| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
TULPINA
Primul grup de plante la care se constanta existenta unei tulpinite il constituie Muschii frunzosi din Increngatura Bryophyta, la care, pe o tulpinita dreapta se insera, in mod ordonat, frunzulitele, sugerand o organizare similara cu a plantelor vasculare. Totusi aceasta tulpinita a Bryophytaelor difera de cea a plantelor vasculare prin faptul ca ea reprezinta gametofitul, constituit din celule haploide si nu sporofitul ca la plante vasculare; In al doilea rand tulpinita Bryophytelor este foarte putin diferentiata, lipsita de un tesut conucator libero-lemnos tipic. La aceste plante nu exista nici radacini, tulpinita prezentand la partea sa bazala rizoizi.
Rezulta ca o tulpina adevarata vom intalni numai la grupul de plante Cormofite, care au diferentiate toate cele 3 organe vegetative: radacina, tulpina si frunza si aici se incadreaza Pteridophytele, Gymnospermele si Angiospermele.
La Cormofite tulpina reprezinta organul vegetativ ce creste in pozitia verticala, prezinta, de regula, o simetrie radiara, fiind adaptata mediului aerian. Rolul tulpinii este de a sustine frunzele, ramurile si organele de reproducere si face totodata legatura intre radacina si frunze, adica asigura si conducerea sevei brute si a celei elaborate, prezentand deci atat un rol morfologic cat si unul fiziologic. Tulpina prezinta o crestere terminala , varful vegetativ nefiind protejat de o formatiune speciala ca In cazul radacinii.
Originea filogenetica se refera la modul de diferentiere a tulpinii in decursul evolutiei regnului vegetal. Exista teoria filomului care sustine ca tulpina s-a dezvoltat pornind de la baza frunzei. Teoria cauloidului, bazata pe date paleontologice, sustine ca tulpina a aparut inaintea frunzelor, odata cu adaptarea plantelor la modul de viata terestru. Frunzele s-au dezvoltat ulterior, fie ca emergente, fie prin turtirea organelor caulinare (varfurile ramificatiilor), aceasta teorie fiind in prezent aproape unanim acceptata.
Originea
ontogenetica se refera la diferentierea tulpinii
in cursul dezvoltarii individuale a plantelor. Tulpina isi are originea in embrionul
semintei: din tigela se va forma axa
hipocotila, cuprinsa intre baza tulpinii si
cotiledoane iar din gemula (sau plumula) se va forma axa
epicotila, cuprinsa intre cotiledoane si prima
frunza normala, precum si celelalte internoduri ale tulpinii
purtatoare de
Legatura intre radacina si tulpina se realizeaza in regiunea coletului.
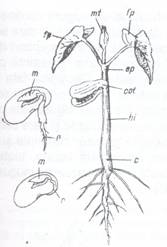
Fig. 100. Plantule de fasole in diferite stadii de dezvoltare:
r- radicula; m- mugurasul; c- colet; hi- axa hipocotila; ep- axa epicotila; cot- cotiledoane; fp-frunze primare; mt- mugur terminal.
Morfologia tulpinii
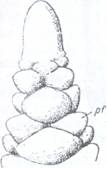
Portiunea de tulpina impreuna cu frunzele formate pe ea se numeste lastar. Acesta se formeaza pe seama activitatii conului vegetativ (Fig. 101) care determina cresterea in lungime a internodurilor.
A B
Fig. 101. Conul vegetativ al
tulpinii de Elodea, cu primordii de
A- vazut din
profil; B- vazut in sectiune longitudinala; d-
dermatogen;
pf- primordii foliare, T- tunica; c- corpusul.
Desi tulpinile celor mai multe cormofite traiesc in aer, exista numeroase plante cu tulpini subterane sau acvatice, aceste medii producand schimbari profunde in morfologia si anatomia lor.
1.1. Tulpinile aeriene
Locul mai umflat (dilatat) de pe tulpina in care se insera frunzele poarta numele de nod, iar portiunile cilindrice mai subtiri cuprinse intre doua noduri se numesc internoduri sau internodii. Unghiul in care se insera frunza pe tulpina se numeste axila sau subsioara frunzei. In acest loc se gasesc plasati mugurii axilari sau laterali iar ultimul internod se sfarseste cu mugurele terminal (Fig. 102, 103).
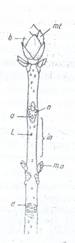

Fig. 102. Ramura de castan salbatic: (Aesculus hippocastanum) Fig. 103. Muguri de la Aesculus hippocastanum;
mf- mugur terminal; n- noduri; in- internodii; m.a - muguri axilari; c- catafile; cf- cicatricea frunzei; ma- mugure axilar;
a- cicatricele frunzelor cazute; b-catalife; mt- mugure terminal; uf- urmele fascicolelor vasculare.
d- cicatricele catafilelor cazute; L - lenticele.
1.1.1. Mugurii Din punct de vedere structural un mugure
contine in partea centrala conul vegetativ al tulpinii avand grupuri
de celule initiale sub care se gaseste meristemul primordial,
nascut prin diviziunile acestora. Lateral, incepand de
la baza spre varf conul vegetativ contine proeminente exogene
care reprezinta primordiile frunzelor, iar la subsioara lor
primordiile mugurilor axilari. Deoarece fata
extema a primordiilor foliare creste mai repede decat cea
interna, aceste primordii se acopera unele pe altele si
acopera si varful conului vegetativ. Intregul mugure este protejat de solzi bruni care reprezinta
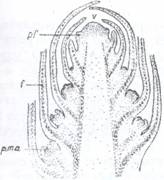
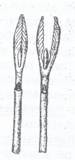
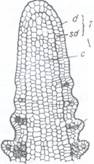
Fig. 104. Sectiune logitudinala printr-un mugur terminal: Fig. 105. Muguri nuzi de Viburnum lantana.
V- Varful vegetativ al tulpinii; pf- primordii foliare; p.m.a.
- primordii de muguri axilari; f -
Clasificarea mugurilor se poate face si in
functie de organele ce se vor dezvolta din ei.
Astfel pot fi muguri foliari, florali, si micsti
dupa cum vor da nastere la
Mugurii normali sunt cei ce se nasc (se dezvolta) din primordiile conului vegetativ. Ei se formeaza vara si raman in stare de repaus iarna, dezvotandu-se in primavara anului urmator. Multi dintre ei raman in stare de latenta chiar mai multi ani si astfel de muguri se numesc muguri dorminzi si reprezinta o rezerva a plantei in caz de inghet, distrugerea frunzelor de catre agenti patogeni sau insecte, distrugerea mecanica a frunzelor.
Mugurii adventivi sunt acei muguri care nu se dezvolta din primordiile conului vegetativ sau la subsioara frunzelor ci in locuri nedeterminate, pe ramuri, radacini sau pe frunze si, spre deosebire de mugurii normali, au o origine endogena (din cambiu sau din periciclu). Mugurele axilar cel mai apropiat de mugurele terminal se numeste mugure de inlocuire si poate lua locul acestuia in caz de distrugere. Mugurele terminal exercita o actiune inhibitoare asupra mugurilor axilari si prin taierea lui se favorizeaza dezvoltarea mugurilor axilari, procedeu utilizat in pomicultura in scopul formarii coroanei arborilor si dirijarii cresterii acestora.
1.1.2. Ramificatia tulpinii
In cazul palmierilor, unor graminee (ex. bambusul) si alte cateva Monocotiledonate, tulpinile raman constituite dintr-un ax unic neramificat. La cele mai multe plante insa tulpina se ramifica in mod diferit: dichotomic monopoidal si simpodial. Ramificatia dichotomica consta dintr-o bifurcare a varfului vegetativ in doua ramuri de ordinul I a caror varf se bifurca in acelasi mod, In cate doua ramuri de ordinul II s.a.m.d. Dicotomia reprezinta un mod primitiv de ramificare ce era intalnit la multe plante din era paleozoica, azi fiind intalnit la Lycopodium. Selaginella. Ramificatia monopoidala (Fig. 106 A), se caracterizeaza prin existenta unui ax principal mai lung si mai gros si care creste nelimitat in lungime prin mugurele terminal. Din mugurii axilari ai acestuia se dezvolta ramuri laterale de ordinul 1, care la randul lor produc ramuri de ordinul II si care se comporta in acelasi mod, formarea ramurilor avand un sens acropetal. Asezarea ramurilor pe axul principal poate fi altelma, opusa sau verticilata. (Fig. 107). Ramificatia simpodiala (Fig. 106 B) prezinta axul principal format prin suprapunerea mai multor ramuri de ordine diferite, tulpina principala avand o crestere limitata in lungime, prin disparitia mugurelui terminal se favorizeaza aparitia unor ramificatii simpoidale, procedeu utilizat in pomicultura.

Fig. 106. Scheme ale ramificatiei monoidale A si simpodiale B a tulpinii
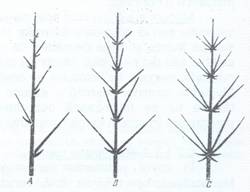
Fig.107. Scheme ale ramificatiei monopoidale, cu ramuri dispuse altern A, opuse B si verticilat C
Ramurile arborilor si arbustilor pot avea doua aspecte: ramuri scurte si ramuri lungi. Cele scurte se mai numesc si microblaste sau brachiblaste, au o crestere anuala foarte redusa, noduri dese si internoduri scurte. Ele nu se ramifica si cad dupa cativa ani dar au importanta mare in cazul pomilor fructiferi, producand de regula flori si fructe, de aceea se mai numesc si ramuri de rod. Ramurile lungi se mai numesc macroblaste, au cresteri anuale foarte accentuate cu noduri rare si internoduri lungi.
Ramificatii anormale formate din numeroase ramuri subtiri si dese, observate uneori pe arbori si arbusti si denumite popular 'maturi de vrajitoare' se datoresc actiunii unor insecte sau ciuperci parazite prin care se declanseaza dezvoltarea simultana a mai multor muguri dorminzi.
Habitusul arborilor reprezinta infatisarea caracteristica a fiecarei specii lemnoase. Acest aspect exterior, habitusul sau portul arborilor depinde de tipul de ramificatie al tulpinii, de numarul ramurilor, unghiul de ramificare, de fenomenele de corelatie dintre muguri, de densitatea plantelor, lumina fiind un factor de mediu important in aceasta privinta.
Durata in timp a arborilor este legata de specie. Cea mai lunga durata o are chiparosul (Toxodium mexicanum), baobabul (Adansonia digitata), care pot atinge cca. 5.000 ani. Arborele mamut (Sequoia gigantea) 4.000 ani, cedrul de Liban (Cedrus libani) si tisa (Taxus baccata) 3.000 ani, maslinul (Olea europea) si castanul comestibil (Castanea sativa) 2.000 ani, stejarul (Quercus robur) si molidul (Picea excelsa) 1.200-1.500 ani, teiul (Tilia sp.) si fagul (Fagus silvatica) 900-1.000 ani.
Inaltimea arborilor poate atinge 142 m (Sequoia gigantea) si circumferinta de 36 m iar Eucaliptus globulus 155 m inaltime. Unele liane tropicale ating si 300 m lungime dar ele se incolacesc si se sustin pe alti arbori.
1.1.3. Clasificarea tulpinilor aeriene
In clasificarea tulpinilor aeriene se tine seama de mai multe criterii.
cilindrice (paiul de graminee),
comprimate (Opuntia),
prismatice cu sectiunea triunghiulara (la Cyperaceae),
patrunghiulara (Lamiaceae), pentagonala (dovleac)
Tulpinile Umbeliferelor poarta creste logitudinale si se numesc sulcate, iar la Equisetaceae tulpinile se numesc striate si prezinta dungi longitudinale foarte fine.
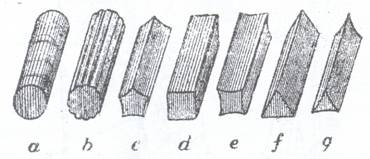
Fig. 108. Forme de tulpini: a- tulp. cilindrica; b-tulp. sulcata; c,d,e,f- diferite tipuri de tulpini prismatice.
Dupa organizare, adica dupa cum
impartirea in noduri sau internoduri este
evidenta sau nu, tulpinile se clasifica in articulate si
nearticulate. Daca sunt goale in interior si au
nodurile umflate se numesc pai sau culm, iar
tulpina fara noduri umflate, cu
Tulpinile nearticulate
prezinta in realitate, nodurile foarte dese, incat toate frunzele dau
impresia ca se prind la acelasi nivel. Asemenea tulpini scurte,
carnoase, neramificate si care traiesc mai multi ani se
intalnesc la Agave, Sempervivum. Altele sunt insa inalte, cu
frunzele columnare, purtand numai in varf un buchet de
3. Dupa orientarea in spatiu tulpinile pot fi ortotrope si plagiotrope. Cele ortotrope pot fi, la randul lor, drepte si urcatoare. Tulpinile drepte pot fi erecte daca au pozitia verticala, de la baza pana la varf, nutante, daca au varful aplecat in jos, geniculate daca au partea bazala oblica sau orizontala si, incepand de la un nod, se indreapta (Fig. 109). Tulpinile urcatoare nu prezinta tesuturi mecanice capabile sa le mentina in pozitia verticala. Ele pot fi agatatoare si volubile. Cele agatatoare se agata de suport fie cu peri agatatori (Galium aparine), fie cu carcei (la mazare sau la vita de vie Bryonia. Echinocystis) sau cu radacini adventive ca la iedera (Hedera helix) (Fig. 110)
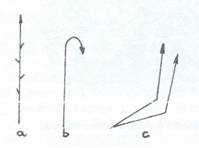
Fig. 109. Tulpini drepte: a- erecte; b- nutante; c-geniculante.
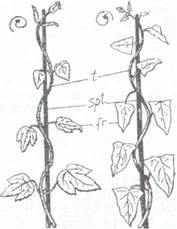
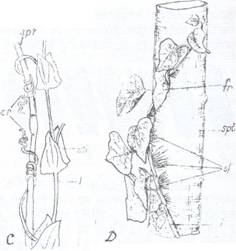
Fig. 110. Tulpini volubile: A- Humulus lupulus; B- Convolvus arvensis;
fr- frunza; spt- suport; t- tulpina si agatatoare la Lathyrus aphaca (C) si iedera (Hedera helix) (D)
Tulpinile volubile se invartesc in jurul altor plante sau suport, ca de ex. la hamei (Humulus lupulus) sau volbura (Convolvulus arvensis).
Tulpinile plagiotrope cresc orizontal sau oblic, neavand nici un stereom dezvoltat nici organe agatatoare. Unele stau culcate la pamant, impreuna cu ramurile si se numesc prostrate (ex. Herniaria glabra) altele taratoare, avand tulpina prinsa de pamant cu radacini adventive ex. coada racului - (Potentilla anserina), galbasoara (Lysimachia nummularia) (Fig. 111). Alte plante prezinta tulpini erecte dar cu ramuri taratoare, care se numesc stoloni si care la noduri formeaza radacini adventive si realizeaza pe aceasta cale o inmultire vegetativa (Fig. 112), ex. la fragi, menta.
Unele plante prezint tulpina taratoare cu ramuri ascendente (ex. cimbrisorul de camp (Thymus serpyllum), saschiu (Vinca minor) iar la Lycopodium clavatum de pe tulpinile taratoare se ridica numai ramurile fertile purtatoare de spice sporifere.
Fig. 111. Tulpini taratoare de galbasoara (Lysimachia nummularia)
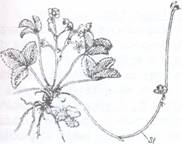
Fig. 112. Planta de fraga (Fragaria vesca) cu stoloni (St)
4. Dupa durata vietii
tulpinilor si numarul infloririlor Angiospermele se impart in
doua mari grupuri: monocarpice care
infloresc o singura data in viata lor dupa care mor si
policarpice, care infloresc si fructifica de mai multe
ori. Cele monocarpice pot fi, la randul lor, anuale,
bienale si plurianuale. Cele anuale
traiesc un singur an sau chiar mai putin,
perioada in care infloresc si produc fructe. Plantele
anuale pot fi, la randul lor efemere, cu vegetatie
foarte scurta, de cateva saptamani. Aceste
plante se inmultesc repede, avand mai multe generatii Intr-o
perioada de vegetatie (ex. Stellaria, Veronica, Lepidium).
Se noteaza cu O plantele anuale de vara (se
noteaza cu 0) sunt acelea care germineaza primavara si mor toamna (ex. inul, ovazul, porumbul, tutunul,
macul), plantele anuale de iarna (notate cu 00) care
germineaza toamna, ierneaza cu o tulpinita frunzoasa,
iar primavara urmatoare fructifica si apoi mor (ex. graul -
Triticum vulgare, Centaurium umbellatum). Plantele bienale (se
noteaza nu 0) Isi dezvolta in primul an radacina
si o rozeta de
Plantele policarpice traiesc mai multi ani si pot produce flori si fructe de mai multe ori in viata. Dupa consistenta tulpiniilor, plantele policarpice pot fi: ierboase, perene, carnoase, lemnoase.
Plantele ierboase perene (sau vivace) pot trai mai multi ani prin organele lor subterane, dar tulpinile lor aeriene mor in fiecare an. Plantele carnoase prezinta tulpini metamorfozate, groase si asimilatoare, fara frunze, sau cu frunze transformate in spini, cu tesuturi acvifere bine dezvoltate (Cactaceae, Crassulaceae, Euphorbiaceae ).
Plantele lemnoase se pot prezenta sub forma de semiarbusti, arbusti si arbori. Semiarbustii au baza tulpinii lemnoasa, aparata de suber si care nu ingheata iarna, iar portiunile dinspre varf, ierboase si moi, ce mor in fiecare an, ca de ex. la pelin (Artemisia absinthium), rozmarin (Rosmarinus officinalis), isop (Hysopus officinalis), lavanda (Lavandula), afin (Vaccinium myrtillus), jales de gradina (Salvia officinalis).
Arbustii prezinta tulpina complet lemnoasa,
dar nu au un trunchi, tulpina lor ramificandu-se
direct de la baza. Inaltimea acesteia este
de regula redusa (3-6 m) ca de ex. la soc (Sambucus nigra), porumbar
(prunus spinosa), maces (
Arborii prezinta tulpina lemnoasa, inalta si groasa, diferentiata in trunchi si coroana.
1.1.4. Adaptarea tulpinii la mediu (Tulpini metamorfozate)
Adaptarea tulpinilor aeriene la functii nespecifice duce la modificarea formei si structurii acestora, cele mai frecvente functii nespecifice fiind: asimilatia, depozitarea de substante de rezerva, inmultirea vegetativa si apararea.
1. Tulpini asimilatoare indeplinesc functia de fotosinteza, prezinta o culoare
verde si tesuturi asimilatoare. Asemenea tulpini se intalnesc la
unele plante din desert, apartinand fam. Cactaceae (Fig.113)
si Euphorbiaceae, cu tulpini groase si carnoase, cu
frunze reduse la solzi sau transformate in spini, cu cuticula groasa
si stomate putine si adancite, cu suc vacuolar concentrat, bogat
in mucilagii si acizi organici, capabile sa absoarba si sa
retina apa pe care o depoziteaza in tesuturi acvifere bine
dezvoltate. La pipirig (Juncus effusus) si coada calului (Equisetum
arvense), tulpinile aslmilatoare au forma cilindrica, fara
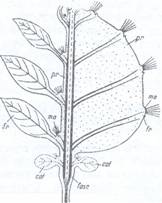
Fig.113. Formarea tulpinii la cactaceae: in partea
stanga a figurii este reprezentata o
cactacee cu
pr-scoarta primara la
suculente s-a transformat intr-un tesut acvifer (punctat); fasc-fascicule
conducatoare; fr-frunze; m.a.-muguri axilari cu
![]()
Fig.114. Genista sagittalis, tulpina transformata in organ asimilator (cladodiu).

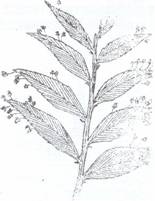
Fig.115. Ramuri transformate in organe asimilatoare (filocladii)
la Ruscus aculeatus (stg), Phyllanthus (dr.)
2. Tulpinile care depoziteaza materii de rezerva isi dezvolta foarte mult parenchimurile si ca urmare se tuberizeaza, marindu-si mult volumul. Asa se intalnesc de ex. in cazul tulpinii comestibile de gulie.
3. Tulpini (aeriene) adaptate la functia de inmultire vegetativa se intalnesc la grausor (Ranunculus ficaria) la care, la subsioara frunzelor se formeaza muguri axilari, fara frunze, cu partea centrala plina cu substante de rezerva. Aceste formatiuni se numesc tuberule, se pot desprinde si dau nastere la noi plante. La dentita (Dentaria bulbifera) mugurii axilari se numesc bulbili si prezinta frunzisoarele tuberizate, pline cu substante de rezerva (Fig.116).
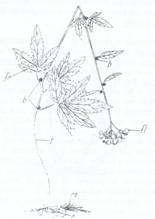
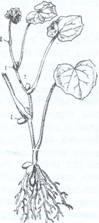
Fig.116. Muguri transformati in organe de inmultire vegetativa:
bulbili (b) la Dentaria bulbifera (stg) si tuberule la subsioara frunzelor de Ficaria verna (grausor)
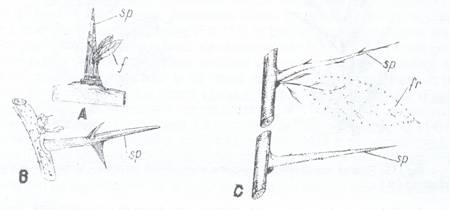
Fig.117. Tulpini transformate in spini
A - La Prunus spinosa; B - La Gleditschia triacanthos; C - La Crataegus monogyna.
1.2. Tulpinile subterane
In cazul acestor tulpini se intalnesc
adaptari la viata sub nivelul solului, dar in acelasi timp
reprezinta tulpini metamorfozate, adaptat la depozitarea de materii de
rezerva. Traind in acelasi mediu cu radacinile se
aseamana cu acestea, dar pot fi deosebite de acestea prin
urmatoarele caractere: au varful neacoperit cu piloriza, poarta
muguri la subsioara unor
1. Rizomii sunt considerati microblaste subterane
si mai rar macroblaste. Dupa pozitia lor in sol rizomii pot fi ortotropi,
cand cresc vertical in sol ca de ex. la ciubotica cucului (Primula
sp.), plagiotropi cand cresc oblic (ex. Arnica montana,
Drvopteris filix-mas) si orizontali cand cresc paralel
cu suprafata solului cum sunt rizomii de lacramioare (Convallaria
maialis) (Fig.118). Forma rizomilor poate fi foarte
variata (cilindrica, comprimata, filiforma) la fel si
dimensiunile lor. De la nodurile rizomilor se
formeaza radacini adventive, iar din muguri se vor forma tulpinile
aeriene. Rizomii pot avea o crestere indefinita prin
mugurele terminal, care creste orizontal in pamant in timp ce lastarii aerieni iau nastere din mugurii
axilari (ex. la lacramioara, dalac
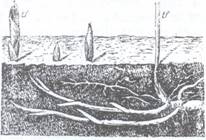

Fig.118. Rizomi de lacramioare cu crestere Fig.119. Rizom orizontal de pecetea lui indefinita (Convallaria majalis): tf-tulpini florifere Solomon Polygonatum multiflorum)
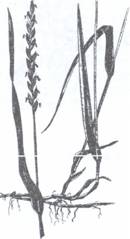
Fig.120. Paris quadrifolia. Fig.121. Stoloni subterani de pir (Agropyron repens)
Rizom cu ramificatie monopodiala (H)
2. Tuberculii reprezinta microblaste subterane scurte, groase si carnoase, formate, in cea mai mare parte, din tesuturi de depozitare a materiilor de rezerva si purtand pe suprafata lor rudimente de frunze in forma de solzi, la subsioara carora se afla mugurii denumiti si 'ochiuri'. Tuberculul poate proveni prin transformarea axei hipocotile (Corvdalis, Cyclamen), care raman in pamant si se tuberizeaza, dar la majoritatea plantelor cu tuberculi, inclusiv la cartof, tuberculul se formeaza prin ingrosarea varfului unor stoloni subterani. In cazul tuberculului de cartof, ca urmare a aparitiei unei zone generatoare felogenice se formeaza un suber care va inveli la exterior tuberculul, iar spre interior o scoarta secundara, bogata in substante de rezerva, in primul rand amidon (Fig.122).

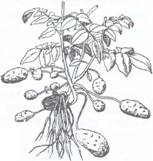
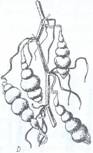
Fig. 122. Diferite forme de tuberculi A-B-cartof, C - la napi porcesti (Helianthus tuberosus) si D - la Stachys
3. Bulbo-tuberii fac
trecerea intre tuberculi si bulbi fiind inveliti la exterior in
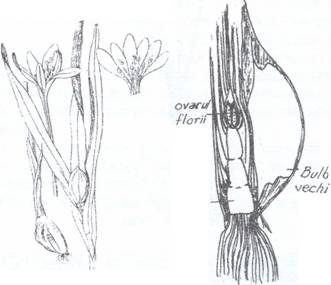
Fig.123. Bulbo-tuberi de sofran (Crocus) (1) si brandusa de toamna (Colchicum) (2)
A - In sectiune longitudinala
4. Bulbii reprezinta microblaste subterane, invelite de
Dupa cum frunzele solzoase ale bulbului se acopera complet unele pe altele bulbii pot fi tunicati ca la lalea (Tulipa gesneriana) si ceapa (Allium cepa) (Fig.124), zambila (Hyacynthus) sau pot fi solzosi cand solzii nu se acopera complet, ca la crin (Lilium candidum) (Fig.125).
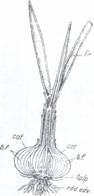
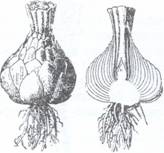
Fig.124. Allium cepa, Bulb in sectiune Fig.125. Bulb solzos de crin (Lilium candidum)
longitudinala: fr-frunze; cat-catafile; bf-baza
frunzelor plina cu materii de rezerva; tulp-tulpina,
rad.adv. - radacini adventive.
1.3. Tulpinile acvatice
Plantele acvatice pot pluti la suprafata apei si in cest caz se numesc natante ca de ex. la lintita de apa (Lemna minor) si pestioara (Salvinia natans) sau pot fi scufundate sub nivelul apei, caz in care se numesc submerse (ex. Elodea). Exista si plante amfibii ce pot trai si pe uscat si in apa ca de ex. limbarita (Alisma plantaooaouatica), troscotul de apa (Polygonum amphibium), piciorul cocosului de apa (Ranunculus aquatilis).
Datorita conditiilor specifice mediului acvatic (saracia in O2, CO2, temperatura mai putin variabila), aceste plante de apa reprezinta unele caractere comune, rezultat al adaptari lor la acest mediu. Astfel ele au tulpini de dimensiuni mici, sunt ierboase, epiderma nu este protejata de cuticula, dar prezinta celule cu cloroplaste, stomatele lipsesc, prezinta o dezvoltare mare a tesuturilor aerifere in scoarta, ceea ce contribuie la scaderea greutatii (densitatii) lor ce conduce la mentinerea unei pozitii verticale sau de plutire. Vasele conducatoare lemnoase sunt reduse, la fel si tesuturile mecanice.
2 Anatomia tulpinii
2.1. Structura primara
Cresterea in lungime a tulpinii se datoreste existentei si activitatii unor meristeme. Celule initiale din varful tulpinii si meristemele in activitate din aceasta zona se numesc varfuri vegetative, conuri vegetative sau conuri de crestere. Acestea adapostesc in primul lor strat celula sau celulele initiale prin diviziunea carora se formeaza meristemul primordial care contribuie la alungirea tulpinii si care vor da nastere la meristemele primare ce, la randul lor, vor genera tesuturile primare ale tulpinii.
Conform teoriei corpuscului si tunicii, stratul extern de meristem primar este denumit tunica si acopera meristemul primar central format din numeroase straturi de celule numit corpus. In varful vegetativ celulele tunicii se divid numai anticlinal pe cand la o anumita departare de varf se divid si periclinal. Celulele corpusului se divid de la inceput si periclinal si anticlinal. Exista insa si cazuri cand cele doua straturi nu pot fi strict delimitate sau ca numarul straturilor tunicii sa fie mai mare de unu, un tip unic de structura nu este valabil pentru toate speciile. Se poate insa preciza ordinea aparitiei tesuturilor primare: 1) fascicolele conducatoare ce apar din procambiu; 2) epiderma care ia nastere din tunica; 3) tesuturile fundamentale si 4) cele mecanice ce iau nastere din meristemul fundamental.
In structura primara a tulpinii sunt prezente tesuturi primare care isi au originea in meristemele primare ale tulpinii. Studierea acestor structuri se face pe sectiuni transversale, longitudinal-radiare si longitudinal tangentiale. Tesuturile primare ale tulpinii sunt aranjate concentric in 3 regiuni: epiderma, scoarta si cilindrul central.
Epiderma reprezinta tesutul protector al tulpinii si este formata din celule cu peretii bombati la exterior, cutinizati, lipsiti de cloroplaste si de substante de rezerva. Intre celulele epidermice se afla si stomate iar altele se transforma in peri.
Scoarta sau parenchimul cortical este cuprinsa intre epiderma si periciclu. Este alcatuita dintr-un numar mare de straturi de celule parenchimatice, cu pereti subtiri si celulozici, cu spatii intercelulare. In cazul tulpinilor tinere primele straturi ale scoartei contin cloroplaste si pot indeplini si functia de fotosinteza, iar straturile situate mai spre interior acumuleaza substante de rezerva. In straturile externe ale scoartei se mai pot diferentia si tesuturi mecanice, ca de ex. colenchim la Lamiaceae si Apiaceae, sclerenchim la Poaceae, fie sclereide izolate. La alte tipuri de tulpini scoarta poate contine tesuturi secretoare inclusiv celulele cu secretie intracelulara, celule secretoare izolate ca la dafin (Laurus nobilis), canale secretoare (la Conifere, Apiaceae), celule mucilagene (ca la Malvaceae). La Euphorbiaceae si Moraceae in scoarta apar laticifere.
Cel mai profund strat al scoartei, desi nu este asa de net diferentiat ca la radacina, constituie endodermul. La rizomi el apare ingrosat in forma de 'U', iar la altele apare ca un strat cu celule dispuse ordonat si avand continut de amidon din care cauza acest strat se mai numeste teaca amilifera. Exista cazuri cand endodermul nu este diferentiat asa cum se intampla la unele Monocotiledonate si la Ranunculus.
Cilindrul central sau Stelul cuprinde toate tesuturile situate la interiorul endodermului Majoritatea plantelor poseda in tulpina un singur cilindru central si se numesc monostelice. Unele ferigi si specii de Primula si Gunera prezinta mai multe cilindre centrale fiecare cu endoderm propriu, fenomen numit polistelie. In cazul altor specii (Equisetum limosum) tulpina nu are cilindru central (astel) fiecare fascicol conducator prezentand un endoderm propriu.
In cursul evolutiei filogenetice stelul a trecut prin diferite etape (Fig.126) de la protostel (o coloana centrala de lemn inconjurata de una de liber), actinostel (in care coloana centrala de lemn prezinta mai multe creste radiare), sifonostel (in partea centrala apare o coloana de maduva), dictiostel (cilindrii de liber si lemn prezinta intreruperi determinate de ramificatiile frunzelor si ramurilor), eustel intalnit la Gymnospermae si la majoritatea Dicotiledonatelor la care stelul este alcatuit din fascicole libero-lemnoase dispuse ordonat, sub forma unui cerc, cu parenchim medular (maduva) in partea centrala si de la care pornesc razele medulare ce ocupa spatiile dintre fascicole. La Monocotiledonate se intalneste un atactostel, format din numeroase fascicole libero-lemnose, dispuse neordonat, intr-un parenchim medular (Fig.128 A)
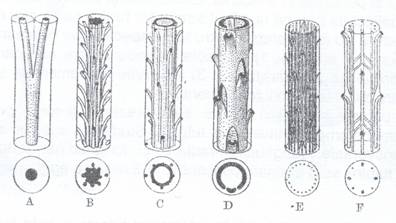
Fig.126. Tipuri de stel: A - Protostel; B - Actinostel; C - Dictiostel; E si F - Eustel.
Cilindrul central tipic, eustelul, este alcatuit din urmatoarele tesuturi: periciclu, fascicolele conducatoare libero-lemnoase, razele medulare si maduva.
Periciclul este situat sub endoderm si la exteriorul fascicolelor conducatoare. Este format din unul sau mai multe straturi de celule cu pereti celulozici dar uneori acestia se pot lignifica sau pot apare numai unele zone (pachete) de fibre periciclice, in alte cazuri periciclul poate chiar sa lipseasca.
Fascicolele conducatoare reprezinta cele mai importante tesuturi ale cilindrului central. In cazul tulpinii se intalnesc fascicole comune libero-lemnoase, cel mai adesea fascicol de tip colateral la care tesutul lemnos este dispus spre interior si cel liberian spre exterior. Dezvoltarea tesutului lemnos din fascicolele tulpinii este centrifug, protoxilemul fiind situat spre centrul tulpinii si metaxilemul spre exterior, in timp ce elementele liberului se dezvolta centripet, cu protofloemul spre exterior si metafloemul spre interior. Tesutul lemnos al fascicolelor este alcatuit din vase lemnoase si putin parenchim lemnos. Adesea fiecare fascicol este protejat de cate un arc de sclerenchim, cel dinspre partea externa (liberiana) fiind mai dezvoltat (Fig.127, 128).
Razele medulare primare reprezinta cordoane de celule parenchimatice care ocupa intervalul dintre fascicole si fac legatura intre maduva si periciclu.
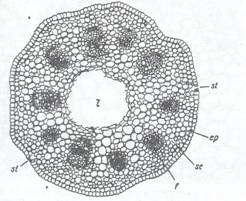
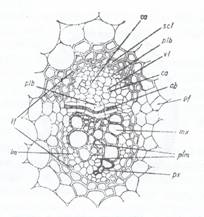
Fig.127. Structura primara a tulpinii de Ranunculus repens (sectiune transversala):
A - ansamblu; B - fascicul conducator marit; ca-celule anexe; cb-cambiu liberolemnos; cm-canal medular; cp-celule de pasaj; ep-epiderma; fil-fascicul libero-lemnos; lb-liber; lm-lemn; mx-metaxilem; pf-parenchim fundamental; plb-parencim liberian; plm-parenchim lemnos; pm-parenchim perimedular; pr-protoxilem; se-scoarta; scl-sclerenchim; st-stomate; tf-teaca fasciculara; vl-vase liberiene (tuburi ciuruite)
Structura primara a tulpinii de Dicotiledonate: Ranunculus repens (Fig.127) (pentru detalii a se vedea Caietul de lucrari practice). Primele straturi ale scoartei sunt usor colenchimatice, endodermul nu este diferentiat si se considera ca scoarta se termina odata cu celulele ce vin in contact cu fascicolele conducatoare. De asmenea nici cilindrul central nu este bine delimitat spre exterior, lipsind un periciclu diferentiat. Fascicolele conducatoare sunt dispuse pe un singur cerc, sunt de tip colateral deschis avand un cambiu fascicular intre liber si lemn (acest cambiu reprezinta cateva straturi de celule care si-au pastrat caracterul embrionar, alcatuind deci un cambiu fascicular). Intre fascicole se gasesc razele medulare primare, iar centrul tulpinii este ocupat de o lacuna medulara rexigena.
La Monocotiledonate (Zea mays) (Fig.128), epiderma este formata din celule mici, cu peretii externi puternic impregnati cu bioxid de siliciu, prezentand insa si stomate. Scoarta prezinta straturi cu celule lignificat care constituie o hipoderma.
Scoarta nu poate fi delimitata spre interior. Spatiul situat sub hipoderma este format dintr-un parenchim fundamental care contine numeroase fascicole liberolemnoase de tip colateral inchis, imprastiate fara ordine in acest parenchim. Fascicolele situate spre exterior sunt mai mici si complet inconjurate de sclerenchim, iar fascicolele dispuse spre partea centrala a tulpinii sunt mai mari iar tecile de sclerenchim nu se unesc prin marginile lor. Tesutul lemnos al fascicolului este dispus in forma de 'V', avand protoxilemul format din 1-2 vase cu diametrul mai mic cuprinse intr-o lacuna lisigena, aceste vase fiind dispuse in varful 'V'-ului. Metaxilemul este situat spre exterior si este alcatuit din doua vase cu diametrul mare, dispuse indepartate unul de altul:
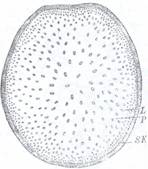
Fig.128 A. Structura primara a tulpinii de Zea mays A - schema sectiunii transversale;
I - facicol conducator; P - parenchim fundametal.
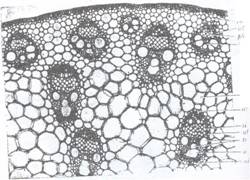
Fig.128 B. portiune din sectiunea transversala prin tulpina vazuta la microscop,
ep-epiderma; h- hipoderma; Co-scoarta; scl-sclerenchim; ph - floem; x-Xilem; par-parenchim medular; si-vase liberiene; tr-traheide.
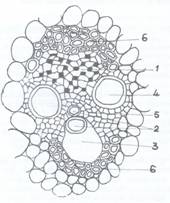
Fig C. Structura unui fascicol marit:
1 - tesut liberian; 2 - protoxilem; 3 - lacuna lisigena; 4 - metaxilem;
5 - parenchim lemnos; 6 - arcuri de sclerenchim
Cresterea in grosime a tulpinilor cu structura primara, adica a tulpinilor unor plante ierboase si ale unor monocotiledonate lemnoase lipsite de zone generatoare (meristeme) secundare cresc in grosime prin cresterea numarului celulelor si prin marirea volumului acestora. La Monocotiledonate apar si functioneaza temporar unele mantale meristematice intre tunica si corpus, in zona varfului vegetativ si, prin diviziuni anticlinale si periclinale, produc cresterea in grosime a tulpini lor.
Variatiuni in structura primara a tulpinii
Fata de structurile primare ale tulpinii, descrise la caracterizarea generala, la Dicotiledonate si Monocotiledonate, exista plante care se abat in privinta structurii primare de la aceste tipuri principale si constituie variatiuni.
La Pteridofite structura primara a tulpinii reflecta pozitia sistematica inferioara a acestei Increngaturi fata de Gymnosperme si Angiosperme. Unele prezinta cilindrul central in forma de protostel, actinostel (Psilotum triquetrum) sau sifonostel (Marsilia quadrifolia). La multe Pteridofite fascicolele prezinta o dispozitie radiara, la care lemnul se dezvolta centripet la fel ca la radacina Polistelia este, de asemenea, un fenomen frecvent intalnit, la fel si structura asimetrica a tulpinii.
La Dicotiledonate un exemplu de variatiune in structura tulpinii se intalneste la planta Aristolochia sipho, (Fig.129, 130) care prezinta urmatoarele caractere diferentiale: scoarta prezinta doua regiuni distincte: una externa, colenchimatica si una interna, parenchimatica ce alcatuieste un adevarat clorenchim, cu celule alungite radiar in care sunt prezente cloroplaste si care dau culoarea verde tulpinilor tinere. Endodermul este bine diferentiat, cu celule ce contin amidon si poarta denumirea de teaca amilifera. Periciclul este pluristratificat si prezinta, de asemenea, doua zone distincte: una externa sclerenchimatica si una interna, parenchimatica, ambele avand forma unor arcuri de cerc spre interior ce inconjoara fascicolele conducatoare. Fascicolele conducatoare sunt de tip colateral deschis si sunt lipsite de arcuri de sclerenchim proprii. Cambiul fascicular se continua si cu cambiul interfascicular, formand astfel un inel continuu.
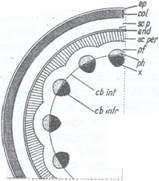
Fig. 129: Schema structuri; tulpinii de Aristolochia sipho
ep - epiderma; col - scoarta colenchimatica; sc. p. - scoarta parenchimatica; end - endoderm; sc. per. - periciclu sclerenchimatic; pf - periciclu parenchimatic; ph - liber, x - lemn; cb. int. cambiu interfascicular; cb. intr. - cambiu intrafascicular; m- maduva
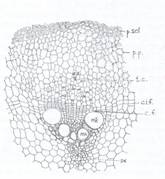
Fig.130. Sectiune transversala printr-un fascicol libero-lemnos de Aristolochia;
p.scl.- periciclu sclerenchimatic; p.p. - periciclu parenchimatic; c.an. - celula anexa; te. - tub ciuruit; c.f. - cambiu fascicul ar; c.i.f. - cambiu interfascicular; mx-metalixelm, px-protolixem
La Monocotiledonate vom studia ca exemplu de variatiune structura primara a tulpinii de Convallaria maialis (Iacramioara) (Fig.131, 132), in structura primara a unui rizom se pot observa celulele endodermului ingrosat in forma de potcoava, cu lignina. La un rizom in varsta se observa o ingrosare completa, lignificata si a stratului de celule care alcatuieste periciclu. In cilindrul central sunt prezente 2 tipuri de fascicole libero-lemnoase; la periferie se gasesc fascicole colaterale deschise care se sprijina cu liberul pe periciclu iar vasele lemnoase sunt dispuse in forma literei 'V', cu deschiderea spre periciclu. In zona centrala se gasesc insa fascicole de tip leptocentric (sau amfivasal), cu liberul situat la interior si inconjurat de lemn.
Tulpinile plantelor acvatice se caracterizeaza prin lipsa stomatelor din epiderma, dar celulele epidermei pot prezenta cloroplaste. Scoarta acestor tulpini prezinta tesuturi aerifere formate din lacune mari cu aer, iar cilindrul central este redus fata de scoarta, vasele conducatoare lemnoase se gasesc in numar redus.
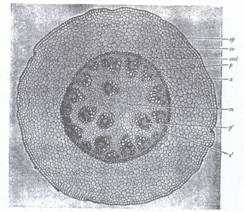
Fig.131. Sectiune transversala prin rizomul de lacramioara (Convallaria majalis); ep epiderma; coscoarta; end-endoderm; p-liber si x-lemn din cadrul fascicolelor colaterale; p si x-liber si lemn din fascicolele leptocentrice; m-parenchim medular.
2.3. Structura coletului
Trecerea de la structura primara a radacinii (cu fascicole separate) la cea a tulpinii (cu fascicole mixte) are loc la nivelul coletului. Aceasta trecere se realizeaza dupa 3 tipuri: (Fig.133)
1. Tipul Mirabilis, la care fascicolele liberiene din radacina isi continua traiectul prin colet si tulpina, iar fascicolele lemnoase se dubleaza, se rasucesc cu 180 si se aseaza cate 2 jumatati diferite la cate un fascicol liberian, alcatuind in tulpina fascicole comune libero-lemnoase. In acest caz numarul fascicolelor formate in tulpina este egal cu al celor liberiene sau lemnoase din radacina.
2. Tipul Cucurbita este mai frecvent intalnit si la acesta se dedubleaza atat fasciculele liberiene cat si cele lemnoase si, dupa o rotire de 180, fiecare jumatate de fascicol lemnos se dispune spre interiorul unei jumatati de fascicol liberian. In acest mod numarul total al fascicolelor libero-lemnoase din tulpina este dublu fata de cel al fascicolelor liberiene sau lemnose; separat, din radacina.
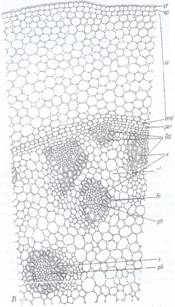
Fig.132. Sector din sectiunea transversala in rizomul de Convallaria majalis; ct - cuticula;
ep - epiderma; sc - scoarta; end-endoderm; per - periciclu; f.c. - fascicol colateral; f.1. - fascicol
leptocentric; x - xilem; ph - floem; pf - parenchim. fundamental.
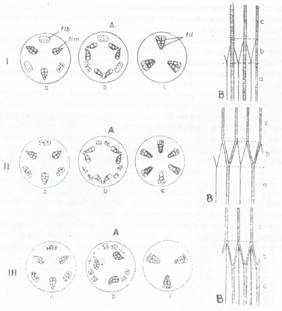
Fig.133, Reprezentarea schematica a trecerii fasciculelor conducatoare din radacina in tulpina: 1Mirabilis jalapa; II-Cucurbita pepo; III-Pisum sativum; A-sectiuni transversale; B-sectiuni longitudinale; a-sectiune in radacina; b-sectiune in colet; c-sectiune in tulpina; flb-fascicul de liber; flm-fascicol de lemn; fil-fascicol libero-lemnos.
3. Tipul Pisum este invers tipului Mirabilis, in acest caz se dedubleaza fascicolele liberiene din radacina, iar cele lemnoase se rotesc doar pe loc cu 180, dupa care jumatatile liberiene se dispun cate doua la exteriorul fascicolelor lemnoase intregi. Si in acest caz numarul noilor fascicole libero-lemnoase din tulpina este egal cu cel al fascicolelor liberiene sau lemnoase, separat, din radacina.
In concluzie, trecerea de la structura primara a radacinii la structura primara a tulpinii consta in schimbarea progresiva a dispunerii in alternanta a fascicolelor liberiene si lemnoase la o dispunere in comun a fascicolelor conducatoare liberolemnoase in tulpina.
2.4. Structura secundara a tulpinii
In tulpina Gymnospermelor si Dicotiledonatelor lemnoase, cu o crestere anuala in grosime, apar si functioneaza meristemele secundare care determina cresterea diametrului tulpinii. Tesuturile primare, epiderma si scoarta in primul rand, nu-si mai pot indeplini functiile in noile conditii, ele se rup si se exfoliaza si vor fi inlocuite cu tesuturi noi, secundare, nascute din meristemele secundare. Tesuturile secundare se depun peste cele primare, fara a le inlatura, tulpina capatand astfel o noua structura, secundara.
Tesuturile meristematice secundare ale tulpinii, numite si zone generatoare, sunt cambiul si felogenul. Ele au o pozitie laterala, forma cilindrica, iar in sectiune transversala sunt de la inceput circulare, din care cauza si tesuturile secundare care iau nastere din aceste meristeme prezinta aspect inelar, ca niste mansoane continue, concentrice (Fig.134).
Cambiul sau zona generatoare libero-lemnoasa este situat totdeauna in cilindrul central, intre partea liberiana si lemnoasa a fascicolelor primare. La tulpinile cu fascicole libero-lemnoase de tip bicolateral (Solanaceae, Cucurbitaceae) cambiul intern va produce spre interior liber perimedular. La cartof in structura tuberculului predomina elementele de parenchim liberian secundar perimedular. Celulele cambiului sunt lungi, prozenchimatice, de forma prismatica turtita, sunt celule vii si foarte active, se divid in sens tangential, dand nastere, alternativ, la celule fiice atat spre interior cat si spre exterior, totusi se produc un numar mai mare de celule spre interior. Celulele care iau nastere din cambiu sunt dispuse in serii radiare si se diferentiaza pe masura indepartarii lor de cambiu astfel: cele de la exterior se transforma in elemente de liber secundar, iar cele dinspre interior in elemente de lemn secundar. Celulele cambiului situate in dreptul fostelor raze medulare primare raman parenchimatice, cu pereti celulozici, alungite in directie radiara si vor forma razele medulare secundare. Mai rar celulele cambiului se divid insa si in sens radiar, cambiul creste in grosime, proces cunoscut sub numele de dilatatia cambiului.
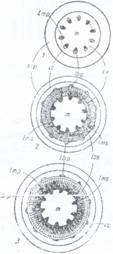
Fig.134. Formarea inelelor anuale in tulpina dicotiledonatelor:
1 - structura primara; 2 - structura tulpinii la sfarsitul anului intai; 3 - la sfarsitul anului al doilea;
cv - cambiu vascular; cc - limita dintre scoarta si clindrul central; lbp - liber primar; lbs - inele cu liber secundar; lmp - lemn primar; lms - inele de lemn secundar; m - maduva; scp - scoarta primara.
Celulele cambiului raman vii in tot cursul vietii plantelor, activitatea lor insa se desfasoara cu intermitenta. Astfel in zona temperata activitatea cambiului incepe primavara si se termina toamna. Prin producerea, an de an, de tesuturi secundare in forma de mansoane, sau circulare in sectiune transversala, care se suprapun peste tesuturile primare, cambiul inlocuieste fosta structura primara a cilindrului central, cu una secundara, caracteristica. Astfel partea lemnoasa a fascicolelor libero-lemnoase primare este impinsa tot mai spre interiorul tulpinii, pe cand partea liberiana este presata in afara, iar cu timpul tesuturile primare nu mai pot fi identificate.
Spre interior cambiul produce lemn secundar, care reprezinta cea mai insemnata parte din aparatul vegetativ al Gymnospermelor si Angiospermelor lemnoase. In structura lemnului secundar se disting urmatoarele elemente: traheide, trahei, fibre lemnoase si parenchim lemnos, raportul cantitativ intre aceste elemente variaza foarte mult, in functie de specie (Fig.135).
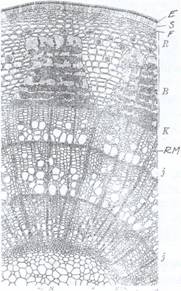
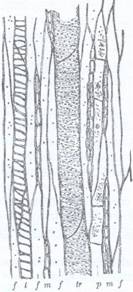
Fig.135A. Structura secundara a tulpinii la o ramura B. Structura lemnului secundar de tei: f - fibre de tei (Tilia) de 3 ani, s - suber secundar; R - lemnoase; t - traheide; m - raze medulare; tr scoarta; B - liber secundar cu palnii de dilatare; K - -trahei; p - parenchim lemnos (stg) cambiu; J - inele anuale; E - epiderma; f - felogen; RM - razele medulare secundare
Traheidele formeaza aproape intreaga masa lemnoasa la Gymnosperme. Sunt celule lungi de 2-10 mm, de regula ascutite la capete cu peretii puternic lignificati si cu punctuatiuni areolate. Pe langa rolul de conducere traheidele indeplinesc si un rol mecanic.
Traheele se intalnesc numai in lemnul Dicotiledonatelor. Ele prezinta ingrosari de tip punctat la maturitate, dar mai pot prezenta si punctuatiuni areolate si semiaerolate si servesc la conducerea sevei brute. Forma lor poate fi cilindrica, umflata la mijloc sau prismatica. Sectiunea lor transversala, in forma de cerc sau poligon, se numeste por, diametrul lor este mai mare in lemnul timpuriu sau 'de primavara
Fibrele lemnoase (sau libriforme) sunt celule lungi, ascutite la capete (fuziforme), cu pereti puternic ingrosati, cu lumen mic, prezentand punctuatiuni simple, mici si oblice, in forma de butoniera si numai foarte rar punctuatiuni areolate. Fibrele lemnoase se intalnesc numai in lemnul Dicotiledonatelor unde constituie masa principala a lemnului, rolul lor fiind, in primul rand unul mecanic.
Parenchimul lemnos este constituit din celule vii in care se depun substante de rezerva, in special amidon si lipide. Aceste elemente iau nastere din unele celule ale cambiului situate spre interior, care cresc activ in directia longitudinala si apoi se divid prin pereti transversali formand 'coloane' de celule scurte, suprapuse. Substantele de rezerva depuse in acest tesut vor fi folosite primavara, dupa o prealabila hidroliza a lor, de catre mugurii care se deschid sau de catre cambiul care isi va relua activitatea. Unele specii lemnoase (tei, mesteacan, salcam), pot prezenta, pe langa parenchimul obisnuit si unul de forma fibrelar, fuziform, dar viu, cu peretii subtiri, care poarta numele de parenchim lemnos fusiform. In parenchimul lemnas se pot intalni canale rezinifere, cum este cazul la tulpina coniferelor.
Liberul secundar ia nastere din
celulele rezultate spre exterior in urma diviziunii cambiului. Grosimea
liberului secundar este mai redusa in
comparatie cu a lemnului secundar din cauza producerii mai multor celule
spre interior de catre cambiu (un raport de cca
Zona liberului secundar, impreuna cu cea de periderm (scoarta secundara), nascuta din activitatea felogenului este numita adesea coaja si, impropriu chiar scoarta, constituind partea tulpinii cu utilizare farmaceutica, denumita 'cortex'.
Liberul secundar contine, din punct de vedere histologic, 4 tipuri de elemente: tuburi ciuruite, celule anexe, parenchim liberian si fibre liberiene.
Tuburile ciuruite sau vasele liberiene reprezinta singurele elemente conducatoare ale liberului prin care circula seva elaborata. Sunt celule vii, scurte dar largi, cu pereti subtiri si celulozici, cei terminali fiind oblici si cu placi ciuruite multiple.
Parenchimul liberian se aseamana cu cel lemnos atat in ce priveste forma car si functia.
Fibrele liberiene secundare sunt celule fuziforme, cu peretii puternic ingrosati cu lignina, cu punctuatiuni simple, rare si oblice, lumenul foarte mic si joaca un important rol mecanic. La unele plante lemnoase (dud, tei) tesutul mecanic din liberul secundar este alcatuit numai din fibre, pe cand la stejar, frasin este format din fibre si sclereide izolate sau in grupuri, iar la fag si molid numai din sclereide. La tulpina de in (Fig.136) si canepa (Fig.137) fibrele liberiene reprezinta fibrele textile.
Celulele anexe apar numai in liberul Angiospermelor si au aceeasi origine cu tuburile ciuruite (provin din aceeasi celula prin diviziune longitudinala) si reprezinta, o rezerva a acestora, ele putandu-se transforma in tuburi ciuruite.
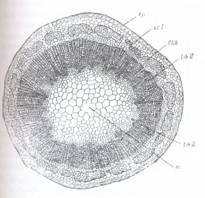
Fig.136. Sectiune transversala prin tulpina de in: ep - epiderma; sc l - scoarta primara; f.lib. - fibre liberiene; lib II - liber secundar; lm 11- lemn secundar; m - maduva.

Fig.137. Portiune dintr-o sectiune transversala prin tulpina de canepa: a - epiderma; b - parenchim clorofilian; c - endoderm; d - fibre liberiene primare; e - parenchim; f - fibre liberiene secundare; 9 - liber secundar; h - cambiu; 1- portiune intralemnoasa a razei medulare; k - lemn secundar.
In liberul secundar se pot intalni si celule si tesuturi secretoare, ca la dafin, arborele de scortisoara sau lactifere ca la unele plante din fam. Moraceae. Liberul format numai din elemente vii (tuburi ciuruite, celule anexe, parenchim liberian) se numeste liber moale, iar cel care contine si elemente mecanice poarta numele de liber tare.
Razele medulare secundare sunt produse de cambiu si dispuse sub forma unor benzi radiare (Fig.138). Ele servesc la schimburile gazoase si conducerea sevei in sens radiar (transversal) prin tulpina, precum si la depozitarea substante lor de rezerva. Razele medulare secundare iau nastere, mai ales, din zona interfasciculara a cambiului. Ele se intind de la maduva pana la scoarta secundara, fiecare raza avand o parte intralemnoasa si una intraliberiana, care se continua fara intrerupere. Razele medulare secundare sunt formate din celule parenchimatice, dreptunghiulare, alungite in directia razei tulpinii. Celulele sunt vii, cu peretii foarte usor lignificati, cu spatii intercelulare, iar prin intermediul lenticelelor comunica cu mediul exterior. Razele medulare au urmatoarele dimensiuni caracteristice: lungimea, care se masoara in directie radiara, pe sectiune transversala; latimea (sau grosimea) se masoara in directie tangentiala. Latimea creste prin alaturarea mai multor raze (raze compuse); Inaltimea se masoara in directia longitudinala si apare pe sectiune longitudinal-radiara. Aspectul caracteristic al razelor medulare pe diferite sectiuni, impreuna cu aspectul inelelor anuale dau desene variate ale lemnului si permit recunoasterea acestuia.
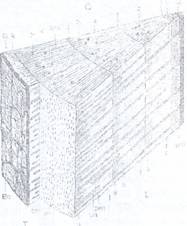
Fig.138. Sectiune transversala (Q) longitudinal radiara (L) si longitudinal - tangentiala (T) in tulpina de pin de 4 ani; M - maduva; I - 4 inele anuale; pm - raze medulare primare; sm - raze medulare secundare; f - lemn de pimavara; s - lemn de toamna; h - canale rezinifere; k - cambiu; b - liber secundar.
Inelele anuale (Fig.1390. Primele straturi de lemn secundar dintr-un inel anual care s-au format primavara si la inceputul verii reprezinta vase lemnoase cu diametrul mare si cu peretii mai putin ingrosati si prin care circulatia sevei brute, abundenta in aceasta perioada, se face usor. Acesta se numeste lemn de primavara. Spre sfarsitul verii si toamna, cambiul produce vase cu diametrul mic si cu peretii din ce in ce mai ingrosati, cu o proportie tot mai mare de fibre lemnoase si acesta reprezinta lemnul de toamna. La cele mai multe esente lemnoase de la noi lemnul de primavara este inelo-poros prezentand vase cu diametrul mult mai mare decat cele din lemnul de toamna si astfel inelele sunt usor de identificat (stejar, castan, salcam, frasin). La alte esente exista un lemn difuz-poros la care vasele au diametre egale pe toata grosimea unui inel anual, ca la mesteacan, plop, artar si la care inelele anuale pot fi identificate numai la baza predominantei elementelor mecanice.

Fig.139. Inele anuale la salcam: A - alburn; B - duramen
Alternanta lemnului de primavara cu cel de toamna permite identificare inelelor anuale si, pe aceasta baza, stabilirea varstei tulpinii si a arborilor. La arborii tropicali, din zona lipsita de anotimpuri, variatiile in structura lemnului secundar sunt determinate de perioadele de ploi si seceta care se succed ritmic in cursul unui an si, in acest caz inelele nefiind anuale, nu indica varsta plantelor. In zonele in care nu apar diferente nici in privinta precipitatiilor, nu apar nici inele anuale, iar activitatea cambiului se desfasoara uniform tot timpul anului. Grosimea inelelor anuale difera cu specia, esentele moi (salcia, plopul) prezinta inele mai groase decat cele tari (fag, stejar). Chiar la aceeasi specie putem intalni inele mai groase sau mai inguste in functie de cresterea mai lenta sau mai intensa pe care a suferit-o copacul in anii respectivi, de variatiile precipitatii lor.
Felogenul sau zona generatoare subero-felodermica reprezinta al doilea meristem secundar intalnit frecvent in cazul tulpinilor lemnoase. Acesta va da nastere tesuturilor de aparare secundare care inlocuiesc epiderma. Felogenul este format dintr-un singur strat de celule, dispuse inelar, care cresc in directie radiara si se divid tangential producand celule noi atat pe o parte cat si pe alta. La fel ca si cambiul activitatea sa este ritmica, incetandu-si activitatea toamna si reluandu-o primavara, dar mai tarziu decat cambiul. Pozitia felogenului este situata intotdeauna la exterior de cambiu si anume: in epiderma la salcie si par, in primul strat al scoartei la fag, tei si mesteacan, in endoderm sau chiar in periciclu ca la coacaze, strugurasi, vita de vie. Cu cat felogenul va fi situat mai spre interior cu atat mai multe tesuturi primare si apoi secundare, ramase in afara acestuia, mor din cauza suberului secundar, nemaiprimind hrana si se vor exfolia spre exterior.
Felogenul produce spre exterior suber secundar, iar spre interior feloderm (sau scoarta secundara) (Fig.140).
Pe sectiune transversala suberul secundar se prezinta sub forma de celule tabulare, moarte, turtite tangential, dispuse in serii radiare si strans unite intre ele (Fig.141).
pap fe pas st sm
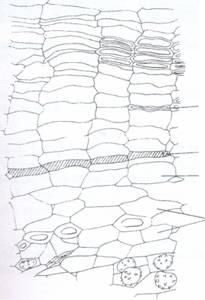
Fig.140. Sectiune transversala printr-o ramura de Ribes rubrum; fe - felogen; p.a.p.- parenchim primar; pas - parenchim secundar (feloderm); sm - suber moale; st - suber tare.
Fig.141. Straturi succesive de suber secundar (K1-K3) din structura secundara a tuipinii alcatuind
ritidomul. S - celule pietroase; pR - feloderm

Fig.141.A Ritidom persistent la nuc Fig.141.B Ritidom inelar Fig.141.C Ritidom solzos
la mesteacan (sau in placi) la molid

Fig.141.D Ritidom fibros la vita de vie.
Felodermul este un tesut secundar viu, care se deosebeste de scoarta primara prin dispozitia celulelor in serii radiare regulate. Celulele sale pot contine cloroplaste, mai ales la tulpinile tinere, apoi substante de rezerva si uneori poate ingloba si tesuturi mecanice, colenchim sau sclerenchim.
Suberul secundar, felogenul si felodermul alcatuiesc impreuna peridermul sau scoarta secundara. Spre deosebire de cambiu felogenul are o viata scurta, dupa cativa ani de functionare sau chiar dupa un an el moare si va fi inlocuit de un alt felogen s.a.m.d. Noile zone generatoare subero-felodermice se formeaza tot mai spre interior, chiar pana la liberul primar sau secundar si vor determina moartea tuturor tesuturilor situate la exteriorul sau si in final se vor exfolia. Ansamblul heterogen alcatuit atat din tesuturi primare cat si secundare moarte si exfoliate la exteriorul tulpinilor batrane ale arborilor poarta denumire de ritidom (Fig.141). Formarea ritidomului incepe cu atat mai repede cu cat el a fost situat mai la interior. Ritidomul poate fi, persistent ca la stejar, nuc (Fig.141 A) si poate ramane atasat de tulpina toata viata sau alteori este trecator si se exfoliaza sub forma de inele ca la mesteacan (Fig.141 B) si cires sau sub forma de placi (ritidom solzos) ca la pin, platan (Fig.141C) si fibros ca la vita de vie (1410).
Din loc in loc felogenul produce spre exterior, in loc de suber celule rotunde parenchimatice cu spatii intercelulare numite lenticele (Fig142) si care asigura schimbul de gaze intre tesuturile conducatoare, scoarta si exterior. Asemenea lenticele se pot observa si cu ochiul liber la soc, mesteacan, cires.
Kl K f B A
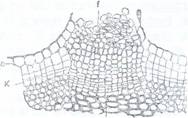
![]()
Fig.142. Lenticele din tulpina de soc (Sambucus nigra) vedere macroscopia (B) si in sectiune transversla prin dreptul lenticelei (A), K - felogen; Kl - felogen la nivelul lenticelei; f - parenchim; eepiderma.
Tulpina ca sursa de medicament
Tulpina ofera, intre organele vegetale, variate si numeroase produse vegetale medicinale cu importante utilizari practice si continut in principii active.
In Farmacopeea Romana ed. X-a sunt inluse urmatoarele 6 produse medicinale vegetale oficinale:
1. Chelidonii herba, parte aeriana de rostopasca (Chelidonium majus) sau negelarita, contine alcaloizi cu proprietati antispastice si colagoge utilizata in afectiuni ale vezicii biliare.
2. Equiseti herba reprezinta partea aeriana a speciei Equisetum arvense - coada calului cu continut de ulei volatil, flavonoide, saruri minerale (Si02) si utilizat ca un bun diuretic.
3. Hyperici herba este iarba de sunatoare, Hypericum perforatum, recoltata in timpul infloririi.Contine flavonoide, ulei volatil, hipericina, cu actiune de vitamina P, cicatrizanta, antiseptica indicata in arsuri.
4. Thymi herba iarba de cimbru de cultura (Tymus vulgaris) cu continut de ulei volatil bogat in fenoli (timol) si actiune antiseptica si coleretica.
5. Frangulae cortex, scoarta de crusin, Rhamnus frangula cu continut de oximetilantrachinone avand proprietati laxative sau purgative in functie de doza. Nu se va folosi scoarta proaspata care produce efecte iritante.
6. Valerianae rhizoma cum radicibus reprezinta rizomii si radacinile de odolean, Valeria officinalis, cu continut de ulei volatil si iridoide fiind unul din produsele de traditie a fitoterapiei pentru efectele sedative, indicat in insomnii, nevroze.
Alte produse vegetale medicinale neoficinale dar foarte mult cautate si apreciate pentru proprietatile lor sunt: Adonidis herba - iarba de ruscuta de primavara (Adonis vernalis) cu continut de glicozide carditonice, Absinthi herba - iarba de pelin (Artemisia absinthium) cu continut de principii amare, ulei voiatil si folosit ca tonic amar si la prepararea bauturilor alcoolice (bitter, vermut) Centauri herba - iarba de fierea pamantului (Centaurium umbellatum) cu continut de glicozide amare utilizat ca tonic amar, Chinae cortex reprezinta scoarta arborelui Cinchona succirubra (fam. Rubiaceae) originar din America de Sud. Contine alcaloizii chinina si chinidina cu actiune antimalariana si antipiretica, respectiv de normalizare a ritmului cardiac. Se utilizeaza si sub forma de extracte ca amaro-tonic in doze mici, Serpylli herba - iarba de cimbrisor de camp (Thymus serpyllum) cu continut de ulei volatil si flavone avand proprietati antitusive, antispastice, Berberidis cortex scoarta de dracila (Berberis vulgaris) cu continut de alcaloizi (berberina, berbamina, oxiacantina) si efecte coleretice si colagoge, indicat in afectiuni hepatobiliare, Querci cortex, scoarta de stejar (Quercus robur) cu continut ridicat de tanin si indicata ca astringenta. Populi gemmae, mugurii (nedeschisi) de plop negru (populus niara) cu continut de rezine, flavonoide si glicozide fenolice, indicati ca antihemoroidali, antiinflamatorii si dezinfectanti. Pini turiones mugurii de primavara deschisi de la pin (Pinus silvestris), cu continut de ulei voiatil si rasini, indicati in afectiuni respiratorii. Rhei rhizoma reprezinta rizomii si radacinile decorticate de revent, Rheum palmatum, si Rh. officinale cu continut de derivati antrachinonici si proprietati laxative si purgative. Gei rhizoma, rizomii de cerentel cu continut de tanin si derivati fenolici avand actiune astringenta si dezinfectanta provin de la Geum urbanum. Aconiti tuber, reprezinta tuberculii de omag (Aconitum napelus) cea mai toxica planta de la noi, cu continut de alcaloizi (aconitina), se utilizeaza la prepararea unor medicamente antitusive. Scillae bulbus reprezinta bulbii de ceapa de mare (Scilla maritima) cu continut de glicozide cardiotonice.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 24550
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved