| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
In dezvoltarea lor ontogenetica, plantele trec printr-o perioada cu activitate fiziologica intensa si printr-o perioada de viata latenta (repaus). In climatul temperat, starea de repaus se datoreste alternarii perioadelor calde cu perioadele reci. Repaus la plante, reprezinta rezultatul unui proces de adaptare la conditiile nefavorabile de mediu. Organele de crestere sunt mai putin rezistente la aceste conditii, in timp ce organele in repaus prezinta o mare rezistenta, astfel incat incetarea cresterii si intrarea in repaus in sezonul nefavorabil asigura supravietuirea. Repausul vegetativ, pentru care multi specialisti utilizeaza termenul de dormanta reprezinta o faza a cresterii ritmice a plantelor.
Starea de repaus se poate instala la planta intreaga, asa cum este cazul speciilor lemnoasesi al arbustilor, la unele organe subterane ale plantelor bienale si perene, care sunt prevazute cu muguri (bulbi, tuberculi, rizomi, tuberobulbi), sau la cele prevazute cu embrioni (seminte).
Pentru multe specii de plante repausul vegetativ este obligatoriu, pentru a putea trece la o noua faza de crestere vegetativa. De asemenea, unele seminte nu pot incolti daca nu au parcurs faza de repaus, chiar daca au asigurate conditiile optime pentru germinare.
In functie de perioada in care se instaleaza repausul vegetativ, Lang (1987) deosebeste:
- ectodormanta (repausul fortat) - cauzata de conditiile de mediu nefavorabile din timpul toamnei, care determina intrarea in repaus inainte de perioada caracteristica
- endodormanta (repausul propriu-zis) - catacterizata prin instalarea in
-ecodormanta (repausul secundar) - este determinata de actiunea factorilor nefavorabili din mediul inconjurator si se produce dupa ce plantele au intrat in vegetatie.
9.1. MECANISMUL FIZIOLOGIC AL REPAUSULUI VEGETATIV
Conceptia clasica privind mecanismul de inducere a repausului vegetativ sustine rolul prioritar al hormonilor. In anul 1968 Milborrow a descoperit acidul abscisic in seminte, iar in perioada care a urmat numerosi cercetatori au studiat efectul acestui hormon vegetal.
Galston s.a. (1980) au precizat rolul acidului abscisic ca hormon inhibitor de crestere si stimulator pentru intrarea in repaus vegetativ. Dupa Karsen si colab. (1983), acumularea acidului abscisic in embrion este factorul primar care determina intrarea in repausul seminal. Martin (1991) considera ca acidul abscisic este un factor secundar in repausul vegetativ al mugurilor, iar sfarsitul repausului nu este corelat cu continutul in acid abscisic.
Acidul abscisic se acumuleaza in tesuturile plantelor din zonele temperate expuse la fotoperioade scurte si la temperaturi coborate. Pe masura ce creste durata perioadei de expunere la temoeraturi coborate, continutul de acid abscisic scade. Astfel, continutul in acid abscisic al semintelor provenite de las pomii fructiferi scade ajungand la zero, dupa 3 saptamini de expunere la temperatura de 30C (Rudnicki, 1969).
Actiune sinergica cu aceste hormon au si unele substante fenolice ca: acidul ferulic, acidul vanilinic, acidul p-cumaric, cumarinele si etilena (Come, 1983).
Expunerea la temperaturi coborate a tesuturilor stimuleaza procesul de sinteza a auxinelor, giberelinelor si citochininelor, favorizand iesirea din repausul vegetativ. Acest efect a fost pus in evidenta de Luckwill (1952) la semintele de Malus stratificate si expuse la temperaturi coborate negative.
Tillberg (1984) a precizat ca auxina nu are un rol
regulator in repausul seminal la
Acidul giberelic scade cantitativ la inceputul perioadei de repaus mugural si creste la sfarsitul acestei perioade. Gianfanga si Rachmiel (1986) au constatat existenta unui proces de acumulare a giberelinelor in semintele expuse la temperaturi coborate, iar Hatch si Walker (1969) au precizat ca aplicarea exogena a giberelinelor poate inlocui necesarul de frig la mugurii de Armeniaca si Persica.
Citochininele stimuleaza cresterea mugurilor si germinarea semintelor. Dupa Davies (1988), rezultatele difera in functie de specie si cauza ar putea fi viteza diferita de transport a citochininelor prin seminte spre embrion.
Dupa Dilley (1969), influenta hormonilor se manifesta atat in urma actiunii lor individuale, cat si prin modificarea raportului dintre hormonii stimulatori si cei inhibitori de crestere.
In procesul de inducere a repausului vegetativ sunt implicate nucleotidele adenilate a caror sinteza este blocata pe parcursul acestei perioade. Nucleotidele reprezentate de uridin trifosfat (UTP), citozin trifosfat (CTP) si guanozin (GTP) sunt indispensabile pentru cresterea mugurilor, constituind forme de transport pentru glucide. O data cu iesirea din repausul vegetativ, sinteza acestor substante este deblocata (Come, 1992).
Scoala franceza de fiziologie considera ca repausul vegetativ este reglat genetic. Cercetarile efectuate de Purvis (1939) au precizat ca la Secale cereale, soiul Petkus, repausul este controlat de o gena, iar la Triticum si Hordeum (tetraploid) sunt implicate cel putin doua gene. Smith (1951) considera insa ca Hordeum este o singura gena de control a repausului vegetativ.
La specia Arabidopsis thaliana, controlul genetic al repausului mugural este asigurat de 4 gene (Napp-Zinn, 1969). Dupa Thompson s.a. (1985), repausul mugural al pomilor fructiferi este controlat de mai multe gene, cu exceptia genului Alnus la care s-a identificat o singura gena care controleaza acest proces. In cazul plantelor ierboase, Paulez si Perry (1987) au constatat existenta mai multor gene care controleaza repausul vegetativ.
Dupa Martin (1991), controlul genetic al repausului vegetativ este complex si consta in prelucrarea semnalelor din mediul inconjurator, din transformari moleculare si modificari celulare care determina in final transformari fiziologice. In prima etapa factorii ambianti determina expresia genelor repausului, care induc formarea unui ARN-m ce codifica biosinteza protein-enzimelor implicate in sinteza acidului abscisic, care inhiba procesele de crestere. Aceste procese pot decurge conform schemei:
Schema Burzo pag. 304
In cea de a doua etapa au loc modoficari morfologice si fiziologice, corelate cu diminuarea intensitatii tuturor reactiilor metabolice din tesuturile care intra in repaus.
9.2. REPAUSUL PLANTEI INTREGI
Starea de repaus vegetativ se instaleaza la toate organele perene ale plantelor lemnoase: radacini, tulpini si muguri. Intrarea in repaus nu se realizeaza concomitent in toate aceste organe. Astfel, repausul se instaleaza mai intai in muguri, apoi in trunchi si ramuri si, in final, in radacini (Martin, 1991).
Factorii principali care determina instalarea repausului vegetativ la plantele bienale si pere sunt reprezentati de temperaturile coborate din timpul noptii, diferentele mari de temperatura dintre zi si noapte, fotoperioadele scurte si umiditatea redusa a solului, in preioada de inceput a toamnei.
Fuchigami s.a. (1997) au constatat ca speciile lemnoase expuse la temperaturi coborate cateva saptamani, la inceputul toamnei, au un repaus profund. Intensitatea repausului difera: lastari varstnici ai pomilor au un repaus vegetativ mai profund comparativ cu cei tineri, mugurii vegetativi comparativ cu cei florali si tesuturile exterioare comparativ cu cele interioare. Intensitatea maxima a repausului vegetativ se manifesta la o saptamana pana la o luna, dupa caderea frunzelor.
Momentul de intrare in repaus vegetativ la speciile lemnoase este indicat de caderea frunzelor. Acest proces are influente deosebite asupra proceselor fiziologice din plante. Prin caderea frunzelor se anuleaza presiunea negativa generata de procesul de transpiratie, ceea ce conduce la diminuarea absorbtiei apei din sol si a ritmului de transport a sevei brute. Blocarea cu caloza a vaselor liberiene determina scaderea si mai evidenta a transportului substantelor prin plante.
Pierderea apei din tesuturile organelor perene determina concentrarea sucului celular si aparitia plasmolizei de iarna. Concentrarea sucului celular este determinata si de hidroliza amidonului, depozitat ca substanta de rezerva in organele perene si de acumularea glucidelor solubile rezultate din acsest proces (fig. 131). Concentrarea sucului celular determina cresterea rezistentei la inghet a tesuturilor plantelor. Astfel, tesuturile din Acer pseudoplatanus pot ingheta primavara, daca sunt expuse la temperaturi cuprinse intre -1,0 si -5,0 0C, in timp ce in perioada de repaus temperatura de inghet a tesuturilor scade la -12,0 .-50,00C.
Cresterea concentratiei sucului celular determina in acelasi timp si sporirea vascozitatii citoplasmei. Aceasta conduce la scaderea vitezei curentilor citoplasmatici si, ca urmare, a transporturilor intracelulare, cat si la reducerea activitatii enzimatice.
Din punct de vedere citologic, repausul vegetativ se caracterizeaza prin intreruperea plasmodesmelor (Romberger s.a., 1993), fapt ce contribuie la blocarea transportului intercelular pe cale simplasmica.
Pe parcursul perioadei de repaus are loc modificarea balantei hormonale, modificare determinata de scaderea lenta a continutului in acid abscisic din tesuturi si biosinteza hormonilor stimulatori de crestere.
Spre sfarsitul iernii, in tesuturile aflate in repaus vegetativ se observa o intensificare lenta a proceselor fiziologice si biochimice, corelata cu iesirea din repaus. Aceste modificari sunt mai evidente la inceputul primaverii, cand onditiile de mediu devin favorabile pentru desfasurarea proceselor fiziologice. Durata perioadei de repaus vegetativ variaza in functie de specie (fig. 132).
Primele modificari sunt legate de initierea absorbtiei apei in sol, hidroliza calozei din vasele liberiene si refacerea plasmodesmelor. Hidratarea tesuturilor favorizeaza intensificarea activitatii enzimatice a proceselui de respiratie si ca urmare a producerii de energie biochimica (ATP). Toate aceste modificari stimuleaza procesele de biosinteza in care sunt implicate enzimele existente in stare inactiva si ARN-m conservat in celulele aflate in stare de repaus. Primele reactii biochimice nu necesita biosinteza "de novo" a enzimelor, dar ulterior are loc intensificarea procesului de translatie si transcriptie si biosinteza "de novo" a enzimelor implicate in iesirea din repausul vegetativ.
Glucidele de rezerva sunt biodegradate, iar compusii intermediari rezultati sunt utilizati prin biosinteza substantelor necesare pentru cresterea mugurilor vegetativi si florali, precum si pentru producerea de energie biochimica in procesele de repiratie.
Formarea frunzelor asigura autonomia fata de substantele de rezerva, trecandu-se la nutritia autotrof fotosintetizanta.
9.3 REPAUSUL MUGURILOR
9.3.1. Caracterizarea morfo-fiziologica a mugurilor
Plantele perene isi asigura supravietuirea in sezonul nefavorabil prin formarea unor organe specializate numite muguri. Mugurii contin un tesut meristematic care reprezinta primordiile lastarului, invelit in solzi protectori, impermeabili pentru apa si gaze.
Din punct de vedere structural si functional mugurii sunt de doua categorii, si anume muguri vegetativi, din care vor rezulta lastari vegetativi si muguri reproducatori sau floriferi, din care vor rezulta flori.
Mugurii
vegetativi contin un tesut meristematic in care se formeaza
primordii de
acoperiti, prevazuti cu solzi protectori, la majoritatea plantelor;
nuzi, lipsiti de solzi protectori, de exemplu la Rhamnus frangula si Viburnum lantana (fig. 133
Dupa pozitia lor pe planta pot fi (Rivals, 1966
terminali sau apicali, ce apar in varful lastarului (fig. 134
axilari sau laterali, ce apar la noduri, in axila frunzelor;
adventivi, ce apar pe
organe care de obicei nu au muguri, de exemplu radacini si
dorminzi, ce contin un 'meristem de asteptare', care devine activ numai in conditii speciale, de exemplu taieri severe, inghet, etc.
Mugurii vegetativi contin primordiile lastarului vegetativ,
cu o structura metamerica, cu un numar definit de metamere (unitati de structura alcatuite din internod, care la
partea superioara are
Mugurii reproducatori sau floriferi dau nastere la flori. Din punct de vedere structural si functional pot fi muguri florali si micsti.
Mugurii florali sunt total sexualizati. Diferentierea florala cuprinde intregul meristem, iar meristemul vegetativ lipseste. Mugurii florali pot fi:
tip buton, in care se formeaza o singura floare, de exemplu la Forsythia, Rhododendron;
purtator de inflorescente, de exemplu la Syringa.
Mugurii micsti sunt partial sexualizati. Diferentierea florala cuprinde numai o parte din meristem, astfel incat mugurii contin atat primordii florale, cat si meristem vegetativ (fig. 135).
Dupa pozitia primordiilor florale pot fi:
cu primodrii florale terminale, care formeaza un lastar
cu numar definit de metamere, la care axul prezinta o crestere internodala. In
varf prezinta flori solitare, de exemplu la
cu primordii florale axilare, care formeaza un lastar cu varf vegetativ terminal si o crestere nelimitata. Primordiile florale sunt laterale, formand o floare sau o inflorescenta, de exemplu la vita de vie.
|
|
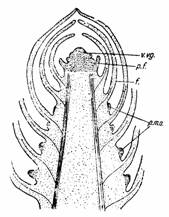
Fig. 134 Schema mugurului terminal: v.vg. -varful vegetativ; p.f. - primordii foliare; f. - p.m.a. - promordiile mugurilor axiali |
|
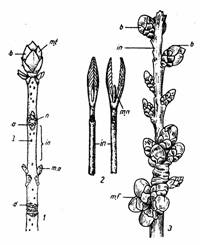
|
Fig. 133 Diferite tipuri de muguri: 1-muguri acoperiti la Aesculus hippocastanum; 2-muguri nuzi la Viburnum lantana; 3-muguri floriferi la Cerasus sp.); m.t. - mugure terminal; b - catafile; n - noduri; m.a. - muguri axilari (laterali); m.f. - muguri florali; in - internod; l - lenticela; m.n. - mugure nud; d - cicatricele frunzelor |
|
|
Fig. 135 - Schema unui mugure reproducator mixt cu primordii florale si meristem vegetativ la mar (dupa Abbott, 1970) |
Mugurii sunt caracteristici atat pentru tulpinile plantelor perene lemnoase, arbori sau arbusti, cat si pentru tulpinile subterane ale plantelor ierboase perene prin bulbi, tubero-bulbi, rizomi, etc.
Evolutia starii de repaus a mugurilor
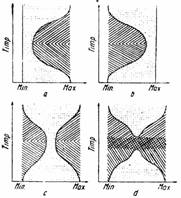
Vegis (1964) arata ca evolutia mugurilor spre starea de repaus se face treptat. In prima faza, numita repaus timpuriu mugurii sunt capabili de crestere, dar intr-o serie din ce in ce mai ingusta de conditii externe. Aceasta faza evolueaza spre repausul propriu-zis, in care mugurii sunt incapabili de crestere, indiferent de conditiile de mediu. In faza a treia, numita post-repaus sau repaus fortat, mugurii sunt capabili sa-si reia cresterea intr-o serie din ce in ce mai larga de conditii externe (fig. 136).
|
|
Fig. 136 - Tipuri de repaus la muguri: a - ingustarea domeniului de temperaturi de crestere in zona temperaturilor scazute; b - in zona temperaturilor ridicate; c - in zona temperaturilor mijlocii; d - intercalarea unui repaus absolut la mijlocul perioadei de repaus. |
Evolutia starii de repaus a mugurilor poate fi apreciata urmarind modificarea capacitatii de pornire a mugurilor in timpul repausului, exprimata prin energia de inmugurire, respectiv procentul de muguri porniti in 15 sau 30 de zile.
Repausul mugurilor vegetativi este mai profund decat cel al mugurilor reproducatori. Desi la intrarea in repaus diferentierea primordiilor florale (sepale, petale, androceu, gineceu) a avut loc, in mugurii reproducatori in timpul repausului au loc modificari metabolice care desavarsesc maturizarea elementelor sexuale.
9.3.3. Influenta factorilor externi asupra repausului mugurilor
Temperatura. Influenta temperaturilor ridicate asupra aparitiei starii de repaus. O serie de ceretari arata ca intrarea mugurilor in repaus se face sub actiunea temperaturilor ridicate. Vegis (1964) deosebeste trei tipuri de repaus.
Primul tip se caracterizeaza prin incapacitatea mugurilor de a creste in conditii de temperatura ridicata. Cauza intrarii mugurilor in repaus este o temperatura prea ridicata pentru cresterea celulelor. Aceasta reactie isi are originea in climatul cald, unde serveste la supravietuirea plantelor la conditiile de caldura si uscaciune din diferite perioade ale anului.
Al doilea tip se caracterizeaza prin capacitatea de a creste la temperaturi din ce in ce mai ridicate. Prin acest tip este impiedicata cresterea toamna, cand temperatura este scazuta. In zonele temperate aceste conditii se manifesta in a doua jumatate a verii. Extinderea repausului si in perioadele reci ale anului este un fenomen de natura secundara si apare atunci cand starea de repaus nu s-a sfarsit la aparitia perioadei reci.
Al treilea tip se caracterizeaza prin cresterea mugurilor numai la temperaturi mijlocii. Acest tip este o adaptare la vara secetoasa si iarna aspra. Este intalnit la majoritatea speciilor lemnoase.
Influenta
temperaturilor scazute asupra intreruperii starii de repaus. Diferite cercetari
arata ca temperaturile ridicate din timpul iernii exercita o actiune de intarziere
a pornirii mugurilor (Samish, 1960). Iesirea
mugurilor din repaus se face numai dupa ce acestia au fost supusi la o anumita
perioada de temperaturi scazute, numita cerinta
de frig. Cerinta de frig a fost stabilita pentru diferite specii lemnoase,
de exemplu molid (Mienstaedt, 1966),
Temperatura scazuta determina modificari metabolice in muguri. La lalele are loc intensificarea activitatii enzimei amilaza, care hidrolizeaza amidonul si mareste continutul de zaharuri solubile in bulbi (Ohyama si colab., 1996). La Aster scaber are loc o scadere a continutului de acid abscisic (Seong, 1996), iar la Fritillaria pallidiflora are loc o crestere a numarului de ribozomi si o intensificare a sintezei ARN (Liu si Wang, 1996
Fotoperioada. Influenta fotoperioadei asupra intrarii mugurilor in repaus. Fotoperioada este unul din factorii cei mai importanti care controleaza intrarea mugurilor in repaus. La majoritatea plantelor lemnoase, conditiile de zi lunga din timpul verii determina cresterea vegetativa, iar conditiile de zi scurta de la inceputul toamnei determina incetarea cresterii si formarea mugurilor de repaus. Wareing (1969) arata ca desi in natura arborii isi opresc cresterea in lunile iunie-iulie, mugurii formati in aceasta perioada se afla in repaus timpuriu. Intrarea in repausul propriu-zis, se face toamna in conditii de zi scurta. La plantele lemnoase fotoperioada scurta mareste rezistenta la ger, iar fotoperioada lunga reduce aceasta insusire (Irving si Lamphear, 1967
Influenta
fotoperioadei asupra iesirii mugurilor din repaus Actiunea fotoperioadei scurte de a inceta
cresterea si de a forma muguri in repaus este perceputa de
Desi iesirea din repaus este conditionata de expunerea la temperaturi scazute, mugurii neexpusi la frig pot incepe cresterea prin expunere la fotoperioada lunga. Aceste rezultate au fost obtinute de Down si Borthwick (1965).
Fotoperioada scurta determina o crestere a
inhibitorilor din
9.3.4. Influenta factorilor interni asupra repausului mugurilor
Starea fizica a protoplasmei. Prin intrarea mugurilor in repaus, protoplasma trece de la starea de sol la starea de gel. In celule scade continutul de apa libera si creste continutul de apa legata, ceea ce blocheaza enzimele si micsoreaza permeabilitatea pentru diferite substante si oxigen.
Scaderea continutului de apa in muguri in timpul starii de repaus a fost semnalata la mar (Suzdalceva, 1970, Voican, 1970), par si gutui (Toma, 1975) si vita de vie (Cojeneanu si colab., 1971
Modificarea permeabilitatii mugurilor de par si gutui pentru diferite elemente minerale in legatura cu starea de repaus a fost pusa in evidenta prin metoda trasorilor radioactivi, folosind P32, S35, Ca45 si Zn65 (Toma, 1976
Iesirea mugurilor din repaus determina trecerea de la starea de gel la starea de sol si cresterea permeabilitatii protoplasmei, fenomen ce se realizeaza sub influenta auxinelor (Samish, 1954
Respiratia. In timpul repausului are loc o reducere a respiratiei. Solzii mugurilor, impregnati cu substante grase, hidrofobe, impiedica patrunderea oxigenului. Intreruperea repausului la Begonia este insotita de o intensificare a respiratiei (Esashi si Nagao, 1973
In timpul starii de repaus la mugurii de cais este inregistrata o scadere a activitatii enzimelor respiratiei, catalaza si ascorbinoxidaza. Activitatea scazuta a catalazei este corelata cu intensitatea scazuta a respiratiei mugurilor, iar activitatea scazuta a ascorbinoxidazei permite acumularea acidului ascorbic redus, cu rol in rezistenta la inghet.
Continutul de glucide. In timpul repausului continutul de zaharuri solubile creste, fiind maxim in timpul lunilor cu temperaturile cele mai scazute, ceea ce favorizeaza rezistenta la ger a plantelor.
Formarea zaharurilor solubile in timpul starii de repaus se realizeaza pe baza amidonului depozitat in ramuri si muguri in timpul toamnei. Acest continut este cunoscut sub numele de 'maximul de toamna' (Priestley, 1970) si este hidrolizat in timpul lunilor de iarna, fiind corelat cu rezistenta la inghet a plantelor pomicole. Acumularea de zaharuri solubile a marit rezistenta la inghet la cais, piersic si migdal (Molnar, 1969). La grapefruit, in timpul inghetului zaharurile solubile au crescut cu 25-50% (Young si Peynada, 1971
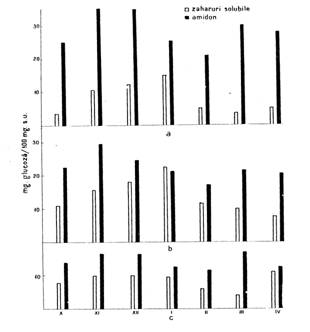
Dintre formele de zaharuri solubile acumulate in muguri in timpul repausului fac parte zaharoza la vita de vie, glucoza si mai putin fructoza la par si gutui (fig. 137
|
|
|
Fig. 137 - Dinamica formelor de glucide in timpul starii de repaus in ramura vegetativa la par; a - lemnul si maduva axului; b - scoarta si liberul axului; c - muguri vegetativi |
Continutul de lipide creste in timpul repausului, ceea ce micsoreaza hidratarea si permeabilitatea protoplasmei. Formarea acizilor grasi si a grasimilor este favorizata de reducerea absorbtiei oxigenului prin prezenta solzilor (Pouget, 1961
Compusii cu azot. In privinta azotului total, rezultatele experimentale sunt diferite. Uneori are loc o scadere a acestuia in timpul repausului, alteori continutul de azot total nu se modifica (Hemberg, 1963). Continutul de aminoacizi creste in relatie directa cu rezistenta la ger si profunzimea repausului la Picea glauca (Durzan, 1968) si migdal (Kazaryan si Karapetyan, 1962
La par si gutui, in timpul repausului au fost identificati arginina, acidul asparagic, acidul glutamic, serina si asparagina, iar la mar, acidul glutamic, asparagina si prolina. Prolina determina rezistenta la inghet si profunzimea repausului, iar arginina si asparagina sunt principalele forme de depozitare a azotului la plantele pomicole din familia Rosaceae (Tromp, 1970
Sabinin (cit. de Vegis, 1964) a sugerat ideea ca la arbori cresterea ritmica este datorata modificarii in intensitate a sintezei acizilor nucleici. In muguri, in timpul perioadei de repaus are loc o scadere a continutului de acizi nucleici, pe cand la iesirea din repaus cantitatea acestora creste (Vegis, 1964 Tuan si Bonner (1964) presupun ca repausul este controlat prin reglarea sintezei acizilor nucleici. In tuberculii de cartof in repaus are loc represiunea activitatii genetice, blocarea ADN incapabil sa suporte sinteza ARN mesager cu rol in sinteza proteinelor.
Stimulatorii de crestere In timpul repausului continutul de auxine din muguri este redus sau lipseste, dar creste la intreruperea repausului.
La mugurii de pin auxina inregistreaza o dinamica
sezoniera in acest sens (Alden, 1967). Continutul de
gibereline este minim in timpul repausului si creste la intreruperea repausului
mugurilor de Betula pubescens (Eagles si Wareing, 1963), Salix viminalis (Michniwics si colab., 1970) si alte specii
lemnoase (Netien, 1969). Continutul de
gibereline creste in lastarul florifer de lalea dupa iesirea din repaus (Rebers si colab., 1995). La Populus robusta (Hewet si Wareing, 1973) continutul de
citochinine este foarte scazut in timpul repausului mugurilor si creste la intreruperea
acestuia. La
Inhibitorii de crestere. La diferite specii lemnoase in timpul repausului mugurilor au fost puse in evidenta diferite substante inhibitoare cu caracter acid, care au fost denumite inhibitor beta. Libbert (1964) arata ca acesta actioneaza antagonist hormonilor stimulatori. In 1964, in mugurii de repaus de la diferite plante lemnoase a fost descoperita dormina, care prin identificare cu abscisina II, a primit denumirea de acid abscisic.
Acidul abscisic se acumuleaza in muguri in timpul perioadei de repaus manifestand o actiune competitiva stimulatorilor de crestere, prin inhibarea sintezei acizilor nucleici, in special a ARN mesager, cu rol in sinteza proteinelor. Continutul de acid abscisic scade in muguri la iesirea din repaus.
Cercetari efectuate prin metoda biotestelor cu testul coleoptilului de Avena au aratat ca la Gladiolus x tubergenii, iesirea mugurilor din repaus a determinat o crestere a continutului de stimulatori si o scadere a continutului de inhibitori in tubero-bulbi.
9.3.5. Intreruperea starii de repaus a mugurilor
Repausul fortat, datorat unor conditii externe nefavorabile (temperatura, umiditate) poate fi intrerupt prin expunerea la conditii favorabile cresterii. Ramuri de Forsythia suspensa aduse la temperatura camerei in luna martie infloresc cu cateva saptamani mai devreme decat cele din camp. Repausul propriu-zis, datorat unor factori interni, de natura fiziologica, nu poate fi intrerupt prin crearea unor conditii favorabile cresterii, ci numai prin operatia de fortare.
Fortarea reprezinta un ansamblu de tehnici de intrerupere a repausului propriu-zis, care au la baza mecanisme fiziologice. Rezultatul practic consta in grabirea pornirii in vegetatie si a infloririi plantelor. Metoda este deosebit de utila in practica floricola, deoarece realizeaza completarea sortimentului de flori in sezonul nefavorabil (decembrie-februarie), prin tipul de cultura fortata.
Tratamente culturale. Desicatia sau deshidratarea partiala este aplicata la Syringa si Calla. Metoda are la baza reactia ecofiziologica a plantelor la care in zonele de origine, conditiile nefavorabile ce determina repausul sunt reprezentate de seceta. Desicatia grabeste caderea frunzelor, maturarea mugurilor si a lemnului, scade continutul de stimulatori si grabeste acumularea acidului abscisic.
Defolierea sau indepartarea mecanica a frunzelor este aplicata la Hydrangea si grabeste intrarea in repaus si desfasurarea fazelor repausului.
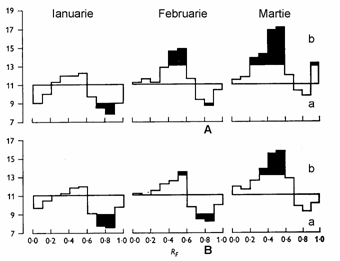
Tratamente fizice. Tratamentul cu frig are la baza cerinta de frig a mugurilor in general, pentru intreruperea repausului si a mugurilor reproducatori in special, pentru maturizarea elementelor sexuale. Temperatura scazuta determina scaderea continutului de inhibitori, de exemplu acidul abscisic si cresterea continutului de stimulatori endogeni in muguri (fig. 138). La lalele tratamentul cu frig la 50-170C, timp de 12 saptamani a produs o crestere rapida a lastarilor floriferi si un continut de gibereline de 2-3 ori mai mare (Rebers si colab., 1995
|
|
|
Fig. 138 - Efectul tratamentului cu frig asupra continutului de inhibitori (a) si gibereline (b) endogene in mugurii de Ribes nigrum stabilit cu ajutorul biotestului de porumb pitic. A - tratament cu frig 20C in luna ianuarie; B - martor in conditii naturale (dupa El Antably, 1965) |
Conditiile de racire, respectiv cerinta de frig la plantele floricole dureaza de la cateva saptamani la 1-2 luni, la temperaturi cuprinse intre -50 - -60C pana la 100 - 120C. Astfel, la Tulipa este necesara o racire la 80- 90C timp de 5-6 saptamani, la Convalaria -10 - -30C timp de 3-4 saptamani, iar la Syringa si Forsythia -50 - -60C timp de 5 - 6 saptamani (fig. 139). La Freesia tratamentul cu frig la 120C a grabit inflorirea cu 20 zile. Prin aceasta metoda, in conditii artificiale plantele intra in vegetatie si infloresc in epoci cu totul diferite fata de natura.
|
|
Fig. 139 - Intreruperea repausului hibernal la mugurii de Syringa vulgaris (dupa Kimball) |
Baia calda se aplica la ramuri detasate, prin scufundarea in apa la 300- 400C, timp de 3-12 ore. Efectul tratamentului este local si se datoreste creerii unor conditii de anaerobioza care declanseaza intreruperea repausului prin scaderea continutului de acid abscisic (Pouget, 1969). Efectul tratamentului se manifesta numai in timpul repausului propriu-zis si anume in noiembrie-decembrie la Syringa si Convalaria si in noiembrie-februarie la Forsythia.
Tratamente chimice. Eterul sau cloroformul actioneaza asemanator frigului si sunt eficace pe ramuri si muguri. Au actiune locala, determinand scaderea temperaturii. Se aplica la Syringa, Convalaria, Forsythia prin expunere la vapori de eter 48 de ore la temperatura de 180 - 200C. Alcoolul etilic a stimulat pornirea mugurilor la ramuri de Syringa vulgaris si Ligustrum vulgare care au fost scufundate cu treimea bazala timp de 15 minute (Yang si Read, 1996
Tratamente hormonale. Tratamentul cu giberelina GA3 10 sau 40 ppm urmat de tratamentul cu frig de 45 de zile la 120C a intrerupt repausul mugurilor si a grabit inflorirea la Freesia (Shi si colab., 1995), iar tratamentul cu giberelina 200 ppm a determinat intreruperea repausului mugurilor la Lilium japonicom (Kawarabayashi, 1995
9.4 REPAUSUL SEMINAL
.1. Structura semintei
Caracteristicile repausului seminal
Semintele apar in urma procesului de fecundare din transformarea ovulului. Formarea semintelor incheie ciclul vital al plantelor, asigurand inmultirea sexuata. Partile componente ale semintei sunt embrionul, endospermul si tegumentul.
Embrionul este alcatuit din tesut meristematic ce contine primordiile organelor vegetative ale plantei: radicula, tigela, gemula si cotiledoanele. Endospermul sau albumenul este un tesut de depozitare a substantelor de rezerva, iar tegumentul este invelisul protector al semintei. Semintele cu aceasta structura se numesc albuminate.
Prin procesul de germinatie, embrionul trece de la viata latenta la viata activa. La 25% din speciile vegetale, semintele sunt capabile sa germineze in conditii favorabile, imediat dupa recoltare. La celelalte specii, dupa recoltare semintele trec printr-o perioada de repaus.
Repausul seminal poate fi propriu-zis, biologic, provocat de cauze fiziologice, care nu poate fi intrerupt de conditiile externe favorabile si fortat, provocat de conditiile externe nefavorabile, care poate fi intrerupt cand acestea devin favorabile. Aparitia si caracteristicile repausului seminal sunt dictate de actiunea conditiilor de mediu din zonele ecologice de origine ale acestora, imprimate genetic.
Semintele multor specii intra in repaus vegetativ inca din timpul formarii lor. Dupa Hilhorst si Downie (1995), acest repaus primar reprezinta o strategie pentru supravetuirea in conditii nefavorabile.
Intensitatea procesului de respiratie al semintelor aflate in starea de repaus vegetativ este mult mai scazut comparativ cu a celorlalte organe ale plantelor, creand posibilitatea pastrarii acestora o perioada lunga de timp.
Cercetarile efectuate de autori au stabilit urmatoarea variatie a valorilor intensitatii procesului de repiratie a semintelor;
|
Ardei |
1,41 mg CO2/kg/h |
Morocv |
0,54 mg CO2/kg/h |
|
Ceapa |
- " - |
Ridichi de luna |
- " - |
|
Dovlecei |
-" - |
Spanac |
- " - |
|
Fasole |
- " - |
Tomate |
- " - |
|
Grau |
-" - |
Varza de toamna |
- " - |
|
Porumb |
- " - |
Vinete |
- " - |
|
Telina |
- " - |
Gulii |
- " - |
|
Leustean |
- " - |
Repausul seminal este initiat spre sfarsitul fazei de crestere a semintelor. Incetarea aprovizionarii cu apa si cu substante asimilate a semintelor, cat si deshidratarea acestora genereaza conditii de stres care determina represarea unor gene specifice si blocarea sintezei proteinelor si a acizilor nucleici.
In tegumentul seminal au fost identificate cumarine (Evenari, 1949) si fenoli (Murphy si Noland, 1981), care limiteaza accesul oxigenului spre embrion, reducand oxidarile celulare si producerea de energie pentru procesul de crestere. Aceste substante pot fi indepartate prin imersia semintelor in apa.
Substantele inhibitoare de crestere care induc repausul seminal se gasesc atat in tegument, cat si in embrion si se biosintetizeaza in perioada ce precede intrarea in repausul vegetativ.
Denis s.a. (1978) au constatat existenta in seminte a unui complex inhibitor alcatuit din mai multe substante printre care se pot mentiona unii compusi fenolici si acidul abscisic. Acest complex inhibitor se biodegradeaza pe parcursul perioadei de repaus (Kefeli si Kodyrov, 1971).
Acidul abscisic, principala substanta inhibitoare de crestere a fost identificata in semintele a numeroase specii de plante din genurile: Pisum, Juglans, Fraxinus, Malus, Acer, etc.
Aceasta substanta hormonala a fost identificata in: tegument, cotiledoane, embrion (Enu-Kwesi si Dumbroff, 1977). Dinamica continutului in acid abscisic din semintele de Malus este prezentata in figura 140.
Alaturi de substantele inhibitoare de crestere, in seminte au fost identificati si hormoni stimulatori de crestere. Nikolaeva (1967) a determinat prezenta in semintele de Acer tataricum a auxinei, alaturi de acidul abscisic. Continutul in auxina endogena activa scade treptat pe parcursul intrarii in repaus a semintelor.
Lewak (1985) a precizat ca giberelinele si citochininele din seminte au o concentratie scazuta pe parcursul perioadei de repaus, iar Amen (1986) considera ca semintele se afla in starea de repaus vegetativ nu numai din cauza prezentei substantelor inhibitoare, ci si din lipsa substantelor stimulatoare de crestere.
Continutul in substante stimulatoare si inhibitoare de crestere variaza in diferite parti ale semintelor. Acest raport poate regla translocarea hormonilor, biosinteza acestora sau transformarea formelor inactive (conjugate) in forme active (Lewak, 1985).
Existenta simulatana in seminte a substantelor stimulatoare si inhibitoare a repausului seminal face posibila existenta urmatoarelor situatii.
|
Starea semintei |
Gibereline |
Citochinine |
Acid abscisic |
|
Germinare |
|||
|
Germinare |
|||
|
Germinare |
|||
|
Repaus |
|||
|
Repaus |
|||
|
Repaus |
|||
|
Repaus |
|
||
|
Repaus |
("+" - prezenta hormonului; "-" absenta hormonului)
Dupa Bewley si Black (1982), exista doua tipuri de mecanisme care induc repausul seminal: repausul indus de embrion si cel indus de tegument.
9.4.2. Influenta factorilor externi asupra repausului seminal
Temperatura. Semintele diferitelor specii de plante isi intrerup repausul seminal numai dupa ce au fost supuse un anumit timp la temperaturi scazute.
Semintele de mar, piersic si maces germineaza la temperatura de 200C, numai daca au fost expuse anterior la o temperatura de 0-50C. Perioada de frig necesara intreruperii repausului este de 1-6 luni.
Expunerea la frig este necesara pentru intreruperea repausului la diferite plante floricole, de exemplu, Bellis, Primula, Salvia, Viola sau la diferite specii lemnoase (tab. 23
Tabelul 23
Specii lemnoase care necesita frigul pentru
intreruperea repausului seminal
(dupa Wareing, din Peterfi si Salageanu, 1972)
|
Acer saccharinum |
Juglans nigra |
|
A. pseudoplatanus |
Liriodendron tulipifera |
|
Betula sp. |
Picea sp. |
|
Cornus florida |
Pinus sp. |
|
Corylus avellana |
Ribes glossularia |
|
Crataegus sp. |
Rosa sp. |
|
Cupressus macrocarpa |
Sequoia gigantea |
|
Fagus silvatica |
Tilia sp. |
|
Fraxinus excelsior |
Thuja occidentalis |
Temperatura scazuta este asociata cu conditii de umiditate. Acest tip de repaus este numit psihrolabil.
Alte specii necesita temperaturi ridicate. Astfel, la Magnolia dealbata, repausul semintelor a fost intrerupt de o baie calda, timp de 24 de ore la 400C (Vovides si Iglesias, 1996), iar la Lilium x formolongi, expunerea la 140C pentru intreruperea repausului a necesitat un pretratament anterior de 290C timp de o saptamana - (Roh si Sim Yong Gu, 1996
Lumina. Starea de repaus a semintelor la diferite specii floricole este influentata de lumina, repausul fiind fotosensibil, pozitiv sau negativ (tab. 24
Repausul fotosensibil pozitiv se numeste fotolabil. Intreruperea repausului
seminal se face numai la lumina la unele specii de legume, cum sunt salata si
tomatele, la unele specii floricole, cum sunt Petunia hybrida si Salvia
splendens (Sonea si colab.,
1979) si la unele specii de buruieni cum sunt Amaranthus retroflexus, Capsella bursa-pastoris, Echium vulgare. La
Tabelul 24
Specii fotosensibile
(dupa Wareing din Peterfi si Salageanu, 1972)
|
Plante la care lumina stimuleaza germinatia |
|
|
Betula sp. |
Kalanchoe blossfeldiana |
|
Bidens tripartitus |
Paeonia sp. |
|
Digitalis purpurea |
Perilla ocymoides |
|
Escholtzia sp. |
Primula sp |
|
Plante la care lumina inhiba germinatia |
|
|
Helleborus niger |
Nigella damascena |
|
Nemophila insignis |
Phacelia tanacetifolia |
Repausul fotosensibil negativ se numeste scotofil. Intreruperea repausului seminal se face numai la intuneric la Freesia, Phlox, Delphinium (Sonea si colab., 1979
Majoritatea speciilor (65%) au fotosensibilitate pozitiva, numai 25% au o fotosensibilitate negativa, iar restul sunt lipsite de fotosensibilitate.
Mecanismul de actiune a luminii asupra intreruperii repausului seminal se explica prin participarea fitocromului, pigmentului de culoare rosu-deschis, cu absorbtie maxima in lungimea de unda 660 nm (P660) care trece sensibil intr-o forma de culoare rosu-inchis, cu absorbtia maxima in lumgine de unda de 730 nm (P730). P660 stimuleaza intreruperea repausului la lumina, in timp ce P730 stimuleaza intreruperea repausului la intuneric.
Umiditatea. La Impatiens balsamina si salata, intreruperea repausului seminal are loc prin expunere la uscaciune - repaus xerolabil. Actiunea temperaturilor scazute in intreruperea repausului necesita umiditate ridicata in sol.
9.4.3. Influenta factorilor interni asupra repausului seminal
Natura si structura tegumentului seminal imprima impermeabilitatea pentru apa la Robinia pseudacacia, Juniperus virginiana si teiul american sau pentru oxigen la seminte de Pinus strobus si frasin american. Dintre plantele floricole, diferite specii au seminte cu tegumentul dur: Canna, Cyclamen, Pelargonium.
Duritatea si impermeabilitatea tegumentului seminal pentru apa reprezinta o insusire genetica, care poate fi influentata si de conditiile climatice din timpul maturatiei semintei.
La plantele de cultura anuale, duritatea tegumentului seminal este redusa, ceea ce favorizeaza o germinatie si o rasarire uniforma. La culturile furajere duritatea tegumentului seminal este ridicata, ceea ce favorizeaza autoreinsamantarea acestora, de exemplu la Trifolium resupinatum si T. aureum.
Formarea tegumentului seminal dur este favorizata de umiditatea redusa in sol la lucerna sau de umiditatea redusa si temperatura ridicata la soia, in timpul maturarii semintelor.
Duritatea tegumentului opune o rezistenta mecanica pentru cresterea embrionului si determina impermeabilitatea pentru gaze, in special O2. Germinatia semintelor la majoritatea plantelor agricole este inhibata de absenta O2, cu exceptia orezului.
Starea fiziologica a embrionului. La diferite specii, embrionul provenit in urma fecundatiei nu este complet matur, ci necesita o perioada de post-maturatie. Fenomenul este intalnite si la semintele de frasin si pin.
In timpul post-maturatiei au loc procese metabolice calitative. Starea de post-maturatie corespunde repausului propriu-zis al embrionului. In aceasta etapa, activitatea metabolica este incetinita ca urmare a reducerii respiratiei aerobe si a incetinirii activitatii enzimatice. Are loc scaderea continutului de acizi nucleici, a capacitatii ADN de a suporta sinteza ARN mesager cu rol in sinteza proteinelor, scaderea continutului de stimulatori endogeni si acumularea acidului abscisic.
Substantele inhibitoare din seminte sau fructe au fost puse in evidenta la diferite specii. Natura inhibitorilor este diferita, si anume pot fi acizi organici, de exemplu acid malic, acid tartric, acid citric in pulpa fructelor de mar, struguri, citrice, glicozizi cianogenetici, de exemplu amigdalina la seminte de prun si migdal alcaloizi de exemplu nicotina, cofeina si cocaina in semintele de tutun, cafea si cacao, substante volatile din fructe si seminte, de exemplu din fructele de citrice sau semintele de umbelifere.
9.4.4. Intreruperea repausului seminal
La majoritatea speciilor cultivate, semintele germineaza usor si uniform. La unele specii se aplica diferite tratamente care au ca scop intreruperea repausului embrionar, inlaturarea barierei mecanice a tegumentului sau a substantelor inhibitoare din seminte sau fruct.
Stratificarea se face prin expunerea semintelor cu tegument dur la temperatura de 10-100C si umiditate, timp de 50-90 de zile, functie de specie. Stratificarea satisface cerinta de frig necesara intreruperii repausului. Este folosita la semintele de piersic.
Vernalizarea se face prin imbibarea semintelor la temperatura optima pana la inceputul germinatiei, expunerea acestora timp de 10-14 zile la temperaturi joase de 20-50C urmata de expunerea la temperatura optima de germinare. Racirea timp de 14 sau 28 de zile stimuleaza germinatia la Dianthus superbus, (Sim si colab., 1996 Vernalizarea satisface de asemenea, cerinta de frig pentru intreruperea repausului si este folosita la Festuca sp.
Tratamentele hormonale sau hormonizarea consta in imbibarea semintelor cu substante stimulatoare de tipul auxinelor, giberelinelor etc. Tratamentul cu GA a stimulat germinatia semintelor la salata si ovaz (Gradner si colab., 1994), iar tratamentul cu giberelina in concentratie de 0,02 %, timp de 12 ore a stimulat germinatia semintelor la garoafe si petunii. La Primula vulgaris, tratamentul cu giberelina a sporit efectul stratificarii (Benne, 1996
Imbibarea se aplica semintelor cu tegumentul dur. Imbibarea consta in mentinerea semintelor in apa la temperatura de 250- 350C, timp de 24-48 de ore, pana cand tegumentul crapa. Imbibarea favorizeaza gelificarea substantelor pectice din tegument. Se aplica la semintele de Pelargonium si Freesia.
Tratarea hidrotermica se aplica semintelor cu tegumentul foarte dur si consta in expunerea semintelor alternativ, timp de 2-3 min. la apa clocotita si gheata, de 2-3 ori. Se aplica la Canna.
Degradarea mecanica a tegumentului se aplica la seminte cu tegumentul foarte dur. Se face prin pilire, frecare cu nisip sau pietris sau crestare cu un cutit. Se aplica la semintele de Canna, palmieri si lucerna.
Tratamentele cu substante chimice se aplica de asemenea, la seminte cu tegumentul foarte dur sau impregnat cu rasini. Se folosesc tratamente cu acid sulfuric, acid clorhidric sau diferite saruri. La cactusii Echinocactus grussonii si E. platyacanthus, germinatia este stimulata cu acid sulfuric concentrat (De la Rosa si colab., 1994), iar la Tagetes erecta, cu azotat de potasiu 0,75% (Selvaraju si Selvaraju, 1994
Indepartarea pericarpului. Germinatia semintelor la Zinnia violacea a fost stimulata prin indepartarea pericarpului (Miyaima, 1996
9.4.5. Germinarea semintelor
Germinarea semintelor reprezinta procesul prinn care acestea initiaza un nou ciclu de dezvoltare ontogenetica, cand sunt asigurate conditii favorabile de mediu.
Din punct de vedere agronomic, se considera ca samanta a germinat cand da nastere la o noua plantula care se poate hrani autotrof. Din punct de vedere fiziologic, se considera ca procesul de germinare este incheiat cand radicula a penetrat prin fisurile tegumentului in sol (Mazliak, 1982).
Principalii parametri fiziologici ai semintelor prin intermediul carora se poate aprecia procesul de germinare a semintelor sunt urmatorii:
- longevitatea, care reprezinta durata de viata a semintelor pastrate in conditii normale. Acest parametru variaza in functie de intensitatea metabolismului, structura semintei si canitatea de substante de rezerva;
- facultatea germinnativa reprezinta procentul de seminte care germineaza in conditii optime pentru specia respectiva;
- viabilitatea reprezinta insusirea embrionului de a-si mentine reactiile caracteristice pentru tesuuturile vii (testul cu saruri de tetrazoliu).
Iesirea semintelor dinn repaus vegetativ se realizeaza in conditii favorabile de mediu ( temperatura, umiditate, lumina) care determina activitatea genelor implicate in biosinteza ADN-ului, respectiv ARN-m care codifica enzimele ce catalizeaza reactiile caracteristice pentru acest proces: biosinteza hormonilor stimulatori si ai enzimelor (Mazliak, 1982).
Germinarea se manifesta numai la semintele mature, care au iesit din starea de repaus. Semintele cu embrionul imatur se remarca prin dimensiunile mici ale acestuia si inainte de germinare embrionul trebuie sa creasca si sa se dezvolte. Bewley si Black (1982) mentioneaza ca unele specii se caracterizeaza prin prezenta unor embrioni morfologic imaturi ca: Anemone coronaria, Fraxinnus excelsior, Fraxinus nigra, Pinus sylvestris, Ranunculus biternatus, etc.
Maturarea semintelor de Fraxinus nigra se poate realiza prin expunerea semintelor timp de 18 saptamani la temperatura de 20-300 C urmata de pastrarea timp de 2-4 luni la temperatura de 5-100C (Vanstone si le Croix, 1975).
Semintele pomilor nu germinneaza inainte de a fi postmaturate. Acest proces se realizeaza prin expunerea semintelor umectate si stratificate la temperaturi coborate. Prin acest tratamente se modifica raportul dintre hormonii stimulatori si inhibitori in favoarea primilor. Astfel, continutul in gibereline din seminte creste dupa 2-3 saptamani de postmaturare, cel de citochinine dupa 4 saptamani si cel de auxinne, dupa sapatamani, concomitent cu diminuarea continutului in acid abscisic.
Tabelul 25
Durata perioadei de mentinere a facultatii germinative, a perioadei postmaturare si epoca de stratificare
|
Specia |
Durata de mentinere a facultatii germinative |
Durata de postmaturare |
Epoca de stratificare |
|
Armeniaca vulgaris |
6 luni |
5 luni |
august |
|
Castanea sativa |
2-3 luni |
5 luni |
oct.-noiembrie |
|
Cerasus avium |
7 sap. |
6 luni |
august |
|
Juglans regia |
6 luni |
5 luni |
oct. |
|
Malus domestica |
6 luni |
8-10 sapt. |
10-15 zile dupa recoltare |
|
Persica vulgaris |
6 luni |
5 luni |
oct. |
|
Pyrus communis |
6 luni |
8-10 sapt. |
10-15 zile dupa recoltare |
|
Prunus domestica |
6 sapt. |
5 luni |
august |
|
Vitis vinifera |
4 ani |
3-4 luni |
oct.-noiembrie |
Dupa Baldini (1986), durata perioadei de mentinere a facultatii germinative si a postmaturarii semintelor la temperatura de 4-50C variaza in functie de specia arborilor fructiferi, in limitele prezentate in tabelul 25.
Maturarea semintelor de Triticuum se realizeaza in momentul in care greutatea proaspata scade, iar semintele incep sa se deshidrateze (Mitchell s.a., 1980).
Permeabilitatea inselisului seminal variaza mult in functie de specie. Invelisul permeabil este alcatuit din celule moarte, care au intre ele spatii mari pline cu aer. Invelisurile impermeabile sunt alcatuite la exterior din mucilagii (Malus) sau din celule strans unite, fara spatii intre ele (Lactuca)
Stimularea germinarii semintelor cu invelisul impermeabil se poate realiza prin scarificare.
Dupa semanat, conditiile corespunzatoare de temperatura, lumina si umiditate determina intreruperea repausului seminal.
Temperatura optima pentru germinarea semintelor variaza intre 25-280C pentru plantele originare din zonele temperate si 30-350C pentru cele tropicale. Dupa Noggle si Fritz (1984), temperatura minima, optima si maxima pentru germinarea semintelor este urmatoarea:
|
Specia |
Minim |
Optim |
Maxima 0C |
|
Triticum sativum |
|||
|
Hordeuum sativum |
|||
|
Secale cereale |
|||
|
Zea mays |
|||
|
Oryza sativa |
|||
|
Cucumis melo |
In cazul semintelor de Lactuca sativa, germinarea are loc numai dupa expunerea acestora la luminna, cateva minute sau ore, in functie de specie. Prin aceasta fitocromul inactiv (P660) se transforma in fitocrom activ (P730), care stimuleaza trecerea de la starea de repaus vegetativ la cea de viata activa, adica la germinarea semintelor.
Aceste seminte au nevoie de un raport ridicat fitocromi activi/fitocromi totali. Astfel, semintele de Lactuca germinate au valoarea raportului P730/Pt mai mare de 0,4, in timp ce semintele nefotoblastice au valoarea acestui raport mai mica de 0,02.
Pe parcursul procesului de germinare a semintelor se pot deosebi 4 etape mai importante.
In prima etapa, care are loc la contactul direct dintre seminte si apa care alcatuieste solutia solului, se desfasoara procesul de imnibitie a semintelor. Pe durata acestui proces, substantele fenolice din tegument si alti inhibitori difuzeaza in solutia solului, eliminandu-se astfel unul din factorii ce induc repausul seminal. Apa din solutia solului patrunde prin tegument, in cazul in care este permeabil, sau prion hil sau micropil, in cazul semintelor cu tegumentul impermeabil, determina imbibitia coloizilor celulari. Semintele de Triticum, bogate in glucide, absorb circa 47 g apa, iar cele de Phaseolus bogate in proteine absorb 200-400 g apa, pentru 100 g seminte. In timpul imbibitiei semintelor hidrofilia citoplasmei creste, iar lipofilia si viscozitatea descresc. Ca urmare a desfasurarii acestui proces, radicalii liberi ai coloizilor celulari leaga dipolii apei si isi maresc volumul, generand presiunea de imbibitie. Aceasta depaseste 1000 de bari, determinand ruperea tegumentului, ceea ce stimuleaza penetrarea apei si a oxigenului in samanta.
In contact cu solutia solului, semintele uscate ale caror membrane plasmatice sunt deteriorate, pierd prin difuzie o parte din substantele solubile. Cantitatea de substante solubile care difuzeaza in solutia solului este dependenta de ritmul de refacere a integritatii membranelor plasmatice si este corelata cu capacitatea de germinare a semintelor. In practica se determina conductometric cantitatea de ioni ce difuzeaza in solutia solului, care constituie un indicator al capacitatii de germinare a acestora.
In etapa a doua care are loc dupa hidratarea celulelor se constata stimularea activitatii enzimelor existente si a procesului de respiratie.
Biosinteza giberelinelor are loc in scuutelum, de unde migreaza in celulele cu aleurona (fig. 141). In ribozomii de pe reticulul endoplasmatic rugos, din celulele cu aleurona are loc sinteza "de novo" a α-amilazelor (Davies, 1987). Aceste enzime sunt transportate in complexul Golgi, de unde prin intermediul veziculelor golgiene ajung la plasmalema pe care o strabat prin pinocitoza. In continuare, amilazele sunt transportate pe cale apoplastica pana la endosperm, unde determina biodegradarea amidonnului (Purohit, 1985).
Plantele tinnere sunt dependente de auxina existenta in tesuturi, in timp ce ciotchininele sunt puuse in libertate din acizii nucleici, ca urmare a actiunii ribonucleazelor (Sebaneck, 1992).
Trecerea embrionului de la faza de repaus la cea de vegetatie este foarte rapida. Astfel, in semintele de Pisum dupa 14 ore de imbibitie are loc sinteza amilazelor, iar dupa 24 de ore se ajunge la viteza maxima de diviziune a celulelor. In aceasta perioada are loc si refacerea struucturii membranelor plasmatice, care s-a deteriorst in timpul repausului vegetativ.
Dupa Burton (1982), intensitatea procesului de repiratie a semintelor creste de 45,9 ori, prin sporirea continutului in apa de la 11,1 la 17,0 %.
Procesul de respiratie a semintelor creste foarte rapid in primele ore de imbibitie, cresterea este pusa pe seama activitatii enzimelor respiratorii (fig. 142.). In faza urmatoare, dupa 10-25 ore de la imbibitie, se constata o diminuare a intensitatii procesului de respiratie a seminntelor de Pisum. Catul respirator este 3,0, indicand existenta unui proces de respiratie anaeroba, care este determinat de penetrarea lenta a oxigenului prin pericarp. In fata a treia, dupa 25-60 de ore de la imbibitie, are looc o noua intensificare a procesului de respiratie, determinata de penetrarea oxigenului prin micile rupturi ale tegumentului, precum si datorita biosintezei "de novo" a enzimelor implicate in biodegradarea substratului respirator. In ultima faza dupa 60 de ore de la imbibitia semintelor, intensitatea procesului de respiratie scade, datorita bioodegradarii celulelor din cotiledoanele diin care substantele de rezerva au fost consumate.
O dinamica similara a intensitatii procesului de respiratie s-a constatat si la semintele de Phaseolus lunatus, Lactuca sativa, Raphanus sativus, Arachis hypogeae, Zea mays, Glycine max si Lathyrus odoratus (Bbewley si Black, 1978).
In semintele altor specii: Avena fatua, Hordeum vulgare, Ricinus communis, Sinapis arvensis, Oryza satica, etc. faza a doua de anaerobioza nu are loc. Intensitatea procesului de respiratie a semintelor, pe durata procesului de germinare, este determinata de temperatura (fig. 143) .
Studiile de microscopie electronica au precizat ca intensificarea procesului de respiratie este insotita in cazul semintelor de oryza de cresterea numarului de mitocondrii si a numarului cristelor mitocondriale (Yoo, 1970). In acelasi timp, s-a constatat intensificarea activitatii enzimelor ciclului Krebs (Breidenbanch s.a., 1967).
In etapa a treia a procesului de germinare are loc mobilizarea substantelor de rezerva dinn seminte (fig. 144.). Acest proces este initiat imediat dupa imbibitia semintelor, iar cele mai mari modificari se observa dupa 72 de ore (Noggle si Fritz, 1984).
Principalele modificari biochimice care au loc in timpul germinarii semintelor sunt prezentate an schema din figura 145.
Substatele de rezerva: glucidele, lipidele si protidele sunt biodegradate pana la zaharoza, aminoacizi, acetil CoA, glicerol, etc., care sunt translocate din endosperm sau cotiledoane spre embrion, unde sunt utilizate prin biosinteza compusilor implicati in procesul de crestere sau pentru producerea de energie biochimica in mitocondrii.
In scutelmul semintelor de Hordeum a fost identificata prezenta β-amilazei (Mory s.a., 1972), iar dupa 22-24 ore a fost pusa in evidenta si biosinteza β-glucozidazei si a α-amilazei in celulele cu aleurona.
Amilazele si maltaza catalizeaza biodegradarea amidonului din amiloplaste pana la glucoza, care este translocata in citoplasma, unde in prezenta unei izomeraze formeaza fructoza, iar in prezenta zaharozo frutozei, cele doua hexoze dau nastere la o molecula de zaharoza Acesta este translocata in embrion, unde este biodegradata la monoglucide care prin glicoliza formeaza acidul piruvic. Aceasta prin aminare formeaza aminoacizi, prin degradare produce acetil CoA din care se biosintetizeaza acizii grasi si prin biodegradare in ciclul Krebs se obtine energia necesara proceselor de biosinteza.
Semintele uscate de Pisum contin putini aminoacizi liberi, dar continutul acestora creste imediat dupa imbbibitie (Larson si Beevers, 1965). Cresterea respectiva este rezultatul ativitatii proteazelor din veziculele ce contin proteine. Aminoacizii sunt transportati in embrion, unde are loc sinteza proteinelor, in prezenta poliribozomilor, a ARN-m, ATP si a enzimelor. Biosinteza acestor substante a fost identificata in semintele de Phaseolus dupa 30-60 minute de la imbibitia acestora (Gillard si Walton, 1973).
ARN-m necesar pentru biosinteza proteinelor a fost identificat in semintele uscate de Triticum (Weeks si Marcus, 1971), probabil, asociat cu proteinele, iar in primele minute dupa imbibitie se evidentiaza cresterea cantitativa a ARN-m in embrion, urmare a biosintezei "de novo". Concomitent s-a constatat si cresterea numarului de poliribozomi. O situatie similara s-a observat si in cazul semintelor de Gosypium (Hammett si Katterman, 1975). Bbewley si Black (1982) apreciaza ca primele etape ale germinarii semintelor nu este necesara biosinteza "de novo" a ARN-m pentru sinteza proteinelor.
In semintele de triticum in curs de germinare a fost pus in evidenta procesul de biosinteza a ADN-polimerazei si a ARN-polimerazei (Mazus si Buchowicz, 1972), implicate in biosinteza acizilor nucleici, iar semintele de Phaseolus a fost pusa in evidenta activitatea aminoacil-ARNt-sintetazei, implicata in biosinteza proteinelor (Anderson si Fowden, 1969).
Principala substanta de rezerva din semintele plantelor oleaginoase o constituie lipidele, reprezentate prin trigliceride. Acestea se gasesc depozitate in sferozomi (oleozomi), cromoplaste sau sub forma de picaturi in citoplasma.
Biodegradearea lipedelor de rezerva din semintele de Ricinus a fost studiata de Cavin si Beevers (1961), iar ceea a lipidelor din semintele de Helianthus annuus de Brodbeer si Stumpf (1959).
Lipidele din sferozomi sunt biodegradate in prezenta lipazelor pana la glicerol si acizi grasi care sunt translocati in glioxizomi, unde sunt catabolizati prin procesul de β-oxidare, pana la acetil CoA. Aceasta poate fi biodegradata in ciclul Krebs sau poate fi metabolizata in ciclul acidului glioxilic, care se desfasoara in glioxizomi. Compusul final al acestui proces este succinatul, care poate fi metabolizat in mitocondrii pana la oxalat. Acesta este translocat in citoplasma, unde este transformat in piruvat, care printr-un proces de glicoliza inversa este transformat in glucozi (fig. 146), poate fi utilizat in biosinteza aminoacizilor sau poate fi metabolizat in ciclul Krebs.
Metabolizarea glucidelor de rezerva din semintele de Ricinus a fost pusa in evidenta de Muto si Beevers (1974) dupa trei zile de la imbibitia acestora.
In etapa a patra a procesului de germinare a semintelor are loc cresterea radiculei. Compusii intermediari rezultati din biodegradarea substantelor de rezerva si energia biochimica produsa in respiratie sunt utilizate de embrionul seminal in procesul de crestere. Mai intai are loc extensia celulelor radiculei, iar ulterior diviziunea lor. Dupa cateva zile de imbibitie, radicula embrionului penetreaza spre exterior, prin micropil sau prin rupturile pericarpului, ceea ce marcheaza sfarsitul procesului de germinare si inceputul procesului de crestere a plantulei.
APLICATII PRACTICE DE LABORATOR
Starea de repaus constituie o perioada de viata latenta instaurata ca o reactie de adaptare a plantelor la trecerea alternativa de la sezonul cald la sezonul rece, cu factori nefavorabili desfasurarii proceselor biologice.
Parcurgerea perioadei de repaus se face sub forma de seminte, muguri, bulbi, rizomi, tuberculi etc. care pastreaza viabilitatea plantelor pe durata sezonului nefavorabil.
Stabilirea viabilitatii semintelor prin tesutul histochimic cu T.T.C. (metoda Lakon)
Principiu. Prezenta celulelor vii sau moarte din seminte aflate in stare de repaus se pune in evidenta prin tratamente cu diferite substante care isi modifica culoare in functie de actiunea substratului enzimatic.
Materiale necesare seminte de mazare, cristalizor cu apa, bisturiu, vas Petri, solutie de 1% T.T.C. (clorura de 2, 3, 5 - trifeniltetrazolium).
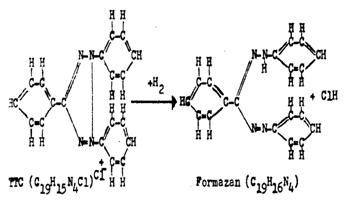
Fig. 147 Transformarea enzimatica a T.T.C. in formazan
Modul de lucru Se numara 100 seminte de mazare care se pun la imbibat timp de 2 ore. Semintele imbibate cu apa se sectioneaza longitudinal si se pun intr-un vas Petri care contine solutie de 1% de T.T.C. unde raman la temperatura camerei, in intuneric timp de o ora. Se scot semintele din solutie si se examineaza individual: cele viabile au embrionul colorat in rosu carmin, iar cele neviabile au embrionul necolorat.
Interpretare La tesuturile vii complexul enzimelor respiratorii (succin - dehidraze) transfera H2 de pe substratul biologic pe solutia incolora de T.T.C. care este redusa intr-un compus colorat in rosu numit formazan (fig. 147).
Durata experientei observare 15 minute dupa pregatirea semintelor timp de 3 ore.
Stimularea germinatiei semintelor prin scarificarea tegumentului
Principiu. Prin scarificarea tegumentului seminal se rupe o parte din tegumentul impermeabil pentru a usura patrunderea la embrion, a apei si a oxigenului.
Materiale necesare seminte de sulfina (Melilotus officinalis), lucerna (Medicago sativa) sau mazariche (Vicia sativa), doua placi captusite cu glaspapir, acid sulfuric concentrat (d = 1,84).
Modul de lucru Semintele de sulfina, lucerna sau mazariche se impart in trei loturi: lotul I se pune la germinat cu tegumentul intact; la lotul II se scarifica tegumentul seminal inainte de a trece la germinatie folosind metoda mecanica de frecare a semintelor intre doua placi captusite cu glaspapir; la lotul III se scarifica tegumentul pe cale chimica prin imersia semintelor timp de cateva minute in acid sulfuric concentrat si apoi se spala cu apa. Dupa mentinerea semintelor la germinat timp de 24 de ore se compara procentul semintelor germinate in cele trei loturi.
Interpretare. Semintele unor specii din familiile Leguminoasae si Malvaceae prezinta un tegument tare, impermeabil pentru apa si oxigen ceea ce impiedica procesele de imbibitie si germinatie. Prin scarificare pe cale mecanica sau pe cale chimica se subtiaza tegumentul seminal dand posibilitatea declansarii germinatiei semintelor.
Durata experientei observare 10 minute dupa 24 de ore de germinatie.
Stimularea germinatiei prin incalzirea semintelor
Aparate si dispozitive: termostat electric.
Materiale necesare: seminte de castraveti sau pepeni, vas Petri, hartie de filtru.
Modul de lucru In vasul Petri captusit cu hartie de filtru se pun la germinat 100 seminte. Apa de imbibitie se incalzeste la 35-400C inainte de a umecta hartia de filtru. Vasele Petri se tin timp de 24 de ore intr-un termostat electric la temperatura de 35-400C. In paralel se pun la germinat 100 seminte, dar vasele Petri se tin la temperatura de 18-200C. Dupa 24 de ore vasele Petri se trec la temperatura camerei. Timp de o saptamana se noteaza zilnic procentul de seminte germinate la cele doua variante, urmarindu-se energia germinativa si facultatea germinativa a semintelor.
Interpretare. Incalzirea semintelor determina o stimulare a incoltirii semintelor, marind atat energia germinativa (procent de seminte germinate intr-un interval de timp limitat - 3 zile) cat si facultatea germinativa (procente de germinare a semintelor pe o durata nelimitata de timp).
Durata experientei: observatii de 10 minute pe o perioada de 7 zile de germinare a semintelor.
Intreruperea fortata a repausului la mugurii floriferi
Principiu La plantele lemnoase si la unele plante erbacee (bulboase, tuberculifere etc) repausul hibernal se realizeaza prin muguri, ca o adaptare biologica la conditiile nefavorabile din timpul iernii. Durata repausului biologic mugural este variata in functie de specie si de aceea iesirea din repausul hibernal se esaloneaza pe o lunga perioada in primavara. Scurtarea sau prelungirea perioadei de repaus biologic mugural se poate dirija de catre om folosind diferite metode chimice, fizice si agronomice.
a. Metoda bailor calde (H.H. Molisch).
Aparate si dispozitive: termostat.
Materiale necesare ramura de liliac (Syringa vulgaris) sau de forsitia (Forsythia intermedica), vase cu apa, bec de gaz, termometru, vas Dewar, flacon Erlenmeyer.
Modul de lucru Ramuri de liliac sau forsitia aflate in stare de repaus biologic mugural (lunile decembrie-ianuarie) se scufunda total sau partial intr-un vas cu apa calda la temperatura de 30-350C, timp de 12 ore. Temperatura apei se controleaza la intervale scurte de timp pentru mentinerea intre limitele propuse. Pentru aceasta se introduce vasul intr-un termostat sau se utilizeaza un vas Dewar cu pereti termoizolanti. In paralel se aleg ramuri control, tinute in apa la temperatura camerei. Dupa tratamentul termic se scot ramurile din vasele cu apa si se trec cu baza in flacoane Erlenmeyer cu apa, unde se tin 2 saptamani. Se observa ca dupa circa o saptamana, ramurile supuse tratamentului cald inmuguresc pe cand cele control se mentin in stare de repaus.
Interpretare. Tratamentul termic intrerupe repausul biologic al mugurilor si declanseaza trecerea la starea de viata activa.
Durata experientei 1-2 saptamani.
b. Metoda tratarii cu eter.
Materiale necesare: ramuri de liliac (Syringa vulgaris) sau rizomi de lacramioara (Convallaria majalis) flacon Erlenmeyer, cristalizor, eter, acid sufluric concentrat (d = 1,84).
Modul de lucru In flaconul Erlenmeyer cu apa, se pun ramuri de liliac sau rizomi de lacramioare care se trec sub un clopot de sticla in care se afla un cristalizor cu acid sufluric concentrat si un pahar Berzelius cu eter (0,5ml la un litru de aer). Se mentin plantele in aceasta atmosfera timp de 1-2 zile, dupa care se scot si se trec in atmosfera camerei. In paralel, se urmaresc ramurile control tinute numai in atmosfera camerei, cu baza in flacoane Erlenmeyer cu apa. Dupa cateva zile se constata ca la plantele tratate cu eter mugurii se deschid si declanseaza inflorirea, in timp ce la plantele control mugurii raman in stare de repaus biologic.
Interpretare Tratamentul chimic cu vapori de eter determina intreruperea repausului biologic mugural.
Durata experientei montarea si observarea 10 minute la un interval de cateva zile.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 10157
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved