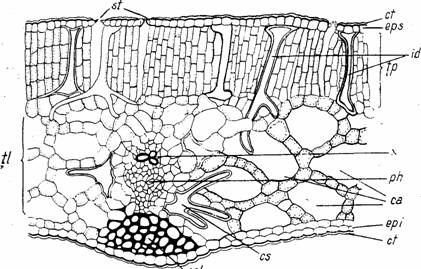
Fig. 9 - Structura unei frunze natante de Nymphaea alba (sectiune transversala): ca-canale aerifere; col-colenchim angular; cs-celula stelata; ct-cuticula; epi-epiderma inferioara; eps-epiderma superioara; id-idioblaste; ph-floem; st-stomate; tl-tesut lacunar; tp-tesut palisadic; x-xilem (d. Poplavskaia)
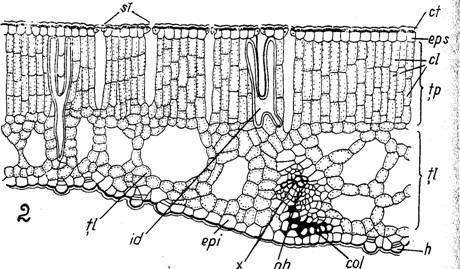
Fig. 10 - Structura unei frunze natante (2) de Nuphar luteum (sectiune transversale): ca-canale aerifere; cl-cloroplaste; col-colenchim angular; ct-cuticula; epi-epiderma inferioara; eps- epiderma superioara; h-hidropote; id-idioblaste; pa- parenchim aerifer; ph-floem; st-stomata; tl-tesut lacunar; tp-tesut palisadic; x-xilem (d. Poplavskaia)
Din punct de vedere structural, frunzele plantelor acvatice se caracterizeaza printr-o dezvoltare ampla a tesutului aerifer. Canalele aerifere care strabat frunza, inlesnesc atat schimbul de gaze cu mediul extern cat si ridicarea si mentinerea lor in stare de plutire mai aproape de lumina. Slaba reprezentare a tesutului conducator lemnos este o consecinta atat a reducerii transportului de apa, care difuzeaza prin toata suprafata corpului cat si a lipsei procesului de transpiratie.
La plantele care au si frunze submerse (Potamogeton lucens, Trapa natans, Nuphar lutea s.a.) mezofilul este omogen nediferentiat in tesut palisadic si lacunar. Intregul mezofil este strabatut din abundenta de canale aerifere mai mari sau mai mici. (Fig. 10.1; Fig. 11)
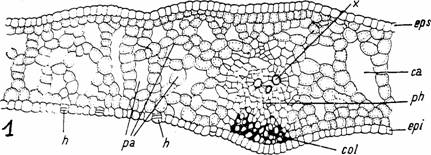
Fig. 10 - Structura unei frunze submerse (1) de Nuphar luteum (sectiune transversale): ca-canale aerifere; cl-cloroplaste; col-colenchim angular; ct-cuticula; epi-epiderma inferioara; eps- epiderma superioara; h-hidropote; id-idioblaste; pa- parenchim aerifer; ph-floem; st-stomata; tl-tesut lacunar; tp-tesut palisadic; x-xilem (d. Poplavskaia)
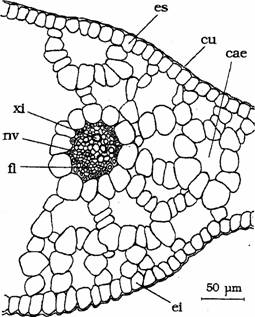
Fig. 11 - Structura frunzei de Potamogeton lucens (sectiune transversala): cae-canal aerifer; cu-cuticula; ei-epiderma inferioara; es-epiderma superioara; fl-floem; nv-nervura; xi-xilem (d. Sarbu,1999)
Aceste spatii intercelulare ale mezofilului se dezvolta de obicei schizogen, lizogen, iar la Typha sp.-rexigen.
La Elodea canadensis structura frunzei este reprezentata de 2 straturi de celule cu exceptia nervurii mediane.
Structura petiolului. Depinde in mare parte de simetria sa. La plantele cu petiol cilindric (Nuphar lutea, Nymphaea alba) si cu simetrie radiara structura este de tip monofacial.
Petiolul prezinta aceleasi particularitasi intalnite si la nivelul laminei: aerenchim bine dezvoltat, stereom (tesuturi mecanice) reprezentat in special prin sclereide (astrosclereide), tesut conducator lemnos redus etc (Fig.12; Fig.13).
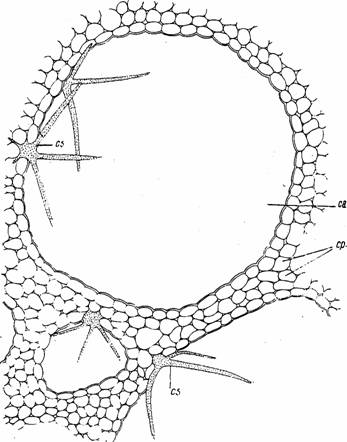
Fig. 12 - Parenchim aerifer la Nymphaea alba (sectiune transversala prin petiol): ca-canal aerifer; cp-celule parenchimatice; cs-celula stelata (d Tarnavschi & colab.)
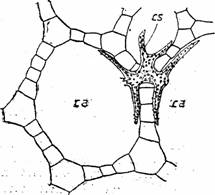
Fig. 13 - Parenchim aerifer la Nuphar luteum (sectiune transversala prin petiol): ca- canal aerifer; cs-celula stelata (d. Andrei,1976)
O adaptare specifica a plantelor acvatice si palustre consta in prezenta tesutului aerifer bine dezvoltat atat la nivelul organelor vegetative cat si reproducatoare.
Celulele acestui tesut lasa intre ele mari spatii sau lacune, de origine lizigena, care iau forma de canale mari ce strabat intregul corp al plantei respective. In aceste spatii se inmagazineaza mari cantitati de aer care determina micsorarea greutatii specifice a plantelor respective, facandu-le sa pluteasca sau sa se mentina in pozitie verticala.
Pe de alta parte aerul atmosferic din spatiile intercelulare constitue o rezerva interna din care planta utilizeaza oxigen pentru respiratie si dioxid de carbon pentru fotosinteza. Sistemul de spatii, respectiv canalele aerifere de la plantele acvatice, reprezinta o adaptare a acestora la conditiile mediului acvatic, unde gazele necesare proceselor metabolice fie ca nu pot fi absorbite, fie ca se gasesc in cantitati insuficente.
Canalele aerifere se formeaza in tesutul cortical la Myriophyllum sp., in tesutul medular la Juncus sp., sau in ambele tesuturi la Nymphaea alba. In raport cu presiunea exercitata de apa diferite specii de plante acvatice si-au creat adaptari caracteristice prin care isi mentin deschise (in stare normala) canalele aerifere: sclereide, druze, rafide, tesut diafragmatic.
Tesutul diafragmatic este alcatuit din celule cu pereti subtiri de forma neregulata, prevazut cu spatii intercelulare evidente. Celulele acestui tesut provin din proliferarea celulelor care marginesc canalul. In sectiune transversala el apare sub forma unei perdele prinsa de celule care delimiteaza canalul, din care se si diferentiaza.
Morfologia tesutului diafragmatic difera de la o specie la alta. Acest tip de tesut a fost pus in evidenta atat la plantele acvatice submerse (Potamogeton gramineus) cat si la cele cu frunze putitoare (Nuphar lutea, Nymphaea alba) si palustre (Juncus effusus) (Fig.14).
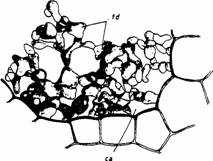
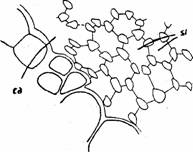
Fig. 14 - Tesut diafragmatic la Nymphaea alba (1) si Potamogeton gramineus (2) (sectiuni transversale prin petiol): ca - canal aerifer; si - spatiu intercelular; td - tesut diafragmatic (d. Andrei, 1976)
Sclereidele, numite si "celule stelate" provin din modificarea puternica a unor celule care delimiteaza canalul aerifer (Fig.12; Fig.13). Dupa forma care o au la diferite plante pot fi incadrate in mai multe tipuri. Sclereidele canalelor aerifere se sprijina unele pe altele, formand un sistem de sustinere eficient, care implica turtirea sau inchiderea acestora. Ele cresc in dimensiune si dobandesc o forma caracteristica de stea, peretii se ingroasa puternic cu substante minerale (carbonat de calciu) ceea ce conduce la piederea vitalitatii in conditiile edificarii unui sistem de sustinere eficient (Fig.15).

Fig. 15 - Sclereide din canalele aerifere ale petiolului de Nymphaea alba (sectiune longitudinala): ca-canal aerifer; sc-sclereide (d. Andrei, 1976)
Druzele si rafidele sunt cristale de oxalat de calciu, grupate in forma de cap de buzdugan (druze) sau aciculare (rafide). Ele se dezvolta in celulele parenchimului aerifer, celule care isi modifica forma si proemina spre lumenul canalului respectiv (Myriophyllum sp.).
CAPITOLUL 3
PARTICULARITATI ALE PROCESULUI DE REPRODUCERE
Plantele acvatice si palustre se reproduc atat sexuat cat si asexuat; predomina insa reproducerea asexuata prin multiplicare vegetativa.
In cadrul reproducerii sexuate intalnim plante anemofile, entomofile dar si numeroase hidrofile.
La Vallisneria spiralis, planta acvatica, dioica, florile feminine au un peduncul foarte lung, spiralat, care se desface astfel incat florile se ridica la suprafata apei dar raman ancorate. La suprafata apei florile feminine se deschid etaland cele trei stigmate (Fig.16: 1, 2). Florile masculine sunt grupate intr-un ament globulos invelit intr-o bractee. O floare masculina prezinta trei tepale si doua stamine. In timpul polenizarii florile masculine se desprind de pe axa inflorescentei si se ridica una cate una la suprafata apei (Fig. 16), aici se deschid anterele si florile masculine plutesc libere pana ce intalnesc florile feminine.
Florile masculine aduse de curentii apei intalnesc stigmatele florilor feminine realizandu-se astfel polenizarea (Fig.16: 3, 4, 5, 6). Ulterior florile feminine se inchid si se retrag treptat in apa datorita pedunculului florar spiralat.
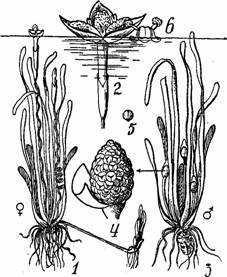
Fig. 16 - Polenizarea la Vallisneria spiralis: 1-planta feminina; 2-floare feminina marita; 3-planta masculina; 4-inflorescenta masculina; 5-o floare masculina desprinsa se ridica la suprafata apei; 6-floare masculina m momentul cand anterele ating stigmatul unei flori feminine (d. Grintescu, 1985)
Formarea si maturarea semintelor si a fructelor are loc in apa pe cand inflorirea si polenizarea au loc la suprafata apei.
La Ceratophyllum sp., planta acvatica, monoica, inflorirea si polenizarea se realizeaza in apa.
Reproducerea vegetativa predomina datorita temperaturilor scazute ale mediului acvatic. Modalitatile de multiplicare vegetativa sunt variate. La o serie de taxoni are loc fragmentarea tulpinii sau a ramurilor (Ceratophyllum sp., Myriophyllum sp., Hydrocharis morsus-ranae, Potamogeton sp., Utricularia sp., Salvinia natans). Numeroase specii palustre se multiplicaprin fragmentarea rizomului (Cyperus, Carex, Phagmites australis)-modalitate de multiplicare si propagare mult mai eficienta decat cea realizata prin seminte.
La unele plante acvatice (Utricularia sp., Strtiotes sp., Hydrocharis morsus ranae, Ceratophyllum) se formeaza toamna turioni, muguri tulpinali tuberizati in care se acumuleaza substante de rezerva, care ierneaza pe fundul baltilor, iar primavara genereaza noi indivizi (Fig.17).
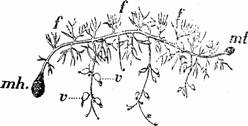
Fig. 17 - Utricularia vulgaris: f-frunze normale; mh-muguri hibernali; mt-mugure terminal; v-vezicule (d.Grintescu,1985)
Sagittaria sagittifolia trece peste perioada de iarna cu ajutorul tuberculilor formati la extremitatea stolonilor.
Cele mai multe plante acvatice ierneaza cu ajutorul mugurilor foliari terminali sau axilari hibernali numiti hibernacule, muguri potejati de frunze modificate, rezistente la temperaturi scazute (Hydrocharis morsus-ranae, Sagittaria sagittifolia, Stratiotes aloides, Potamogeton sp.). Mugurii hibernali sunt de forma rotunda sau ovala; ei pot fi terminali sau axilari; uneori acesti muguri se desprind de pe planta mama si ierneaza in namol (Potamogeton crispus); alteori cad pe fundul apei odata cu tulpina plantei mama si nu se desprind decat primavara cand incep sa "incolteasca".
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 11147
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved