| CATEGORII DOCUMENTE |
| Demografie | Ecologie mediu | Geologie | Hidrologie | Meteorologie |
Intreaga natura (vie si nevie) sunt organizate in corpuri materiale (atomi, molecule, celule, organe, organisme, planete, galaxii etc). Prin organizare se intelege un ansamblu de fenomene si elemente cu interactiuni si functii in cadrul ansamblului si care se subordoneaza functiilor intregului.
Orice organizare implica: structura (existenta relatiilor spatiale si interactiunile dintre elemente) si functie (activitatea desfasurata de un element component ce se subordoneaza functionalitatii intregului).
Unitatea organizatorica a materiei vii este sistemul. Stugren (1975) arata ca Universul nu este o ingramadire intamplatoare de particole ci este ceva foarte ordonat - un sistem (un complex de procese sau elemente care se afla intr-o interactiune permanenta).
Din punctul de vedere al relatiilor cu mediul exterior exista 3 categorii de sisteme:
izolate (nu schimba nici substanta nici energie cu exteriorul);
inchise (schimba cu exteriorul numai energie; trebuie precizat insa ca sisteme complet inchise nu exist adica pe langa energie se va schimba intotdeauna si substanta);
deschise (schimba cu exteriorul si substanta si energie); aici intra cele mai multe sistem, atat biologice cat si cele fara de viata (lac, rau, cadavru, stanca etc). Sistemul biologic este asadar un sistem deschis dar pe langa substanta si energie el schimba cu exteriorul si informatii (schimbul informational fiind cel de-al 3-lea tip de interactiuni in lumea vie). Teoriile moderne arata ca sistemul biologic este un sistem deschis automat, autoreglabil, informational, integral, dinamic, antientropic, autocinetic, teleonomic, autoreproducator, nelinear, oscilator, anizotrop, cu retroactiune etc.
Ca sistem, materia vie ni-se prezinta sub forma unor organisme, a unor"indivizi". Pe de alta parte, biosfera are un caracter "discret" (discontinuu) alcatuita din unitati distincte. Mintal sau artificial, aceste sisteme capabile de viata autonoma pot fi descompuse in unitati inferioare - subsisteme - . Iata o asemenea "descompunere" cu ierarhizarea subsistemelor: atom - molecula - macromolecula - particola coloidala - organit celular - celula - tesut - organ - sistem - aparat - arganism - populatie - specie - biocenoza - ecosistem - biosfera.
Aceasta ierarhizare defineste constituirea unor nivele de integrare (Fig. 30).
1. Clasifiacrea animalelor. Istorie si filiatie
O privire de ansamblu asupra acestui aspect poate fi imaginata azi conform datelor din Fig. 31
A. Regnul Monera (Procariota)
Include: bacterii fototrofe (Cyanobacterii, Rhodobacterii, Chlorobacterii), bacterii chimiotrofe (chimiolitotrofe, chimioorganotrofe).
Termenul de procariot (Chattoy) subliniaza aspectul particular al celulei bacteriene, in opozitie cu cele eucariote. Prin anii 1950 s-a evidentiat ca aceste celule nu au nucleu iar sistemul de membrane inchise este absent aproape cvasitotal. Structural, au un perete intotdeauna rigid (asigurat de peptidoglican). Gramm (1884) descopera o metoda de colorare si conform cu aceasta el denumeste bacteriile: gram pozitive (G+)(se coloreaza in violet) si gram negative (G-)(se coloreaza in roz).
Intre cele doua tipuri de bacterii existe diferente structurale semnificative (vezi Lucrari practice):
- prezent la G- si absent la G+: membrana lipoproteica si lipopolizaharidica externa, carboxizomii, perii, graunciorii de glicogen granulele de polifosfat;
- prezent la G+ si absent la G-: vacuolele cu gaz, epizomii, peretele celular.
Rolul celulei procariote. Importanta bacteriilor
- Rol important in dinamica ciclului carbonului (C ) si azotului (N2).
- 99-100% din aerul actual revine O2 si N2 liber, care la origine este produs de procariote, singurele implicate in schimbul de azot gazos cu atmosfera.
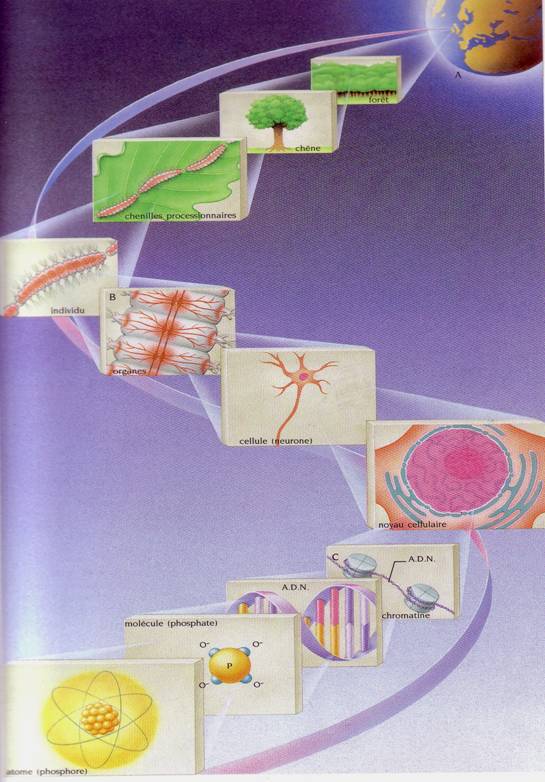
Fig. 30. Nivele de organizare, integrare si ierarhizare in biologie si ecologie (dupa Larousse, 1999).
- Carbonul (C) este elementul esential al materiei vii (toate proteinele au carbon in scheletul lor), iar toate formele de carbon (carbonati, gaz carbonic, bicarbonati) fac parte dintr-un ciclu vast in care C se va reintoarce la forma minerala (anorganica) si poate fi asimilat de catre plante.
- Rol important il au si bacterile fotosintetice (cyanobacteriile = alge albastre), insa exista si bacterii nefotosintetice caresunt capabile sa fixeze C, aceasta fiind singura activitate biologica responsabila de eliberarea oxidului de carbon (CO) si metanului (CH4) in atmosfera; anumite bacterii si toate cyanobacteriile, libere sau in simbioza cu diferite plante, reduc azotul atmosferic la amoniu iar acesta serveste la sinteza compusilor azotati ai materiei vii (in special acizii aminici).
- Numai bacteriile pot oxida amoniacul in nitrit (NO2) si in nitrat (NO3) in procesul denumit nitrificare.
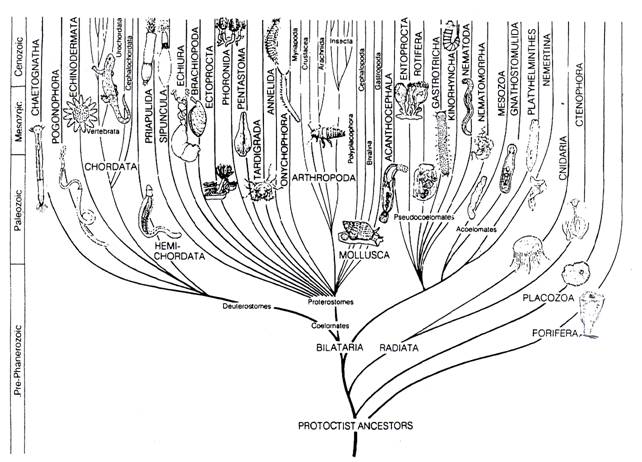
Fig. 31. Istorie si filiatie la nivelul regnului Animalia.
Caracteristici principale ale unor grupe de procariote
a. Bacteriile fotosintetice. Utilizeaza lumina ca sursa de energie. Sunt: cyanobacterii, rhodobacterii si chlorobacterii. Bacteriile fototrofe au aparat fotosintetic provenit din membrana plasmatica:
cyanobacteriile (alge albastre) au fost considerate mult timp ca apartinand numai regnului vegetal dar ele reprezinta toate caracteristicile organismelor procariote. Unele sunt unicelulare dar altele formeaza siruri de celule - alge filamentoase = trichome. Deci este posibil sa fie pluricelulare ?. In aceasta situatie este genul Rivularia. Aici apare pentru I-a data o specializare celulara (o asemenea structura pe care o vom reaminti atunci cand vom discuta ipoteza pluricelularitatii, este constituita din 3 tipuri de celule: heterociste (celule mari, putin pigmentate, incapabile de fotosinteza, specializate in reducerea azotului atmosferic), akinete (celule mari, voluminoase, pigmentate, bogate in rezerve) si necridii sau disjunctorii (celule care se gelifica si fragmenteaza trichoma in segmente denumite hormogonii);
- rhodobacteriile (bacterii rosii); sunt anaerobe; ele utilizeaza hidrogenul sulfurat (H2S) eliminand sulf (S) si oxigen (O2);
- chlorobacteriile (bacterii verzi ; utilizeaza H2S dar sulful ramane in afara celulei si asimileaza gaz carbonic;
b. Bacteriile chimiotrofe. Utilizeaza compusi chimici de natura minerala sau organica, pentru a elabora proprii lor compusi organici,
c. Bacteriile chimioorganotrofe. Reprezinta marea parte a eubacteriilor; sunt folositoare deoarece intervin in procesele de transformare a hranei (iaurt, vin); sunt saprofite, parazite, constituie flora bacteriana intestinala (in tractusul digestiv al omului sunt peste 10.000 de miliarde de bacterii, adica de cca. 10 ori mai mult decat toate celulele corpului la un loc).
B. Regnul Protista (Eucariote unicelulare)
Cuprinde:Ciuperci (drojdii, levuri), alge unicelulare (alge verzi, rosii, brune), Diatomee, Euglenophycee, Dynophicee, Protozoare.
Protoazoarele vor fi analizate pe larg in capitolul urmator, ca "punct de plecare" in Zoologia nevertebratelor. Aici vom prezenta unele caracteristici ale celorlalte grupe. Asa cum se observa, eucariotele unicelulare se constituie intr-un ansamblu heterogen care cuprinde la un loc reprezentanti ai regnului vegetal (algele), animal (protozoarele) si fungi (ciuperci inferioare). Algele si ciupercile prezinta si forme pluricelulare. Asadar, in regnul Protista apare pentru I-a data intrebarea: planta sau animal ?.
- Amoeba (amiba)este un protist ce prezinta toate caracteristicile regnului animal: celula limitata de membrana deformabila, mobilitate prin pseudopode, sursa organica de energie, prada ingerta dupa inglobare in vacuole digestive.
- Diatomeele sunt eucariote unicelulare ce ilustreaza regnul vegetal (membrana limitata la exterior de un perete rigid; sursa initiala de energie este lumina existand plastide care capteaza aceasta sursa; plastidele sunt inconjurate de vacuole; nucleul are pozitie centrala);
- Euglena gracilis, E. viridis - sunt specii aparte prin comportamentul lor: la lumina se comporta ca un vegetal clorofilian autotrof; la intuneric se comporta ca un animal heterotrof; daca organismul este readus la lumina de la intuneric, el isi va relua comportamentul de autotrof (prin interventia cu raze UV sau antiibiotice asupra posturii de autotrof, acesta isi va pierde complet capacitatea de sinteza si va deveni definitiv heterotrof
- Levuri (ciuperci unicelulare, drojdii) (ascomicete, bazidiomicete, fungi imperfecte).Sunt un ansamblu de talifite (ciuperci si vegetale) cu un aparat vegetativ care se reduce la o singura celula, a carei forma variaza in functie de specie. Ne avand clorofila, levurile nu pot utiliza lumina ca surs`a de energie si nici gazul carbonic. Ele oxideaza intotdeauna un substrat organic. Depind de organisme moarte sau parti din organisme vii. Au reproducere asexuata si sexuata (in conditii favorabile). Sunt cunoscute sub denumirea de agenti de fermentatie alcoolica.
- Alge unicelulare (rosii, verzi, brune).Rodofitele unicelulare = Rhodophycea (alge rosii). La acestea se cunoaste numai reproducerea prin diviziune simpla. Unele sunt sub forma de celule independente dar in unele cazuri se agrega in formatiuni de tip cordon, celulele fiind inglobate intr-o masa gelatinoasa. Chlorophyceele (algele verzi), traiesc in ape dulci iar celula vegetativa, de culoare verde se deplaseaza cu ajutorul a 2 flageli. In conditii nefavorabile flagelii dispar, se sintetizeaza pigmenti carotenoizi si se coloreaza in rosu. Un ordin cuprinde celule imobile, fara flageli, grupate in colonii (celulele reproducatoare au insa flageli). Mai exista un alt grup de specii solitare sau grupate (cenobii), terestre sau de apa dulce, la care nu exista reproducere prin diviziune simpla. Ele nu se divid decat pentru a da spori (reproducere asexuata) sau gameti flagelati ori imobili (reproducere sexuata). Chromophyta (alge brune) cuprinde alge unicelulare si un grup de alge pluricelulare. Poseda o mare diversitate de forme, morfologie si mod de viata (prin natura pigmentilor, a rezervelor, implantarea flagelului si organizarea plastidelor se diferentiaza de algele brune propriuzise - chromofite pluricelulare).
- Euglenophyceele au o pozitie intermediara intre Chromophytha (alge brune) si Protozoa. In acest caz, structura unei specii de Chromophyta are caracteristicile unui vegetal dar si particularitati de protozoar. Nu au amidon ci paramilon. Acest grup prezinta toate gradele de nutritie, de la autotrofia perfecta la heterotrofie.
Dupa caracterele citologice unele specii apartin la protoazoarele flagelate (ex. kinetoplastidele); - exista forme care practica fagocitoza, altele sunt pradatoare, coprofage sau parazite; - nu se cunoaste o reproducere sexuata iar inmultirea se face numai prin mitoza particulara.
- Dynophyceae (alge pyrrhophyte). Adesea sunt incluse in Chromophyta. Poseda trihociste (organite devaginabile sub forma de filamente rigide).
De la alge la protozoare. Protiste cu incadrare nesigura. Sunt animale unicelulare cu incadrare nesigura in dicotomia autotrof - heterotrof. Unele se comporta ca si reprezentanti neclorofilieni (dinoficee, euglenoficee) sau protoazoare ce au caracteristici comune cu ciupercile (micetozoare = mixomicete). Asa cum am prezentat deja, Euglenoficeele sunt un grup de legatura intre cele 2 regnuri.
In ceea ce priveste mixomicetele inca se pune intrebarea - protozoare sau ciuperci ?.. In zoologia nevertebratelor sunt incluse printre amoebe (Protozoa; Rhizopoda), dar prin faptul ca formeaza un peduncul care in varf are un corp de fructificare ce produce spori, sunt considerate ciuperci. Cercetari de data recenta au elucidat multe aspecte interesante ale acestui grup.
Experiente efectuate pe specia Dyctiostelium discoideum (Bozzone, 2001) considera acest organism ca o "celula cu personalitate" sau o "amoeba sociala", acest comportament fiind deosebit de interesant si reglat de mediatori chimici. Acest organism este folosit actualmente pentru cercetari morfogenetice, de diferentiere celulara, comunicare intercelulara si comportament. Ciclul sa de viata fiind deosebit de interesan si consideram ca merita sintetizat aici (Fig. 32).
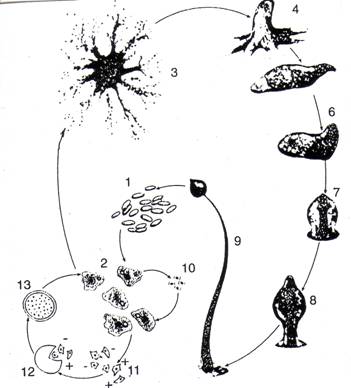
Fig. 32. Ciclul de viata la Dictyostelium discoideum (Protozoa - Rhizopoda). Detalii si semnificatia cifrelor, sunt date in text (dupa Bozzone, 2001).
Cand exista hrana suficienta (ex. bacterii ca Escherichia coli) indivizii se hranesc si comporta ca o amoeba (2 - mixamoeba; stadiu de hranire). In caz de saracire sau absenta a hranei se declanseaza comportamentul social constituindu-se o structura multicelulara (mii de celule) caracterizata de schimbari morfologice, biochimice sau moleculare (3 - agregarea). In continuare se constituie o formatiune denumita pseudoplasmodiu (4) care poate lua forma unei amoebe uriase, ce se poate deplasa dintr-un loc in altul (5) (de retinut ca la unele specii exista chiar plasmodiu adevarat). Treptat in interior se diferentiaza 2 tipuri de celule (celulele externe ale peretelui si sporii), organismul se fixeaza (6), incepe sa se formeze soroforul (sporofor) (7), de pe acesta creste un sorogen (8), iar in final ia nastere sorocarpul (9) care va elibera sporii ce vor da nastere la noi amoebe. Din acest moment dezvoltarea ontogenetica poate lua mai multe directii in functie de calitatea mediului de viata. In anumite conditii se formeaza microcisti (10) care dau nastere din nou la amoebe sau se diferentiaza doua tipuri de celule (+ si -; deci cu caracter de "mascul" si "femela" care vor constitui un organism diploid (12) definind un macrocist (13) din care de asemenea vor lua nastere amobe.
3. Subregnul Metazoare. Caracterizare generala a organismelor pluricelulare. Originea pluricelularitatii. Trecerea de la unicelulare la pluricelulare
3.1. Caracterizarea generala a metazoarelor. Diferenta dintre unicelulare si pluricelulare
Animalele studiate pana acum au avut o structura unicelulare. Sa ne ramintim clasificarea organismelor dupa Wittaker. Exista 5 regnuri: Monera (Procariota), Protista (Eucariota unicelulare), Plantae, Fungi si Animalia (ultimile 3 fiind Eucariota pluricelulare). Primele doua cuprind animale unicelulare; ultimile 3 cuprind animale pluricelulare. Grupul animalelor unicelulare constituie subregnul Protozoare iar cel al animalelor pluricelulare - subregnul Metazoare (vegetalele clorofiliene pluricelulare alcatuiesc Metafitele).
Unicelularele s-au caracterizat prin: material genetic neinclus in nucleu, enzime localizate in peretele celular (nu au organite celulare specializate extrem; cand exista ele de obicei sunt in forme derivate); reproducerea prin excelenta este realizata prin diviziune asexuata (sciziparitate (in cadrul protopzoarelor studiate am intilnit mai multe modalitati reproductive, frecvent cu alternanta de cicluri, mai ales datorita conditiilor de viata specifice impuse speciilor parazite). Am studiat si unele forme coloniale interesante, dar ele sunt probabil punti de trecere in evolutie spre speciile pluricelulare (sa ne reamintim alga albastra Rivularia, flagelatul Volvox). La unicelulare toate functiile vitale se realizeaza in interiorul unei singure celule printr-o specializare a zonei citoplasmatice in zone ce constituie "organite".
Pluricelularele au aparut ca o necesitate evolutiva. S-a realizat astfel "o diviziune a muncii" datorita tocmai unei diferentieri celulare. Diferentierea se face morfologic dar si functional celulele asociindu-se in tesuturi, organe, sisteme, aparate, organism. Studiul pluricelularelor mai implica si delimitarea dintre germen (celulele sexuale) si soma (toate celelalte celule ale organismului). La organismele pluricelulare materialul genetic este inclus in nucleu; au organite celulare distincte; reproducerea este dominant prin mitoza (indirecta, sexuata) dar la formele inferioare vom mai intalni si alte modalitati.
Fiecare animal pluricelular porneste practic in dezvoltarea sa de la o singura celula - celula ou. Aceasta va da prin diviziuni succesive un organism armonios. Specialistii afirma astfel ca celula ou este totipotenta (ea include in materialul sau genetic toate posibilitatile de exprimare ale tuturor celulelor specializate ale organismului). Pornind de la celula ou, totipotenta se va pierde in avantajul specializarii. Celulele vor primi astfel si forma si capacitatea corespunzatoare noilor lor functii (contractie musculara, vedere, transmiterea impulsului nervos, absorbtia substantelor, eliminarea toxinelor etc). Fata de dimensiunile protoazoarelor metazoarele sunt animale in general mari (cca. 30 m) dar este adevarat ca sunt si metazoare foarte mici, microscopice (infuzori, rotifere, acarieni, nematode). Numarul de celule care alcatuiesc corpul unui metazoar este diferit, variind dupa grupe si specii, de la cateva celule (ortonectide) pana la miliarde (de ex. corpul uman are 1014 celule; numai sangele are 259 globule).
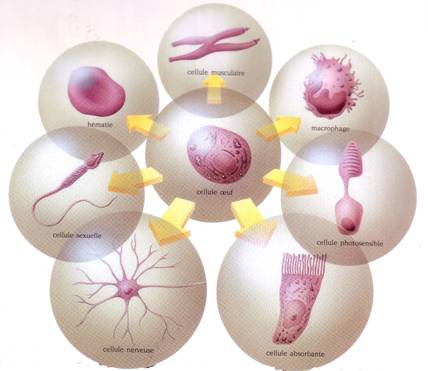
Fig. 33. Cateva tipuri de celule specializate la pluricelulare, datorita totipotentei celulei ou (dupa Encyclop. Larousse, 1993).
3.2. Originea animalelor. Originea pluricelularitatii (originea metazoarelor)
O chestiune fundamentala care se pune este acea daca animalele sunt monofiletice (deriva dintr-un singur stramos), difiletice (au doi stramosi) sau polifiletice ( au mai multi stramosi). Punctul de vedere polifiletic este atractiv pentru zoologi.
Aparitia aproape simultana a tuturor animalelor in fosilele din precambrian sau cambrian (cca. 550 milioane de ani in trecut) face dificila o explicatie monofiletica. Daca aceasta aparitie este reala si corecta, atunci exista mai multe explicatii asupra originii pluricelularitatii si mai multe forme de organisme ancestrale la origine.
Invers, similaritratile evidente ale structurii celulare ar inclina balanta spre punctul de vedere conform caruia ar fi existat un singur stramos. De exemplu, anumite jonctiuni celulare sunt similare la toate tipurile de celule animale, proteinele care acompaniaza miscarea sunt de asemenea similare, asterul se formeaza in timpul mitozei la marea majoritate a animalelor. Numai aceste aspecte si ar fi dificil sa se explice originea pe cale polifiletica. Deci ar fi o origine monofiletica ?.
Originea pluricelularitatii este inca inconjurata de mister. Unii autori chiar au format un grup ntermediar intre protozoare si metazoare - Mezozoare. Acesta considerandu-l ca o forma de trecere (aici au incadrat unele metazoare inferioare: Ortonectidele, Diciemidele; altii cred ca acestea sunt doar forme degradate de metazoare, datorita vietii parazitare pe care o duc).
Pana acum ne-am referit deci la organisme care pe tot parcursul vietii lor sunt unice (unicelulare) procurandusi independent materia si energia din mediul inconjuator, utilizandu-le pentru crestere si dezvoltare. Pluricelularele sunt "agregate de celule" dar cresterea si stabilitatea unei celule individuale esate dependenta de asocierea functionala stransa cu celelalte celule (sensul de "agregat" nu trebuie deci inteles ca o gramada de unicelulare adunate la un loc).
Daca ne referim la structuri ce sunt intermediare, intre nivelul de organizare unicelular si cel pluricelular, in urmarirea acestor aspecte este necesara o anumita pracautiune: cu cat analiza parcurge nivele tot mai inalte de complexitate biologica, se atribuie evolutiei, in mod nejustificat, complexitatea crescanda a organismelor actuale. Este normal ca progresul s-a realizat in cursul evolutiei dar nu si in mod necesar pe aceleasi cai.
3.3. Cazuri si forme interesante de trecere de la unicelulare la pluricelulare
Mai intai sa urmarim cateva organisme interesante care par a functiona la limita inferioara a pluricelularitatii. Vom analiza cate cazuri unde tendinta de trecere spre pluricelularitate este marcanta.
a. Cazul Rivularia (cyanobacteriilor; alge albastre Desi aparte, ca inferioritate pe scara evolutiei amintim totusi si acest grup caracterizat prin forma curioasa de organizare (in arborii filogenetici grupul este chiar prezentat separat de alte procariote (vezi Fig. 3). Desi cele mai simple unicelulare, considerate foarte mult timp in regnul Plantae, ele prezinta toate caracteristicile organismelor procariote. Ele au o fotosinteza ca a vegetalelor dar cea mai interesanta este gruparea celulelor in formatiuni care constituie alge filamentoas (trichome). O asemenea specie a grupului (Rivularia) este un organism pluricelular (?) si unde apare pentru I-a data o specializare a celulelor in: heterociste (celule mari, incapabile de fotosinteza, putin pigmentate, specializate in reducerea azotului atmosferic), akinete (celule voluminoase, alungite foarte pigmentate, bogate in rezerve = celule de supravietuire) si necridii sau disjunctorii (celule care se gelifica si impart colonia in segmente mai mici = hormogonii; aceasta este o a doua modalitate de diviziune, alaturi de diviziunea binara).
b. Cazul Pandorina morum. La aceasta specie (Flagelate; Phytomastigophora) indivizii care compun colonia sunt de acelasi fel, inglobati intr-o masa gelatinoasa si nediferentiati unii fata de altii (Fig. 34, A). Am vazut ca la Rivularia, desi mult inferioara, specializarea este marcanta
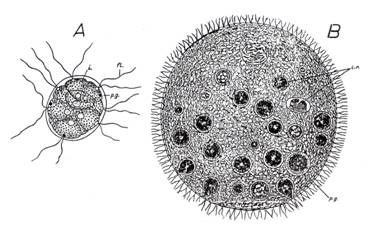
Fig. 34. Pandorina morum (colonie de flagelate fitomonadine)(A) si Volvox globator (un organism colonial) (B), care fundamenteaza ipoteza coloniala a pluricelularitatii. fl. = flagel; i. = individ al coloniei; i.v. = indivizi vegetativi; i.r. = indivizi reproducatori sexuati (micro- si macrogameti); p.g. = patura gelatinoasa (dupa Radu si Radu, 1972).
c. Cazul Volvox globator (Volvocale Este un grad crescut de colonizare si un inceput de diferentiere morfofiziologica. Am studiat deja pe Volvox globator (Fig. 34, B). Celula individuala este tip alga biflagelata, cu clorofila. Intr-o sfera (0,2 - 1,2 mm) cu cca. 10.000 celule (dupa unii doar 2.000 celule), acestea sunt dispuse intr-un singur sir la exterior, inglobate intr-o masa gelatinoasa, iar interiorul este gol. Unitatea sub care se desfasoara activitatea acestui organism poate fi atribuita filamentului protoplasmatic care leaga celulele componente. Initial Volvox a fost considerat o colonie, dar unii zoologi prefera sa-l considere multicelula (analogie marcanta cu notiunea de organism). Volvox actioneaza ca un tot unitar, nu se deplaseaza la intamplare, are o parte anterioara si una posterioara (deci are "cap" si "coada"). Cavitatea acestei sfere formata dintr-un singur strat de celule poarta denumirea de celoblastula. Daca o celula sau un grup de celule (agregatele-fiice) sunt izolate din agregatul pluricelulare si sunt puse in conditii similare, ele nu regenereaza un alt agregat
d. Cazul Dictyostelium discoideum (Myxomiceta) (vezi Fig. 32). O parte din viata aceasta specie exista ca celule amiboidale individuale. Fiecare dintre ele ia nastere dintr-un spor. Cultivate in laborator pe medii speciale, aceste celule traiesc independent, exact ca majoritatea altor organisme unicelulare, ele se hranesc, cresc si se reproduc. Dar, cand sunt conditii neprielnice mai multe asemenea amibe se aglomereaza in jurul unui centru de agregare, se aseaza una peste alta si formeaza o masa gelatinoasa alungita (ca un limax in miniatura !). Aspectul cel mai interesant acum apare: aceasta masa actioneaza ca un tot unitar, ea se deplaseaza intotdeauna cu aceeasi parte inainte si adeseori ca reactie de raspuns la un anumit factor extern (temperatura, lumina). Deplasare aceasta reaminteste de cea a unui melc. La un moment dat masa gelatinoasa se opreste, se fixeaza, incep o serie de miscari morfogenetice precise, prin care masa de celule initiale se ridica pe o tija verticala. Celulele acestea din varf se transforma in spori, care ajunsi intr-un mediu prielnic vor da nastere unei noi amibe. Este evident ca in momentul formarii structurii de fructificare are loc o diferentiere pronuntata in structura si comportament.
3.4. . Ipoteze ale pluricelularitatii
Aparitia pluricelularelor are la baza cateva scenarii plauzibile. Vom incepe cu cele doua ipoteze moderne privind pluricelularitate:
a. Ipoteza coloniala. (Fig. 35, a). Dintr-un protist ancestral care s-a tot divizat, indivizii au ramas impreuna, apoi celulele s-au dispus intr-o anumita forma (vezi Volvox). Invaginarea a determinat formarea celui de al doilea rand de celule, formatiunea caracterizand tipul ancestral cu simetrie radiala. Plecand de aici s-au constituit speciile cu simetrie radiala dar posibil si cele cu simetri bilaterala (vezi in continuare pct. 3.7.).
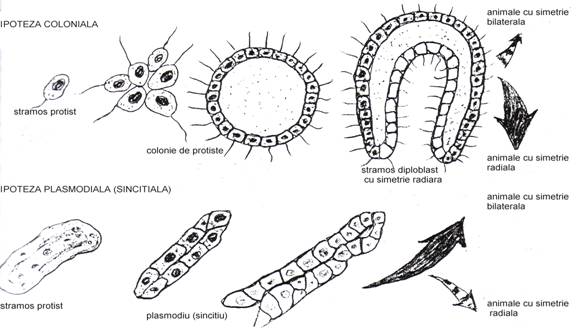
Fig. 3 Cele doua ipoteze privind originea pluricelularitatii. a - ipoteza coloniala; b - ipoteza sincitriala (plasmodiala) (dupa Miller si Harley, 1992).
b. Ipoteza sincitiala (plasmodiala (Fig. 35, b). S-a pornit de la un protist sincitial unde nucleul s-a divizat de multe ori dar citoplasma nu l-a partajat. Astfel, in aceeasi citoplasma erau mai multi nuclei. In etapa urmatoare citoplasma inconjoara nucleii si apar membranele care delimiteaza celulele nou formate. Se delimiteaza mai intai celulele de la periferie care delimiteaza ansamblul. Formatiunea evolueaza spre tipul de individ ancestral cu simetrie bilaterala de unde se trag animalele cu simetrie bilaterala.
In tratate mai vechi, ipoteza coloniala era denumita ipoteza volvocala. Sa mai amintim insa si alte ipoteze care au existat si au fost mai mult sau mai putin combatute. Iata 3 asemenea ipoteze, dar la care vom constata ca in realitate ca punct de plecare toate au la origine o ipoteza coloniala. Astfel sunt:
a. Ipoteza regruparii Organisme unicelulare se grupeaza in mase mai mici sau mai mari si se constituie in regiuni bine definite pentru diferite functii (ex. Mixomicetele
b. Ipoteza gastreei A fost o ipoteza interesanta, elaborata de Haeckel (sf. sec. XIX). Teoria a avut la baza cercetarile embriologice ale lui Kovalewski. Conform teoriei acestuia pentru toate metazoarele s-a generalizat existenta fazelor primitive ale dezvoltarii ontogenetice: morula, blastula, gastrula. Conform principiului sau ca ontogenia repeta filogenia, Haeckel a presupus ca fazele de blastula si gastrula ale dezvoltarii embrionare la metazoare ar aminti de constitutia stramosilor acestui grup. Adica, intr-o anumita etapa a dezvoltarii istorice, stramosii trebuie sa fi avut si o structura simpla, dintr-un singur strat de celule (asemanator cu o blastula). Astfel, el a dat acestor stramosi ipotetici denumirea de blastea. Intr-o etapa urmatoare stramosii au avut si o structura didermica (realizata prin invaginare ca si la celogastrula) si deci ia numit gastreea.
Hesse si Dofflein (1935) au imbinat aceasta ipoteza cu cea volvocala (coloniala) si au sugerat modul in care s-a trecut de la blastea (celoblastula Volvox) la stadiul didermic de gastreea (animalul s-a deplasat mereu in aceeasi directie, polul anterior s-a specializat pentru miscare, iar polul posterior pentru nutritie (fagocitare); polul posterior a fost cel care s-a invaginat fiind creat astfel la interior un spatiu mai favorabil pentru fagocitare = cavitatea gastrala).
c. Ipoteza parenchimelei Mecinikov combate teoria lui Haeckel, pe motiv ca la cele mai inferioare metazoare larva caracteristica nu este o celogastrula ci o parenchimula (la spongieri, celenterate). (Acum stim cum se formeaza aceasta - vezi Fig. 36, A)
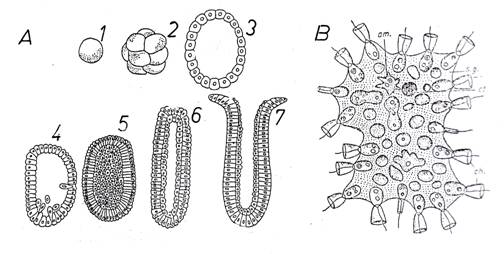
Fig. 36. A. Parenchimula la metazoarele inferioare (model schematic prin ilustrarea ontogenezei hidrei in cadrul inmultirii sexuate); 1 = ou; 2 = morula; 3 = blastula cu blastocel; 4 - imigratia celulelor; 5 = parenchimula; 6 = resorbtie interioara cu aranjarea celulelor interioare pe un singur strat = formarea planulei; 7 = deschiderea cavitatii. B. Proterospongia haeckeli. Am. = individ amiboid; ch. = individ flagelat cu guler (choanocit); ct. = chist; s.g. = substanta gelatinoasa
d. Ipoteza palintomiei. A fost elaborata de zoologul Zahvatkin pornind de la cercetari efectuate asupra embriologiei nevertebratelor inferioare. Conform principiului sau ca: nu exista metazoare care sa ramana pluricelulare tot timpul ciclului vital, ci ontogenetic, starea pluricelulara succede intotdeauna starea unicelulara (oul). Oul este o celula hipertrofica iar segmentarea sa este de tip palintomic. De fapt ce este palintomia ?. El considera acest proces ca un mod de diviziune al unor protozoare sau celule. Diviziunea binara obisnuita este una de tip monotomic (adica cresterea alterneaza regulat cu diviziunea; indivizii care au luat nastere cresc pana isi dubleaza dimensiunea si apoi iar se divid
In palintomie exista o crestere hpertrofica, adica sa depaseste cu mult dublarea volumului initial, iar in momentul in care se declanseaza diviziunea, aceasta se repeta de mai multe ori, fara repaus si deci fara a da timp indivizilor sa mai creasca Astfel ar lua nastere in mod rapid un numar de 2, 4, 8, 16, 32 indivizi sau celule din ce in ce mai mici si procesul se opreste cand se ajunge la marimea de la care s-a pornit. Unii indivizi se pot separa unii de altii dar cei mai multi se constituie in forme coloniale
Conform afirmatiei ca metazoarele sunt pluricelulare sa ne reamintim ca la protoazoare am intalnit unele forme constituite din mai multe celule (la coanoflagelate, radiolari, ciliofore). Toate aceste forme sunt insa colonii. Asadar, in colonii fiecare unitatea asociata isi pastreaza individualitatea si chiar poate trai izolata daca este separata de colonie. In colonie nu exista organe sau organite comune, deci o coordonare de tip superior a unor functii dintre indivizii care compun colonia. Unele formatiuni pluricelulare de la protozoare (ex. pansporoblasii, sporii de la microsporidii) sunt doar stadii trecatoare, sau forme de rezistenta in ciclul evolutiv al protoazoarelor.
La metazoare putem vorbi de organizare, de un sistem, caracterizat de structura si functie (structura arata legatura dintre elementele componente, iar functia indica faptul ca orice subunitate isi subordobeaza functia intregului.
Conform conceptiei totipotentei celulei ou, integrarea si diferentierea fiziologica a partilor in unitatea organismului se face asadar in acelasi timp cu diferentierea morfologica. Toate celulele de acelasi fel se asociaza pentru a indeplini o aceeasi functie. Acestea sunt tesuturile (= o formatiune structurala alcatuita dintr-un grup de celule caracterizate de aceeasi diferentire morfologica, aceeasi structura, aceeasi origine si s-au specializat pentru aceeasi functie) (ex. conjunctiv, epitelial, muscular, nervos, lichid circulant). Fiecare tesut este alcatuit din celule si substanta intercelulara.
Gruparile de celule si tesuturi, diferentiate de asemenea pentru diferite functiuni in cadrul organismului alcatuiesc organe. Fiecare organ nu va functiona insa izolat in interiorul organismului ci intr-o stransa relatie cu celelalte. In organe tesuturile sunt dispuse intr-o anumita ordine conform cu necesitatile functionale (in orice organ, un anumit tesut are o functie predominanta) (de ex. intr-un organ muscular va predomina tesutul muscular).
Organele sunt la randul lor grupate (asociate) in sisteme si aparate.
Sistemul este o unitate morfologica si functionala alcatuit din organe de aceeasi structura, deci formate din acelasi tesut (ex. sistemul osos, sistemul muscular, sistemul articular).
Aparatele sunt grupari de organe cu functie principla comuna dar structura poate fi diferita din punct de vedere morfologic Sunt unitati functionale ale corpului (ex. aparatul locomotor este constituit din oase, muschi si articulatii; are functia de sustinere si locomotie)( trebuie retinut ca ordine de ierarhizare ca aparatele sunt mai cuprinzatoare decat sistemele; vorbim de aparatul uro-genital, alcatuit din sistemul urinar si sistemul genital).
Totalitatea aparatelor si sistemelor constituie corpul (= un tot unitar morfologic si functional, in stransa corelatie cu mediul inconjurator Legatura functionala dintre partile corpului se realizeza pe doua cai: umoral (prin sange, limfa, lichid interstitial) si nervos.
Deosebirea importanta dintre protozoare si metazoare o gaim la nivelul reproducerii. La protoazoare am vazut ca procesele cele mai comune sunt cele asexuate (diviziunea binara, multipla, inmugurirea). Chiar si unde reproducerea sexuata apare ca o necesitate (gamogonia) ea este doar o faza intercalara a ciclului, niciodata predominanta, si cu atat mai mult exclusiva.
Mai intai sa ilustram comparativ modul de diviziune celulara: directa (amitoza) si indirecta (mitoza, cariokineza, citodiereza).
Cu exceptia celulelor nervoase toate celulele organismului se inmultesc. Formarea de celule noi din cele preexistente poarta numele de diviziune.
a. Diviziunea directa (amitotica) este caracteristica animalelor inferioare. Impartirea citoplasmei in doua este in majoritate urmata de diviziunea nucleului.
b. Diviziunea indirecta (mitotica) este procedeul cel mai frecvent al metazoarelor. Ea este de doua feluri: mitoza si meioza.
☻ - mitoza = diviziune celulara asexuata = reproducere clonala. Dupa o faza de crestere si activitate metabolica intensa (interfaza), in care compusii celulari se multiplica in doua, urmeaza diviziunea celulei mama in doua celule fiice cu un partaj egal de informatie genetica (Fig. 37). Asadar, procesul este o alternanta crestere - diviziune se formeaza o populatie de celule asemanatoare, de unde si numele de clona. Se mai numeste si diviziune cariocinetica deoarece se termina cu diviziunea nucleului si apoi a citoplasmei. Mitoza cuprinde o succesiune de 5 faze caracteristice: interfaza, profaza, metafaza, anafaza si telofaza.
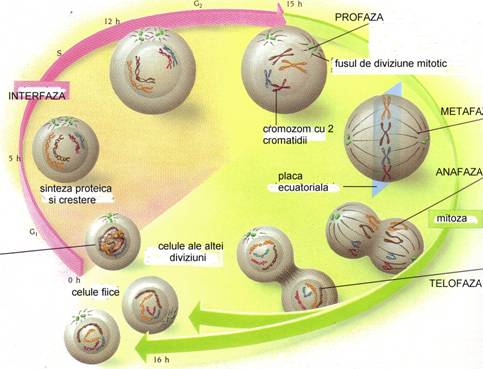
Fig. 37. Mitoza = diviziune celulara asexuata = reproducere clonala (dupa Encyclop. Larousse, 1999).
INTERFAZA. Este faza pregatitoare. Acum au loc 3 faze distincte: o faza de crestere (G1), o faza de replicare a AND (S) si o noua faza de crestere (G2). Dupa aceasta urmeaza mitoza propriu-zisa (M) (Fig. 37).
PROFAZA. Centrul celular se divide in 2 centrozomi (unul ramane la locul initial iar celalalt se duce spre suprafata nucleului si se aseaza la polul opus al celulei); acestia raman lgati intre ei printr-o formatiune denumita centrodesmoza; filamentele centrodesmozei se indeparteaza in zona de mijloc si alcatuiesc fusul de diviziune (fusul acromatic); nucleolii din nucleu se resorb si apar niste granulatii care constituie filamentul nuclear (contine linina si cromatina); la inceput filamentul nuclear este dispus ca un ghem = spirem dar apoi se fragmenteaza in fragmente = cromozomi; se resoarbe membrana nucleara;
METAFAZA. Prin diviziune fiecare cromozom da nastere lka 2 cromozomi fii; perechile de cromozomi se dispun in zona ecuatoriala a fusului acromatic; cromozomii se curbeaza si iau o forma caracteristica;
ANAFAZA. Cromozomii perechi se desprind si se deplaseaza de-a lungul fiecarui filament al fusului spre cei 2 centrozomi;
TELOFAZA. Ajunsi la polul celulei cromozomii se reunesc si refac filamentul nuclear care se transforma in spirem; sucul nuclear se dispune in jurul fiecarui spirem; se reface membrana nucleara; reapar nucleolii; filamentul nuclear dispare; regiunea ecuatoriala a celulelor apare o gatuitura la nivelul citoplasmei, se diferentiaza o membrana plasmatica care va separa celula in doua celula mama, fiecare avand un nucleu realizat prin diviziune .
☻ - meioza = reproducere sexuata (gametica) (Fig. 38). Este caracteristica celei mai mari parti a eucariotelor. Se numeste reproducere gametica = sexuata si insoteste modelul mitotic. Se deosebeste de mitoza prin faptul ca porneste de la o celula numita ou = zigot (celula diploida) care are un numar dublu de cromozomi si rezulta in urma unui proces denumit fecundare a doua celule care au numar redus la jumatate de cromozomi. Se poate spune deci ca meioza este o modalitate de formare de celule haploide denumite gameti. Fenomenul de formare a gametilor poarta denumirea de spermatogeneza si ovogeneza. Sunt procese complicate constand dintr-o succesiune de faze: replicare, recombinare, prima diviziune (reductionala), a doua diviziune (egala).
2 3 4 5 6 7 8 9
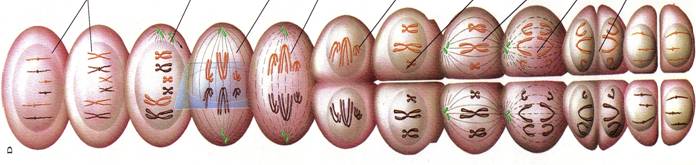
Fig. 38. Meioza = reproducerea sexuata la eucariote (dupa Encyclop. Larousse, 1999).
Succesiunea fazelor meiozei este mai complicata si cuprinde urmatoarele:
A. Gametii, gametogeneza. Ovulul si spermatozoidul sunt celule diferentiate ale organismului pluricelular si se constituie prin procese speciale caracteristice metazoarelor, atunci cand acestea ajung la maturitate sexuala. Procesele consta pe de o parte in reducerea substantei celulare si nucleare prin numitele diviziuni de maturatie. Se mai numesc si reductionale deoarece numarul cromozomilor se reduce la jumatate.
Ovogeneza (Fig. 39). Reprezinta formarea ovulelor. Celulele genitale femele cele mai putin diferentiate = ovogonii se divid de mai multe ori pana ce ajung in stadiul de ovocite (recunoscute prin faptul ca se incarca cu substante de rezerva si deci sunt si mai voluminoase). Se numesc si ovocite de gradul I. Acum urmeaza doua diviziuni de maturatie si din ambele rezulta celule inegale: una foarte mare si una mica (dupa prima diviziune de maturare celula mare este ovocit de gradul II; celula mica este globul polar; dupa cea de a doua diviziune reductionala celula mare este ovul iar celula mica repreznta al doilea globul polar) (Globulele polare sunt omoloage cu ovulele dar sunt degenerate si vor pieri).
Fig. 39. Reprezentarea schematica a gametogenezei la metazoare. Ovg. = ovogonii; Spg. = spermatogonii; Ovc.I = ovocit I; Ovc. II = ovocit de grad II; Spc.I = spermatocit de grad I; Spc.II = spermatocit de grad II; Sptd. = spermatidii; Sp. = spermatozoizi; gl.p. = globule polare. Punctele negre reprezinta stadii diploide (2n) adica cele cu numar dublu de cromozomi; punctele albe reprezinta stadii haploide (n), adica numarul de cromozomi este redus la jumatate
Spermiogeneza (spermatogeneza) (Fig. 39). Este formarea spermatozoizilor. Se porneste de la celule putin difeentiate = spermatogonii care, ca si ovogoniile, sufera mai multe diviziuni si se formeaza spermatocitele de gradul I (acestea nu cresc insa asa de mult ca ovocitele, nu acumuleaza substante de rezerva iar diviziunea este egala). Deci, in urma celor doua diviziuni de maturatie rezulta 4 celule egale = spermatidii, din care vor lua nastere spermatozoizii
Unirea spermatozoidului cu ovulul constituie procesul numit fecundare. Reproducerea prin participarea ambelor sexe, adica pornind de la ou fecundat se numeste amfigonie. Daca este o reproducere din oua fara fecundatie se numeste partenogeneza. Odata ce s-a format oul acesta se va dezvolta si va da nastere unui nou organism pluricelular, trecand prin diferite stadii care la un loc alcatuiesc ontogeneza.
Tipuri de ovule si de spermatozoizi la metazoare Ovulele sunt denumite in sens mai larg si "oua". Sunt gameti femeli imobili si au cantitati variabile de substante nutritive ceea ce face sa existe mai multe tipuri de oua: oligolecit [gr. oligos = putin; lekitos = vitelus], mixolecit [gr. mixis = amestec; lekitos = vitelus], centrolecit [gr. centros = mijloc; lekitos = vitelus], telolecit [gr. telos = capat; lekitos = vitelus] (Fig. 40)
Spermatozoizii sunt elemente mascule, in general mobile, de forme foarte variate. Pot fi amiboizi, cu prelungiri, flagelati (Fig. 40).
Din punct de vedere chimic, spermatozoizi si ovulele au proprietati diferite si specifice. Spermatozoizii prezinta un chimiotactism extrem de pronuntat fiind atrasi spre ovul datorita unor substante (mediatori chimici) denumite fertilizine (elaborate de ovul si eliminate la exterior. Ajunsi la membrana oului spermatozoizii secreta alte substante cu ajutorul carora patrund inauntru. Alti mediatori chimici vor interveni pentru a nu patrunde mai multi spermatozoizi, primul dintre cei mai dotati fiind singurul acceptat.
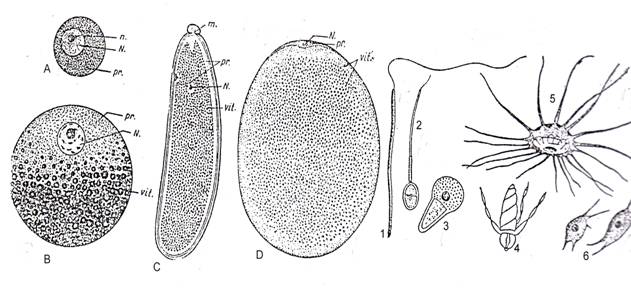
Fig. 40. Diferite feluri de oua (A-D) si diferite tipuri de spermatozoizi (1-6). Oua: A - ou oligolecit (alecit) de stea de mare; B - ou mixolecit de gastropode; C - ou centrolecit de diptere; D - ou telolecit de la cefalopode. m. = micropil; n. = nucleol; N. = nucleu; pr. = protoplasma; vit. = vitelus. Spermatozoizi : 1 - Bithynia sp (Mollusca); 2 - Homo sapiens (Hominidae) ; 3 - Ascaris sp. (Nematoda); 4 - Eupagurus sp. (Crustacea); 5 - Astacus sp. (Crustacea); 6 - Polyphemus pediculus (Crustacea inf.); (dupa Radu si Radu, 1972).
3.7. Ontogeneza. Stadii si etape
1. Segmentatia. Formarea blastomerelor si constituirea morulei (Fig. 41). Primele stadii ale ontogenezei sunt in general comune tuturor metazoarelor. Oul este supus unor diviziuni succesive care constituie procesul denumit segmentatie si se formeaza grupe de mai multe celule = blastomere (acestea nu se vor separa unele de altele, ca la diviziunea binara a protozoarelor, ci raman intr-o unitate formand un masiv de celule = morula (Fig. 41,A;c-g; B;b-c). Segmentatia prezinta diferite aspecte in functie de grupele de animale, dar pe de alta parte, acestea sunt constante asociat cu cantitatea de vitelus a oului iar pe de alta parte cu caracterele sale ereditare. Exista urmatoarele tipuri de segmentare:
a. Segmentarea totala Toate ouale care au putin vitelus (alecit; oligolecit) se divid complet deci au o segmentare totala (Fig. 41, A,B). Aceasta poate fi:
* - egala: (mai corect subegala); blastomerele sunt mai mult sau mai putin egale; cantitatea de vitelus este putina si repartizata uniform in ou;
* - inegala (in cazul oualor mixolecite) (Fig. 41, B). Blastomerele formate sunt in mod evident inegale: cele superioare sunt mici si sarace in vitelus; cele inferioare sunt mari si incarcate cu vitelus (Fig. 41, B).
De fapt se obtine o morula cu micromere (polul animal) si macromere (polul vegetativ).
Dupa cum se dispun blastomerele una fata de alta (atat in segmentarea egala cat si inegala) s disting 4 categorii de segmentatie:
☻- segmentatia radiala (ex. spongieri, celenterate, echinoderme);
☻- segmentatia spirala (ex. la viermi si la moluste);
☻ segmentatia bilaterala (ex. rotifere, nemertieni, tunicieri, vertebrate);
☻ segmentatia bisimetrica (singurul grup = ctenoforele).
b. Segmentarea partiala Toate ouale bogate in vitelus au o segmentare partiala (Fig. 41, C,D) (deoarece vitelusul nu poate fi inclus in blastomerele care se formeaza). Si acest tip este de 2 feluri:
☻ segmentatie discoidala Este caracteristica oualor telolecite (adica ouale la care vitelusul se stringe in cantitate mare la polul inferior al oului = polul vegetativ); ovoplasma este deasupra sa si pluteste ca un disc (discul germinativ) la polul superior al oului (polul animal)(Fig. 40; D).; se va segmenta numai acest disc = disc embrionar (Fig. 41;C); ulterior, marginile discului vor inconjura vitelusul si vor forma vezicula vitelina (sacul vitelin) (ex. la scorpioni si la majoritatea vertebratelor);
☻ segmentatie superficiala. Se intalneste numai la artropode, care au oua centrolecite (Fig. 40; C); se formeaza blastomere numai la periferia oului unde se afla ovoplasma ca un strat subtire; se formeaza astfel un blastoderm periferic care inconjoara vitelusul din toate partile.
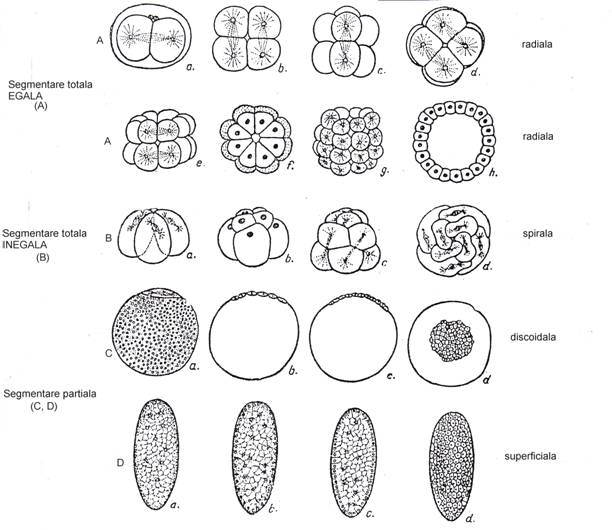
Fig. 41. Diferite tipuri de segmentare ale oului la metazoare. A = segmentarea totala si egala (la Synapia digitata si la ariciul de mare = Paracentrotus lividus -A;d,g.h -; B = segmentare totala si inegala; C = segmentare partiala discoidala (la scorpioni); D = segmentare partiala superficiala (la Hydrophilus piceus); A - segmentare radiala; B - segmentare spirala (la Trochus) (dupa Radu si Radu, 1972).
2. Formarea blastulei si gastrulatia Indiferent care este tipul segmentatiei, blastomerele continua a se inmulti si constituie o vezicula sferica cu peretele format dintr-un singur strat de celule, iar cavitatea interna este fara celule dar plina cu lichid. Acesta este stadiul de blastula, constituita deci dintr-un perete unistratificat de celule = blastoderm si o cavitate interioara = blastocel (cavitate primara; cavitate de segmentatie). Se mai distinge polul animal (cu blastomere mici) si polul vegetativ (cu blastomere mari). Acest tip de blastula = celoblastula. Nu intotdeauna situatia se prezinta in acest fel. In unele cazuri micromerele polului animal formeaza doar o micuta calota deasupra macromerelor fara cavitate de segmentatie iar formatiunea aceasta este o stereoblastula
Gastrulatia Dupa blastula urmeaza stadiul de gastrula. Este o formatiune mai complicata, alcatuita din 2 paturi celulare si care se formeaza prin procese mai simple sau mai complicate.
Se cunosc 3 mdalitati de formare a gastrulei: prin invaginare (embolie), prin epibolie (invaluire) si prin imigratie (Fig. 42).
☻ gastrulatia prin invaginatie embolie). Este cea mai comuna; se realizeaza printr-o invaginarea a blastodermului, aceasta se mareste si se adanceste treptatsi astfel se va forma o larva din doua foite celulare: una la exterior = ectoblastul (ectodermul) si una la interior = entoblastul (endoblast, endodermul). Endoblastul se constituie ca un tub, iar cavitatea astfel formata constituie arhenteronul (intestinul primitiv) si el comunica cu exteriorul printr-un orificiu = blastopor. Intre cele doua foite blastocelul este redus (uneori cele doua foite se alipesc si spatiul devine virtual); atunci cand exista constituie cavitatea primara a corpului (la metazoarele inferioare - spongieri - aceasta este plina cu un tesut nediferentiat care este mezoglea);
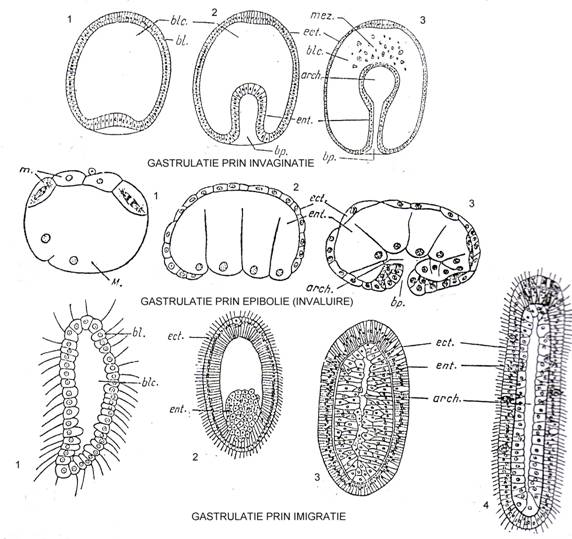
Fig. 42. Formarea blastocelului si gastrulatia. In 1-3 (4) secvente sunt prezentate cele 3 tipuri de gastrulatie: gastrulatie prin invaginatie; gastrulatie prin epibolie (invaluire); gastrulatie prin imigratie. bl. = blastoderm; blc. = blastocel; ect. = ectoblast (ectoderm); ent. = entoblast (endoderm); arch. = arhenteron; bp. = blastopor; mez. = mezenchim (dupa Radu si Radu, 1972).
☻ gastrulatia prin invaluire (epibolie). Aceasta pleaca de la o stereoblastula si are urmatoarele aspecte: blastomerele de la polul animal sunt mai mici decat cele de la polul vegetativ si fiind sarace in vitelus se inmultesc foarte rapid si reusesc sa inconjoare macromerele care sunt greoaie. Dupa ce le inconjoara complet, se constituie o larva tot cu 2 foite (ectoblast si endoblast), deci o gastrula dar care nu are arhenteron si este numita stereogastrula;
☻ gastrulati prin imigratie Daca in primele doua tipuri au existat "miscari celulare" in bloc, in acest tip de gastrulatie celulele se misca individual. Spre polul inferior, vegetativ al blastulei (care este o celoblastula) sau chiar pe toata intinderea blastodermului, din loc in loc se observa celule care ies din rand si se aglomereaza spre interior pana ce umplu blastocelul. Se formeaza astfel un endoblast masiv inconjurat de un ectoblast obisnuit. Un asemenea stadiu intalnit la spongieri si la celenterate constituie larva parenchimula. Ea inoata cu ajutorul cililor de la suprafata.. La celenterate (aceeasi figura), celulele endoblastului se vor organiza apoi intr-un singur strat reducand la o stare virtuala cavitatea primara. In mijloc apare insa o cavitate inconjurata de endoblast si care corespunde intestinului primitiv. Larva aceasta se numste planula
3. Formarea mezodermului La cele mai inferioare metazoare (spongierii) cele doua foite (ectoblastul si endoblastul) reprezinta singurele foite embrionare primordiale pe seama carora se vor forma organele definitive. Ele constituie grupul animalelor diploblaste (didermice). La celelalte metazoare, procesul de gastrulatie duce la formarea unui embrion care odata cu endoblastul formeaza si a 3-a foita numita mezoblast (mezoderm) (am amintit ca la diploblaste exista o structura nediferentiata care este denumit mezoglee).
Procesele prin care se formeaza mezodermul sunt variate si complicate. La nevertebrate doua dintre cele mai frecvente sunt: teloblastic si enterocelic. Schema generala a celor doua procedee este ilustrata in Fig. 43.
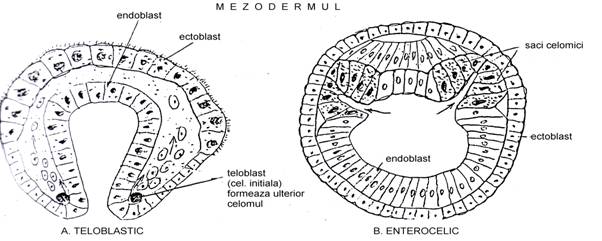
Fig. 43. Reprezentarea schematica a modelului teloblastic (A) si enterocelic (B) de formare a mezodermului
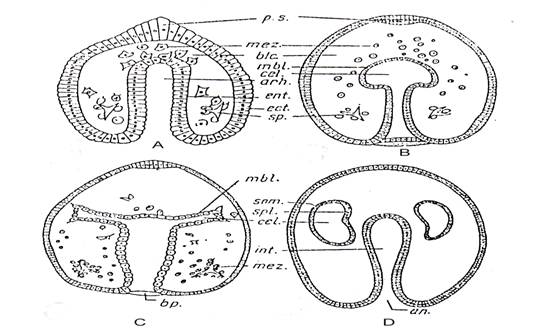
Fig. 44. Formarea mezodermului si mezenchimului la Echinodermata (Asterias sp.). A - gastrula; B - initierea celomului (prin formarea unei dilatatii la extremitatea arhenteronului); C - formarea a doua diverticule simetrice a dilatatiei initiale a arhenteronului; D - formarea a 2 saci celomici din cele doua diverticule. Aici arhenteronul a devenit astfel intestin iar blastoporul - anus; acestea sunt animale deuterostomiene. an. = anus; arh. = arhenteron; blc. = blastocel; bp. = blastopor; cel. = cavitate celomica; ect. = ectoblast; end. = endoderm; ent. = entoblast; int. = intestin; mbl. = mezoblast; mez. = mezenchim; p.s. = placa sincipitala; som. = somatopleura; sp. = spicul; spl. = splanchnopleura (dupa Radu si Radu, 1972).
tipul teloblastic Este un model de formare care porneste de la cateva celule initiale. Se intalneste la viermi, artropode si moluste. In timpul formarii gastrulei, in regiunea blastoporului, in blastocel, intre ectoblast si endoblast apar doua celule denumite teloblaste (initiale). Ele sunt simetrice bilateral. Acestea sunt celulele initiale ale mezoblastului.
Prin inmultire se vor forma niste benzi celulare mezodermice. In continuare, din aceste benzi, prin segmentare transversala si schizocelie, se formeaza vezicule mezodermice (vezicule celomice) (pe masura ce aceste vezicule cresc, ele ocupa locul cavitatii primare si aceasta devine virtuala (Fig. 43). Veziculele mezodermice sunt constituite dintr-un singur strat de celule; peretele dinspre ectoderm se numeste somatopleura; peretele lor dinspre intestin se numeste splanchnopleura; cele doua foite se ating dorsal si ventral si ele formeaza mezenterul dorsal si mezenterul ventral (care sustin intestinul). Cavitatea veziculelor constituie cavitatea celomica sau cavitatea generala (din cauza ca aceasta inlocuieste cavitatea primara a corpului este o cavitate secundara). Cavitatea secundara (ca de altfel si cavitatea primara) apare astfel complet inchisa; ea comunica cu exteriorul numai prin intermediul anumitor organe (ex. organele genitale; organele excretoare).
☻ tipul enterocelic. Se realizeaza prin evaginatii ale endoblastului (endodermului). Se intalneste la metazoarele cele mai superioare (echinoderme, chetognate, cordate; sub forme primitive se intalneste si la unii viermi).
4. Formarea mezenchimului. Jucand un rol important in formarea unor organe, mezenchimul este un tesut evidentiat in special la metazoarele superioare. Fata de foitele embrionare descrise, mezenchimul este constituit din celule imprastiate cu mari spatii intre ele (Fig. 44). El se formeaza din proliferarea unor celule din mezoblast si ectoblast, aceste celule intrand in cavitatea primara a corpului. Mezenchimul se formeaza si la metazoare inferioare, diploblastice unde nu se intalneste mezoblast si unde ia nastere din ectoblast.
Sinteza. In cele ce urmeaza sa incercam sa constituim in imagini succesive segmentatia, gastrulatia, formarea si evolutia mezodermului la un grup evoluat de nevertebrate - protocordatele (acraniatele) (Fig. 45). Aici face parte celebrul Amphyoxus sau altfel spus, veriga mult cautata dintre nevertebrate si vertebrate. Acest animal are caracteristic:
- un sistem nervos central sub forma de tub incomplet inchis (deci este un jgheab), situat in partea dorsala si realizat prin infundarea ectodermului; cavitatea din interuiorul sa se numeste neurocel;
- coarda dorsala (notocordul) se afla sub sistemul nervos;
- tubul digestiv separat de peretele corpului prin cavitatile celomice se afla sub coarda dorsala;
- musculatura corpului are dispozitie metamerica.
Ilustram in Fig. 45 ontogeneza speciei Amphyoxus.
* - din ou, prin segmentare inegala rezulta blastomere (cu micromere si macromere) si se constituie morula;
* - din morula se formeaza blastula iar blastocelul este ocupat de o substanta gelatinoasa
* - gastrulatia se face prin invaginare in mai multe etape: substanta gelatinoasa migreaza spre exterior; cele doua foite sunt practic alipite iar cavitatea primara este astfel virtuala;
* - urmeaza un proces de alungire a gastrulei si astfel se distinge un cap anterior si unul posterior;
* - se schiteaza jgheabul neural si coarda dorsala prin dezvoltarea mai mare a unor celule
* - din endoblast incep sa inmugureasca diverticole (sacii celomici mezodermici)
* - din partile dorso-laterale ale tubului digestiv iau nastere sacii celomici; coarda dorsala se accentueaza (deci ea are origine din tavanul intestinului ca si la hemicordate; in faze mai avansate se constituie tubul neural, coarda dorsala si sacul celomic divizat in epimer, mezomer si hipomer;
* - din fragmentarea cavitatii celomice se vor forma in stadiu mai avansat somitele, nefrotonul si celomul (foita dinspre intestin este splanchnopleura iar cea dinspre ectoblast este somatopleura).
3.8. Organogeneza
Oricare ar fi aspectele primelor procese ale ontogenezei, dupa foarmarea gastrului incep si procesele de formare a organelor. Aceste procese dureaza pana la maturitatea organismelor si imbraca procese variate. Astfel:
- din ectoblast deriva elementele sistemului nervos, organele de simt, parti ale aparatului digestiv, fibre musculare. Ceea ce ramane constituie invelisul corpului - ectodermul (care la randul lui poate produce diferite formatiuni) (Precizare: in mod corect cele doua foite ale gastrulei sunt entoblast si ectoblast; frecvent se uzeaza de terminologia de endoderm, respectiv ectoderm, desi am vazut ca ectodermul este invelisul corpului, acesta formandu-se din ectoblast
endoblastul dupa ce da nastere mezoblastului se numeste endoderm. El constituie epiteliul intestinului mijlociu cu diferitele sale glande digestive (la cordate, tot din endoblast ia nastere coarda dorsala, premergatoare coloanei vertebrale);
- din mezoblast deriva epiteliul celomic sau peritoneal, care limiteaza cavitatile celomice, cea mai mare parte a musculaturii corpului, organele de excretie, organele de reproducere si alte organe;
- din mezenchim iau nastere tesuturile de sustinere (conjunctiv, cartilaginos, osos), cromatofori, vase si globule de sange, musculatura organelor vegetative etc.
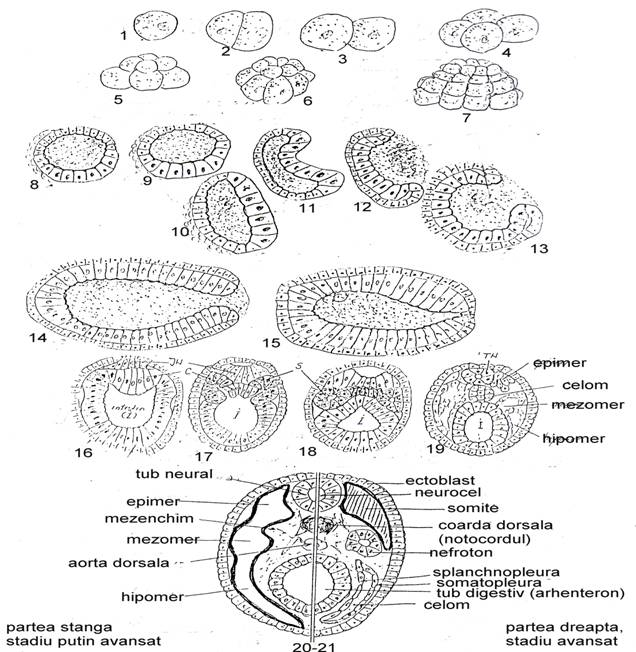
Fig. 4 Segmentatia, gastrulatia, formarea si evolutia mezodermului la protocordatele acraniate (Amphyoxus sp.) (detalii in text) (original).
4. Evolutia la nivelul regnului Animalia
Am aratat ca existenta celor doua foite embrionare (ectoblast si endoblast) definesc grupul animalelor diploblastice, iar aparitia si celei de a 3-a foite (mezoblastul), pe cel al triploblastelor. Pentru evolutia in continuare au fost sugerate doua scenarii (Fig. 46):
a. triploblastele provin dintr-un stramos diploblastic. Astfel dintr-un stramos de tip protista s-au desprins pe de o parte poriferele (spongierii), iar pe de alta parte a luat nastere un stramos cu simetrie radiala. Din acesta s-a tras un stramos diploblastic cu simetrie radiala (aici fac parte cnidarii, ctenoforele), iar pe de alta parte s-a constituit un stramos triploblastic acelomat. Din el au luat nastere: acelomatele, pseudocelomatele si eucelomatele (acestea din urma cu cele doua linii: protostomiene si deuterostomiene) (Fig. 46 - 1);
b. o alta ipoteza sugereaza ca formele triploblastice au avut o evolutie mai rapida si aceasta datorita faptului ca din stramosul protist a evoluat un stramos cu simetrie bilaterala din care s-a tras direct stramosul triploblastic acelomat. Din acesta s-au desprins triploblastele (dupa acelasi model ca in imaginea anterioar dar si diploblastele cu simetrie radiala Fig. 46 - 2).
Fig. 46. Originea si evolutia animalelor. 1 = ipoteza dupa care triploblastele au provenit din stramos diploblastic; 2 = ipoteza dupa care triploblastele au provenit dintr-un stramos cu simetri bilaterala imediat cxe au aparut pluricelularele iar diploblastele cu simetrie radiala se desprind tot din triploblate (dupa Miller si HARLEY, 1992)
Spongierii, Cnidarii si Ctenarii sunt formele cele mai primitive de Metazoare. Se caracterizeaza prin existenta numai a celor doua foite embrionare (ectoblast si endoblast: ectoderm sau endoderm), separate de mezoglee sau de mezenchim.
Porifera (Spongieri . Sunt cei mai putin evoluati. Nu au organe de simetrie, sistemul nervos este rudimentar, difuz, din celule putin diferentiate. Prezenta celulelor flagelate (la gulerul choanocitelor) sugereaza ideea existentei unui parinte comun intre Choanoflagelate si Spongieri (pentru acest motiv in arborele de evolutie animal au o pozitie izolata - Fig. 80). Acest grup asigura astfel trecerea de la Protozoare la Metazoarele adevarate. Aceasta interpretare este indoielnica deoarece multiple caractere ( modalitatile gamnetogenezi si fecundarii, tipurile structurale si functionale de celule: pinacocite, archeocite, choanocite) la face sa fie veritabile Metazoare.
Cnidaria. Sunt deja mai evoluate. Ele poseda o simetrie axiala; sistemul nervos formeaza mai mult sau mai putin o retea sau un plex si asigura coordonarea actibitatii gratie impulsurilor primite de la organe senzoriale diferentiate (tactile, chemoreceptoare, statoreceptoare, fotoreceptoare). Mezoglea lor este umpluta mai mult sau mai putin cu celule migratoare. Se diferentiaza niste benzi mici musculare. Exista un inceput de organe respiratorii, glande digestive si genitale. Cnidarii formeaza un ansamblu omogen caracterizat de aparitia unui tip particular de celule = cnidoblastele (elemente veninoase, urticante, cu multiple functiuni (aparare, captura, fixaere, constructie etc).
Acnidaria (Ctenophora . Plasati mult timp impreuna cu cnidarii se disting tatusi de acestia prin simnetria bilaterala, absenta cnidoblastelor, prezenta constanta a unor celule speciale cu proproetati adezive (coloblastele), localizate pe tentacule si cu rol in prinderea hranei. Mezoglea contine deja foarte multe celule putand fi considerata ca un mezenchim care ar fi debutul cele de a 3-a foite.
Asadar, au existat 3 stadii mari in ontogenia si filogenia animalelor: I = stadiul unicelular; II = stadiul diploblastic (constituit din segmentare si gastrulatie); III (stadiul triploblastic).
Este etapa ultima a avolutiei animalelor, caracterizat deci prin aparitia celei de a 3-a foite embrionare, mezodermul, care substituie mezogleea de la organismele diploblastice. Aceasta aparitie defineste deci animalul triploblastic care va atinge un grad inalt de complexitate: o simetrie bilaterala, o ierarhizare functionala, neurosenzoriala sau neuro-endocrina, definirea unei regiuni cefalice dominante. Destinatia celei de a 3-a foite va permite scindarea organismelor triploblastice in: acelomate, pseudocelomate si eucelomate (frecvent este folosita terminologia de acelomate si celomate).
Intr-o schema sintetica (Fig. 47) este ilustrata reprezentativ diferenta dintre cele 3 grupe dupa modul in care este dispusa si organizata a 3-a foita embrionara.
La acest grup (Fig. 47) mezodermul ramane compact, lipit de ectoderm si de endoderm; el nu se organizeaza niciodata in vezicule de tip inchis. Aceasta foita embrionara participa la constructia unui tesut difuz, parenchim, care umple cavitatea primara, fiind sediul unor organe noi: muschii parietali si viscerali, organe genitale. Ansamblul de organisme triploblastice primitive (acelomate) cuprinde: Plathelminte, Nemertieni, Nemathelminte, Rotifere iar dintre Moluste - Ortonectidele si Dicyemidele.
Platihelmintha. Sau viermii plati. Au un parenchim abundent. Tubul dgestiv este bine individualizat si cuprinde portiuni distincte care asigura o functionare pe masura. Intra in aceasta increngatura si forme libere de dimensiuni reduse (Turbelariate) care conserva arhitectura primitiva a Plathelmintelor, sau forme parazitare de talie mare (Cestode, Cestodari, Trematode).
Nemertea (Nemertieni) Dupa unii nu ar trebui sa fie incadrate la acelomate deoarece au un tip de celom aparte. Astfel, parenchimul este intotdeauna foarte dezvoltat, protonefridiile si organele genitale sunt independente si in mod regulat distribuite de o parte si de alta a unui plan de simetrie bilaterala.
P s e u d o c e l o m a t e l e
Nemathelminta. Sau viermi rotunzi. Mezodermul este redus la cateva celule provenite din tubul digestiv, organele genitale si excretoare si care se misca libere in lichidul perivisceral.. Increngatura comporta specii libere sau parazite.
Orthonectida si Dicyemida (grupa Mezozoare) deriva din Platihelminte parazite si in continuare adaptate la parazitis (la aceste organisme mezodermul a disparut).
*Rotifera, Nematorhyncha (Gastrotricha, Echinodera), plus clasele de Acanthocephala si Priapulida prezinta anumite afinitati cu alte Metazoare triploblastice acelomate.
In unele tratate, grupurile de mai sus sunt incadrate la acelomate. Dupa conceptiile mai noi, aici sunt cuprinse urmatoarele increngaturi: Loricifera, Gnathostomulida, Gastrotricha, Rotifera, Kinorhyncha, Nematoda (Nemathelminta), Nematomorpha, Acanthocephala, Priapulida, Entoprocta.
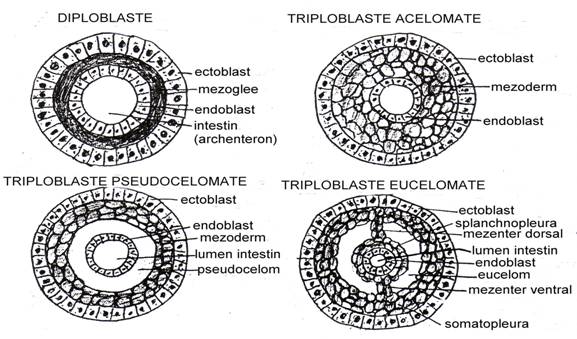
Fig. 47. Planul schematic de organizare al corpului la triploblaste. A = schema comparabila pentru diploblaste (ectoderm la exterior; endoderm spre intestin; intre el este doar mezoglee); B = schema pentru triploblaste; a = triploblaste acelomate (exista mezoderm dar acesta umple complet spatiul dintre ectderm si enfdoderm); b = pseudocelomate (mezodermul este lipit de ectoderm si exista un spatiu intre intestin si mezoderm = pseudocelom); c = eucelomate (sub forma de vezicule inchise, celomul este format normal, derivat din tesuturile mezodermale; prin cele doua mezentere ese sustinut intestinul invelit de splanchnopleura). int. = intestin (dupa Miller si Harley, 1992).
E u c e l o m a t e l e
Sunt organismne triploblastice cu celom adevarat. Celulele mezodermice constituie, de o parte si de alta a tubului digestiv, masive celulare perechii, simetrice care se organizeaza in cursul dezvoltarii embrionare in vezicule inchise = vezicule celomice. Ansamblul acestor vezicule, nascute enterocelic (Echinoderme) sau teloblastic = schizocelie (Anelide) reprezinta celomul adevarat (Fig. 82, 83). Aceasta fragmentare sau segmentare a mezodermului (mozomerie) este insotita de o distributie periodica a maselor musculare (miomerie), nervoase (neuromerie), formatiunilor excretoare (nefromerie) sau genitale. In acest fel, corpul este impartit in unitati anatomice si functionale = metamere. Aceasta metamerizare este insotita la randul ei de o condensare, o cefalizare a elementelor nervoase din partea anterioara, fenomen care atinge apogeul la Cephalopode, Insecte si Vertebrate.
Ansamblul organismelor triploblastice este pe de alta parte heterogen. Daca se urmareste numai destinul blastoporului atunci se disting doua linii evolutive naturale: protostomienii si deuterostomienii.
La aceasta prima linie evolutiva, blastoporul (care la diploblaste are dubla functie de gura si anus) constituie gura si nu da niciodata direct in anus; blastoporul poate chiar disparea complet, gura si anusul fiind atunci de neoformatie. La Protostomieni metamerizarea este foarte neta la formele inferioare si la stadiul de adult dar ea se estompeaza frecvent la formele superioare. Sistemul nervos al acestor Metazoare este condensat, metamerizat si cuprinde o pereche de ganglioni cerebroizi anteriori si dorsali, in relatie cu un lant nervos ventral cuprinzand ce o pereche de ganglioni per metamer, situati sub tubul digestiv. Aceasta dispozitie a sistemului nervos le-a dat calificativul de Hiponeurieni. Aici intra increngaturile: Annelida, Arthropoda, Mollusca, Sipunculida, Echiurida, Onychophora, Lofoforieni (Foronidia, Ectoprocta, Briozoa, Brachiopode).
La aceste organisme triploblastice celomate blastoporul devine anus iar gura este de neoformatie. Segmentarea nu este niciodata de tip spiral, iar mezodermul ia nastere frecvent prin enterocelie (Fig. 82).
Nivelul de diferentiere si pozitia sistemului bervos permite scindarea deuterostomienilor in: Epitelioneurieni si Epineurieni.
a. Deuterostomienii epitelioneurieni. La Echinoderme, Stomocordate si Pogonofore (oraganisme marine arhaice, sistemul nervos este in general incomplet despartit de ectoderm de unde de altfel el si deriva (de aici si numele de epitelioneurieni). Cefalizarea este practic absenta, metameria este estompata iar cordoanele nervoase raman la stadiul ganglionar. Celomul este scindat in 3 perechi de vezicule celomice.
b. Deuterostomienii epineurieni. Aici sunt cordatele (Tunicierii, Cefalocordatele si Vertebratele). Aici centrii nervosi sunt localizati deasupra tubului digestiv, exista o axa scheletica dorsala (coarda), situat intre tubul digestiv si sistemul nervos.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 13108
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved