| CATEGORII DOCUMENTE |
| Bulgara | Ceha slovaca | Croata | Engleza | Estona | Finlandeza | Franceza |
| Germana | Italiana | Letona | Lituaniana | Maghiara | Olandeza | Poloneza |
| Sarba | Slovena | Spaniola | Suedeza | Turca | Ucraineana |
| електронен | изкуство култура | икономика | история | книга | компютри | медицина | психология |
| различни | социология | техника | управление | финанси | химия |
Терминът е въведен през 1886 от немския биолог Ернест Хекел. В труда си “Обща морфология на организмите” той я определя като биологична наука изучаваща успоредните взаимоотношения на животните в окражаващата ги органична и неорганична среда. Етимологиата на термина екология включва гръцките думи oicos – дом, жилище домакинство, икономика и logos – значение, учение, наука.
Йоханзен 1959 г. “Основи на екологията” – общобиологична наука за закономерностите на взаимоотношенията на организмите и средата изучаваща начина на живот на животните и растениата и продуктивност, динамика на численоста на процесите и природата на биоценозите.
Дариус 1975 г. “Основи на екологията” – наука изучаваща условията на съществуване ан живите организми и взаимовръзките между организмите и средата която обитават.
Янкович 1972 г. “Екология” – изучава отношенията на организмите и растителните съобщества към условията на външната среда и връските между самите организми в биоценозите или екологията е наука за домакинството и икономиката на живите организми.
Траян 1975 г. “Обща екология” – икономиката на природата.
Одун “Основи на екологията” – строежа и функциите на природата. Това определия отговора на концепцийте на екосистемите и биосферата, които тои разглежда като глобална екоситема с основна функционална единица наречена екосистема. Екологията е биология на окръжаващата среда, иам за предмет същноста на връзките между организмите и тяхната среда.
Радкович 1983 г. – наука, иследваща закономерностите на жизнената дейност на организмите ( в успоредните им направления и на успоредни нива на интеграция ) в естествената им среда на обитание и при отчитане измененията в средата настъпващи от антропогенната дейност.
Федоров и Гилманов – фундаментално разклонение на биологията. Фундаменталните свойства на животните при надорганизмово ниво на организация. Екологията е наука изучаваща съвкупноста от живи организми взаимодействащи по между си и формиращи със средата на обитание единство. Система чрез която се осъществява трансформация на енергия и органични вещества.
Съществуват различия в гледните точки на отделните автори, но успоредно разглеждат екологията като наука изучаваща взаимовръзката между организмите и обитаваната от тях среда. Подечертават че организмите в природата не се срещат изолирано един от друг и средата, а заедно формират сложни системи биоценози или биогеоценози (екосистеми). Взаимодействието между организмите и обкръжаващата ги среда води до възникването на биологична среда, която е техен дом, жилище, местообитание и в нея се извършва развитието, преживяването и размножаването. Трактовката на някои автори за екосистемата като икономика са свързани с термините oicos . Широкото тълкуване на екологията изисква да се уточни обекта и даже в пределите на надорганизмовите нива на организация на биологичните системи.
Коя е елементарната единицата на екологията – индивид или популация? Ако се преме че елементарната единица (обект) е популацията то пренебрегваме възможното влияние на вътрепопулационата структура и взаимоотношения на самите популаций на междупопулационните влияния в границите на природните съобщества. Подобно схващане е неприемливо – трудно е да се определи от каво в по-голяма степен зависи структурата и функцийте на популацийте в биоценозите от вътрепопулационните взаимоотношения между индивидите. Така че елементарна единица в екологията е индивидът като елемент на популацията. Така че като функционална дисциплина екологията изучава популацийте (формирани от едно и разнородни индивиди), съобществата (ценозите формирани от едно или разнородни популаций), биоценози формирани от разнородните ценози и екосистемите като функционално взаимообусловено единство на боиценози и тяхната среда или биотоп. По-високите нива на организация (ландшафти, биоми и биосфера) са само отчасти обекта на биологията и екологията, тъй като е могата да се разглеждата без участието на географските дисциплини.
Главните подходи в екологията са системния и функционален подход . Системния подход включва три основни положениа:
Корените на екологията се крият още в древноста въпреки че като наука възниква в средата на 18 век и става самостоятелна на границата 19-20 век. Писменни данни има от Египет, Индия, Китай, антична Европа и др.. Други философи Емпедокъл, Аристотел, Теофраст, Приний, Хипократ имали наивни но сравнително правдиви представи за цялата формираща се и развиваща се пророда. Емпедокъл е писал за връзките между развитието на растенията и условията на средата. Хипократ издигнал прогерсивни за времето идеи, че факторите на средта влияят върху здравето на хората. Аристотел класифицира животните на базата на тяхното преживяване и хранене и т.н. Теофраст дава сведения за зависимоста между формите и особеностите в растежа при растенията и почвените и климатични фактори. Плиний в естествената си история характеризира различни явлениа в природата от различни екологични позиций.
3. Епоха на възраждането – характеризира се с подем на всички естествени науки и натрупването на биологични и екологични данни.. Инвентаризацията на живата природа е основна тенденция. В резултат на това се отедлят редица клонове на биологията, които са се формирали като самостоятелни науки. За края на периода са характерни трудовете на Линей поставил основите на растителната систематика. Жак Буфон – 13 тома “Естествена история” в която е обърнал голямо внимание на взаимодействието организъм среда. Ламарк – обръща внимание на значението на външните условия за еволюцията на организмите. Декандол – ботаническа геотрофия – описва се влиянието на биотични фактори върху растителните организми. По нататъшната диференциажия на биологичните науки е свързана с възникването на геотрофийте на растения и животни като самостоятелни дисциплини – голямо значение за развитието на екологията. Хумболт - “Идеи за географията на растенията” – разработени са понятия за изолиний, физиологически форми растения, асоциаций и други по същество екологични термини.
след Дарвин – характеризира се с обособяването на екологията на животните и растеният а като самостоятелни науки. За развитието на екологичните идеи е допринесла творческата дейност на :
и ландшафтознанието. Изследванията му дават силен тласък на изследванията свързани със взаимоотношенията растения почва;
последици върху природаната среда от дейностите на човека и необходимост от преосмисляне на човешката дейност върху природата и цялата биосфера. Екологията бавно се превръща в една от водещите биологични науки което е свързано с факта, че само от екологични позиций е възможно развитието на законите в биологични макросистеми, рационалното използване на ресурсите в биосферата. Въпросите свързани с решаването на редица екологични пробелми възникващи при НТП и свързани с поведението на човека от индустриалното общество в природата. От този период особено отношение имат редица разработки : Пиолевская - “Кратък курс по екология на растенията”;Шенников; Йохансен - “Основи на екологията”; Наумов – “Екология на животните”; Федоров и Гилманов – “Екология”; Пиърс – “Екология на животните”.
Взаимодействието на организмите и средата се изучават от много други науки, за разлика от тях в екологията се засягат тези страни във взаимоотношенията, които способстват развитието, размножаването и преживяването на индивидите, структурата и динамиката на популацийте и съобществата. На определен етап от развитието на човешките представи за природата е настъпило идейно сближаване на екологията с други биологични и небиологични науки. Особено тесни са взаимовръзките с физиологията успешно се развива направлението функционална екология. Между екология и морфология – направлението е ембриология. Екологията е тясно свързана и със систематиката в която са включени екологични критерий за класифицирането на организмите, а на екологията и е необходима точна система за точно определяне на изучаваните организми. Обединение на еволюционисти, генетици и еколози – изменението на екологичните механизми на еволюция в природата. На основата на екологията се развива биогеографията, молеоекологията и др. Екологията е тясно свързана и с много небиологични науки – география, хидрология, геоморфология, ландшафтознание, климатология, метереология, математика и др.
Огромна е ролята на екологията при разработването на редица теоретични проблеми свързани с миграцията на веществата и протичането на енергията в биосферата, с механизмите на еволюциония процес, със структурата и организацията на живта материя, с устойчивостта и управлението на макробиомите. Достиженията на екологията успешно се прилагат в медицината, ветеринарната медицина, провеждането на мероприятия за опазване на природата и рационалното и използване на природните ресурси. Развива се екологична икономика (на биоресурсите), инженерна екология и биотехнология, решаващи проблеми за отстраняване на негативните последици върху околната среда. Соцекологията – наука за опазване на екол. среда.
Проникавнето на екологични идеи във всички раздели на биологията – под въпрос е самостоятелността и като наука. Във всички съществувващи класификации е ясно видимо мястото на екологията сред останалите дисциплини – ниво на организация на биообекта – молекулярно – МБ, БХ; на клетъчно и организмово – тъканно – хистология, анатомия, физиология ; на организмово ниво – ботаника, зоология; в природата всеки вид утвърйдава себе си не като сума, а като единна функционална единица – популация. За йивите организми – биоценози, екосистеми и биосфера. Екологията изучавабиологичните макросистеми.
Йохансен : общи (систематика, морфология, физиология), частни (МБ, ботаника, зоология, антропология) и комплексни (хидробиология, аеробиология, почвознание) науки.
Общите биологични науки – целия организмов свят в строгоопределено направление ( “По малко за всичко”). Екологията е обща – фитоекология, зооекология, микроекология, на гъби, на водорасли, на насекоми, на горите и т.н.
Частни – всестранно конкретни обектина организмовия свят ( зоология изучава морфологията, физиологията, анатомията и т.н. на животните (протозоология, хелминтология и т.н.))
Комплексни – изучават условията на живот за организмите в петте основни среди. Най-задълбочено и всестранно развитие на биологични идеи доминира екологичния подход за изучаване (хидробиология изучава систематиката, морфологията, екологията, физиологията (общи науки) на растенията, животните, МО (частни науки) обитаващи водата.
Алтенология (altos – сам) Шрьодер 1856г. – за индивидите ( взаимоотношенията на индивидите със средата). В основата лежат морфофизиологични реакций към факторите на средата. Так започва всяко екологично изследване (биохимични рекций, водо и газообмрн; хистологични, цитологични, морфологични, анатомични, ембриологични, еколого- географски фактори)
Популационна – демекология (demos – народ). Популацията е елементарна надорганизмова макросистема. Основни проблеми - изясняване на условията при които се формира популация. Изучава се структурата, численоста и динамиката на популацийте.
Съществуват и други деления – мезоекология (характеристиките на средата на обитание); етологична екология (етос – нрав, поведение) - поведение и рекций на организмите в средата на обитание; хроноекология (последствия във времето) - връзка между изменението и динамиката в структурата и функцията на организмите.
Полеви методи – изследвания непосредствено в природата. Изясняват се структурата и функцийте на системите.
Аналитични методи (експериментални) – изясняват се причините и екологичните механизми обуславящи съществуването, функцията, динамиката и развитието на системите при отчитане циклите на динамиката на средата и вътрешната разнородност на макросистемите. На базата им – нови полеви и лабораторни експерименти. Комплекс от методи – БХ, физ., анатомични и др.. Способност на всички биосистеми към саморегулация – методи на теория на информацията и кибернетиката. Моделиране: в различни области живи модели или изкуствени със сходни свойства:
I тип реални (аналогови) – най-съществени черти на оригинала. Пример: аквариум – океан.
II тип знакови – условно отражение на оригинала, описание, концептуални (подробно описание) и математически (описание на структурата и функц. чрез математически символи. Процесът на прехода на физ. И биол. Представи за екосист. в математически зависимости се нарича системен анализ.
В съвременната екологията реални и знакови модели се използват паралелно.
Среда (околна, окръжаваща) – съвкупност от материални тела и енергия, заобикаляща биол. системи. Непосредственото обкръжение на биол. системи е среда на обитание (конкретно пространство). Съществуват 5 типа среди: водна; наземна; въздушна; почва; други организми ( за паразитите).
Различните елементи на средата имат разл. значение. Екологичните фактори са всички елементи на средта които въздействат на организмите, независимо от характера му – светлина, вода, радиоактивност и т.н.. С рзвитието на науката броят на екологичните фактори непрекъснато нараствал. Оказва се, че организмите реагират на шум, магнитно поле и др.. Под условия за съществуване се разбира съвкупността от жизнено необходими екологични фактори, обуславящи възможност за растеж, развитие, преживяване и размножаване – светлина, вода, температура, храна. За разлика от тях останалите екологични фактори не са непосредствено необходими за живота им – вятър и др..
Според Наумов условията за съществуване са:
Многообразни класификаций на факторите – разделянето на елементите зависи от целта на изследване и облекчава рабирането на връзката организъм среда.
Най-разпространената класификация на екол. фактори:
Климатични – светлина, топлина, въздух (състав и движение), влага (почва, въздух).
Върху водните организми – друг комплекс – хидрофизични и хидрохимични фактори.
Фитогенни (пряко или косвено влияние на раст. обитатели)
Друга класификация:
Друга класификация:
Подобно разделяне не е прецизно – един фактор може да е и пряко и косвено действащ – температурата влияе пряко върху растежа и косвено върху водния режим.
Най-приемлива класификация – на Роменски:
Въздействието на екологичните фактори върху организма е акция. Въздействието на организма върху средата – рекция. Взаимодействието организъм среда – коакция.
Ако някои от екологичните фактори се приближава към min си за дадена биологична система значи той действа ограничаващо и на останалите фактори, колкото и благоприятни да са те и така определят крайния резултат от въздействиетона средата върху биологичната система. Изменението на този резултат е възможен само чрез въздействие върху ограничаващия фактор – 1940г. Закон на Либих.
Светлинния режим е ограничаващ за върху тревистите растения под букови гори, където другите 3 режима са оптимални.
Либих е изследвалвлиянието на различни фактори върху културните растения и получил добияи от тях. Добивите се лимитират от хран. елементи присъстващи в почвата в минимални количества.
За успешното прилагане на закона на Либих има два важни принципа:
По нататъшните развития на теорията за влиянието на екологичните фактори върху биосистеми – Закон за Толерантноста на Шелфорд. Лимитиращо влияние не оказва само недостигът, но и излишъкът на даден фактор в средата. Всяка система има екологичен min и max на фактори при които мойе да съществува.
Въздействието на екологичните фактори зависи от интензитета им. Организмите реагират чрез промяна в реакцийте си. Рекцията може да бъде физиологичен процес, морфологичен показател, численост.
Зона на действието (на толерантност) се намира между min и max, при които е възможно съществуването на вида, популацията, съобществото. Съществува оптимална стойност на фактора друдно се определя, затова се говори за зона на комфорт. Крайните участъци изразяват угнетеност при недостиг или излишък на даден фактор – зони на песимум. В близост до min и max се намират сублеталните зони, а извън зоната на толерантност – летална зона. Екологичния фактор, чието ниво в количествено или качествено отношение е близо до зоната на песимума е лимитиращ фактор.
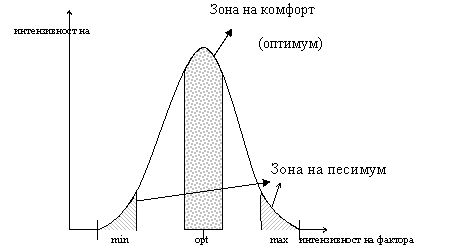
Условията на средат, при които някои от факторите имат стойности извън зоната на комфорта и оказват угнетяващо действие са екстремални – увеличаване или спадане на температурата, увлажняване или суша, тежки метали и т.н.. Различните биологични системи се различват по еколог. закономерност. Различията могат да са свързани с положението на оптимума. В студени местообитания оптимума е при ниски температури и обратно.
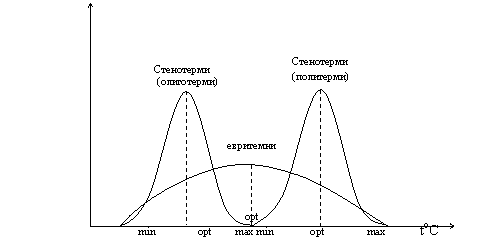
Няко организми предпочитат определен много тесен диапазон (тясна амплитуда) на фактора – стенобионти. А други предпочитат широк диапазон (амплитуда) на фактора – еврибионти : температура – стенотермни и евритермни; вода – стенохидридни и еврихидридни; соленост – стенохалинни и еврихалинни; храна – стенофагни и еврифагни; местообитание – стеноойкни и евриойкни.
Закон за толерантноста. Относителните предели на толерантност при стенбиотни и еврибиотни организми са различни. Например при стенотермните min оптимум и max на реакцията са сближени, така че малки разлики в температурата, които не биха оказали влияние на евритермните видове са критични. Стенотермните включват олиготермни – толерантни към ниска температура и политермни – висока температура. Графическо изразяване:
Формулирани са редица принципни допълнения към закона за толерантността:
Факторите на средата не действат изолирано, а съвместно. Дйствието на факторите до голяма степен зависи от екологичния фон – количествената изразеност на останалите фактори. В природата видовете съществуват в условия различаващи се от лабораторно определените диапазони на фактора. Ако стойноста на екологичния фактор е извън зоната на комфорта, може да се промени диапазона на толерантност на останалите екологични фактори. Например: ниското съдържание на N в почвата е свързано с намаляване на сухоустойчивоста на житните видове; тропическите орхидеи са сенколюбиви, но при по-ниска температура се развиват и при по-силна осветеност.
Известно е че може да настъпи в природата частично взаимозаменяне на екологични фактори.Например интензивноата на фотосинтеза се достига една и съща при силна осветеност или при голяма конц. на СО2 и при по-слаба осветеност. Нито един жизнено необходим фактор не може напълно да се замени.
Ако значението на един от режимите на средат е извън диапазона на тоелрантност за дадена биологична система, то съществуването и е невъзможно за даден екотоп. Фотосинтезиращите организми не мога т да съществуваъ при абсолютна тъмнина дори при оптимални други условия.
Различават се два оптимума – физиологичен или потенциален и екологичен действителен оптимум, съответно при присъствие или отсъствие на биотично влияние.
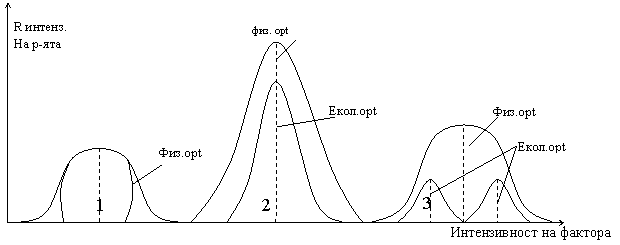
2 - намалява интензивността
При биотично влияние намалява диапазона на толерантност, запазва се интезитета на реакцията. Освен в оптимума се наблюдават различия в географско разпространение - различават се потенциално физиологичен ареал - изразява потенциалните размери на ареал.
Действителен ареал - наблюдава се в природата. В повечето случаи потенциала е по-широк от екологичния. Местообитания достъпни по екологични възможности за даден вид се оказват заети от конкурентно способни видове. Това води до изместване на екологичния оптимум и стесняване на ареалите.
4. Диапазоните на толерантност на видовете могат да се изменят в онтогенезата. За размножаваши се индивиди - семената, яаицата, поницити, личинки. Тези диапазони обикновенно са по тесни от тези на възрастните неразмнойаваши се индивиди.
Това е екологична отправна точка за изследване на макросистемите в природата. Чрез нея се обяснява разпространението, разпределението, взаимоотношението и интезитета на биологичните сиситеми в макробиосистеми и основни закономерности свързани с функционирането. При екологичните изследвания основно внимание се обръща на функциите на екологичните фактори в съответните етапи на развитие на системата. Ако за аддена биологична система е характерен широк диапазон на толерантност към определен ЕФ който е относително постоянен и умерено количествено изразен той със сигурност той не може да е лимитиращ и функц. важен за системата. При условие че разлеглажданата система има тесен диапазон на толерантност към изменчив фактор то именно той се разглежда при екологичните изследвания - може да се окаже лимитиращ. Установяването на лимитиращите фактори и отстраняване на лимитиращото им действие тоест оптимизирането на средата е важен за да са св.с изп.
Концепцията за лимитиращ фактор е отправна точка и при оценка на влиянието на човека върху околната среда. Основни етапи на оценката:
1. Установяването на функц. важните за една система еколог фактори, чрез нгледен анлиз и лабораторни експерименти.
2. Определяне на начините чрез които тези фактори влияят върху основните подсистеми в екосистемите - индивиди, популации и ценози.
3. Прогнозиране на изменениети на средата и съответните екологични следствия.
Основни стратегии на биологичните системи за тяхната преживяемост и приспособеност към средата са :
По този начин биол. системи отслабат лимитиращото влияние на екологичните фактори. Подобни компенсации са особено ефективни на ниво ценози но са възмойни и на видово ниво. например видове с широко географско разпространение почти винаги формират адаптации към местните условия популации - екотипове. Оптимумите и диапазоните на толерантност на екотиповете съответстват на конкретните условия компенсирането мойе да се съпроводи с появата на генетични раси. генетични раси могат да бъдат със или без морфологично проявени признаци или с поява на климатично приспособени индивиди без генетични изменения. Адаптацията (компенсация се постига от видовете чрез изменения на молекулно ниво - ензим субстрат, на клетката и органно ниво при поява на група животни които сезонно мигрират избягват неподходящите условия. Компенсирането на факторите на ниво ценоза се осъществява чрез смяна на видовия състав в зависимост от съществуващия широк или сезонен градиент на условията или чрез изменение на кръговрата на веществата. Непрекъснатост в бедните на хранителни елементи местообитания. Недостигът им мойе да се компенсира чрез активен кръговрат мейду хетеро и автотрофния блок. Например концентрацията на азотни съед. в открития океан е много ниска но наблюдаващата се активна фотосинтеза на фитопланктона. Недостигът на азот се компенсира чрез бързо поглъщане на бактерии.които разлагат мъртвата органика.
Околна среда - постояна динамика. основни причини за тези промени са космични фактори от които се определят денонощни лунни сезони и др (флуктоации) цикли, които са преодични или непереодично главно необратими промени.Цикличните и непереодични влияния заедно с необратимите изменения оформят сложната картина на динамика на почти всички компоненти на средата. Дори при строгат переодичност на денонощните промени и сезонните цикли не се повтарят напълно на периоди или на преодични колебания с голяма продължителност е свойствена още положителна регулярност - измененията на климата са големи периоди от време. Установен е редувания на топли и студени сухи и влажни периоди с продължителност 3-4, 6-7, 11,35,80-100 години.
По известна е 11 годишната переодичност свързана със слънчевата активност - намаляване на слънчевите петна. Това е съпроводено с повишаване ан радиацията, на озона в атмосферата задържане на топли излъчвания на земята и повишаване на температурата на повърхноста и. Същевременно се засилв и атмосферната циркулация - затопляне на големите и малките географски ширини. Възникването и развитието на живота е ставало на фона ан такава променяща се среда. Отрайение са ритмиката на биологичнитеявления - редуване на покой и активни периодичност в растежа и размножаването, колебания в размножаването структарата и численоста на популациите, ценозите. Най дълбоките промени са свързани с вековните промени на климата - разширяване на ареалите на едни и стесняване на ареалите на други видове. За да преживеят видовете се приспособяват не само към стойностите на екологичните фактори но и към переодичноста им. Принудени са да програмира жизнените си цикли така че да използват маx благоприятните периоди. Чрез влияние на индивидите в популационите съобщества и естествен отбор. Целите съобщества реагират на ритмиката в условията на средата. за разлика от много други сезонно изменящи се фактори дължината на деня навсякъде е една и съща. Амплификацията на сезонните изменения нараства със географската ширина. Това позволява на биологичните системи да отчитат не само времето през годината но и географската ширина. Фотопериодът е пусков механизъм за протичане на изменчиво модифицирани биологични процеси водещи до листопад, съзряване на леторастите и лигнифициране на дървесината, растеж или цъфтеж, смяна ан козината и оперението, натрупване на мазнини, скорбяла, миграции, размножаване, настъпване на диапауза при насекомите. Фотопериодът е универсален регулиращ механизъм свързан с механизма - биологичен часовник. Основна проява на биологичния часовник е циркулация, ритъм. Той доказва способноста на биологичните системи да разпределят във времето и переодически да повтарят функциите си дори при отсъствие на светлина при външен сигнал. Чрез биологичните часовници се постига съгласуване на физиологичната ритмика с ритмиката с факторите на средата. Чрез този механизъм биол. системи могат да предвиждат денонощните, приливно отливно и други изменения. В повечето случаи биологичните системи възприемат фото периодизма чрез рецептори или чрез листните пигменти. Те активират един или няколко важни механизми, включват хормони или ензими, в предизвикване на съответните физиологични и поведенчески реакции. Например съобразно влиянието на фотопериодизма върху растежа виреят 2 групи растения : 1. Растения на краткия ден - за настъпване на цъфтеж е необходим фотопериод по малък от 12 часа на ден; 2. Растения на дългия ден - повече от 12 часа.
Фотопериодизма може да контролира раждаемоста и размножаването в популациите. Дългите дни в началото на лятото предизвикват отделянето на неврохормон - диапаузни яайца, които се излюпват през пролетта на следващата година въпреки, че температурата и хран. условия са благоприятни. Съществуват и случаи, при които регулатор на растежа и развитието може да бъде и непероидични изменящи се фактори. Например валежите за пустинната флора, семената на повечето растения съд. инхибитор на прорастването, които може да бъде измит дори от минимални валежи, но с определена сила. Такива семена запазват жизненоста си в продължение на много години до подходящ дъжд. Ако се посеят в парници във влажна почва не покълват. Това е причина за превръщането на пустинята в цъфтяща градина след дъжд.
Значение на еколого-физиологичните режими за структурата и функционирането на биологичните макросистеми - популации, ценози, съобщества, е различно. Често лимитираща роля за наземните екосистеми има топлинния и овлажняващия режим, а за водни - топлинен, солеви режим и аерация.
Съотношението на топлинния режим и разнообразието на овлажняване определят климата на сушата - тясно взаимосвързани. Топлинния режим има по-изразен лимит. Влияе когато режима на овлажняване е близък до критичната си стойност и обратно. Взаимовръзката между двата режима - хигротермален режим, се изразява чрез 2 вида диаграми – климатограми:/липсват/
Режим на аерация – съдържанието на О2 и СО2 се изменя в дълбочина и зависи от фотосинтезата и дишането. епилимнион е наситен с О2 – в резултат на асимилация на СО2. В придънните слоеве, където се извършва окисление на органиката съдържанието на СО2 е високо. В олиготрофните езера (бедни на орг. В-ва) концентрацията на О2 е постоянна, а в еутрофните е сравнително ниска. Кислородът, отделян при фотосинтеза на водорасли в океаните достига до дълбочина зависеща прозрачността. Вълнението и смесването на водите има голямо значение за обогатяване на повърхностните слоеве с О2, като съдържанието му намалява до термоклина . Топлите води съдържат по- малко О2 от студените. Обикновено в океаните се наблюдава високо съдържание на О2 под 1000 м – бавни топли течения от север на юг. Възможно е пълно отъсъствие на О2 в дълбоките слоеве – Черно море под 150-2200м анаеробно разлагане на органични с-я с отделяне на H2S.
Съдържанието на разтворими соли е различно, в зависимост от това – пресни и морски води. Пресните съдържат до 0.5 гр/л соли, които са с много променлив състав. Морските са до 35 гр./л и са с постоянен състав на солите. В пресните води преобладава СО32- до 80 %, а в морските – Cl- средно около 89% и SO42- до 11%. Впресните води най-много се съдържа Са2+ - 64%, Mg++ - 17%, Na+ - 16% u K+ 3%. Количеството им зависи от скалната подложка. Водата в езера на варовит терен – до 800 мгр/л суха утайка, а тези на силикатен терен под 50 мгр/л. В богатите на Са2+ зелените растния поглъщат СО2 при фотосинтеза и предизвикват натрупване на СаСо3. Такъв е произхода на туфите в ручеите, калциевите езерни глини и калциеви включения във водораслите например Cladophora. В зависимост от съдържанието на Са2+ - пресните са меки – под 9мгр/л и твърди 25мгр/л.
Хлоридите, ако се съдържат в прясната вода, попадат с дъжда (морски произход) , а SO42- - в гипсоносни пластове .
В морските води се срещат почти всички елементи. Средната соленост е 35%, като в открити океани е 33-37%, повечето е солеността на тропическите морета поради високото изпарение и по-малко в полярните морета, които се обогатяват с прясна вода от там.
Концентрацията на соли в епиконтиненталните морета е по-висока поради по-голямото изпарение - 41%. При ниска на изпарение солеността е по – малка, в Черно море 18%, Балтийско 12%. Солеността намалява в близост до устията на реките. В зависимост от органичните в-ва се различават:
Съдържанието на органика има +/- значение. Може да съдържа N, P, хормони, токсични в-ва. Средното съдържание на органика N, P, е 0.05мгр/л. Морската вода съдържа средно 0.01 мгр/л Р и 0.02мгр/л N. Вдистрофни езера и водоеми (мъхови формации) се съдържа значително количество кафяви хумусни в-ва.
Киселинността е във връзка с разтворените СаСо3. Във варовитите води рН е 7-8.5, а в повърхностните 8-8.3 ( морски води).
Върху киселинността влияят и организмите чрез дишане и фотозинтезата. Има денонощна динамика на процесите, която влияе върху рН. В еутрофната зона през деня рН е високо (Фотосинтеза), а през нощта е ниско (дишане). Тези промени са незначителни в силикатни или слабо солени води. В кисели води с рН по-ниско от 3.5 се развиват мъхове от род Sphagnum и ларви на някои двукрили. При рН до 7.5 раста папрати от род Isoetes - шилолист. В слобо кисели води рН 7.2 – 7.7 растат вода чума, теснолист папур и др. Рибите рН 5-9 max 6.5 – 8.5 . За някои членестоноги е важно повърхностното напрежение на водата. За чистата вода е 76 дини при 0оС, а при 20 оС 73 дини/см3. Редица насекоми имат тегло по-малко от повърхностното напрежение и се задържат на водата – род Geris (водомерки) и двукрили. На повърхността на морската вода рядко, защото пяната понижава повърхностнотао напрежение.
Движението на воднота среда има разностранно значение. По- обилно снабдените с хранителни в.ва улесняват топлообмена, аерацията и газообмена.
Според типа на водообмен :
Хидатофити – жизнените процеси протичат при постоянен котакт с водата (жизнена среда) нямат приспособления за регулиране на водообмена и не се променя в онтогенезата. Приемането и отдаването на вода е физичен процес. Древните нисши растния в световния океан са типични, от тях са възникнали съвременните. От древните хидатофити, преминали на сушата са възникнали древните 2) сухоземни водни лишеи, мъхове, висши спорови и теснолистни папрати. Те са запазили примитивната организация и водообмен на 1) . Поглъщат вода с цялата повърхност на тялото, нямат приспособления за регулиране на водообмена. За разлика от тях понасят обезводняване – извършва се гелификация на протоплазмата, структурата и процесите не се разр. Пойкилохидрови свойства имат спорите и семената – сухоустойчиви
Пойкилохидрови – ефективно използват водата. При кратковременно овлажняване извършват интензивно дишане. Не променят водообмена си.
Хомеохидрови – повечето папратовидни, цветните и голосемените растения. По- слабо сухоустойчиви от пойкилохидровите. Притежават специфични тъкани и органи за регулация на водообмена. Редуват се и трите типа водообмен в онтогенезата им.семената се развиват при високо водно съдържание в цвета, а след това в плода – сходен водообмен с хидатофитите. Водообмена на възрастното раст е като при хомеохидровите. В узрялото семе се извършва преход от 1 към 2, който е автономен. Поникването на семето е преход от 2 към 3. Хомеохидровите имат много по- голяма водозадържаща и водорегулационна способност.
Огромното разнообразие от растителни видове се оформя от 3. Според набора от приспособления за регулация на водообмена хомеохидровите се делят на няколко екологични групи:
Флората на пресните води е формирана от гликохидатофити и гликохидрофити и се отличава с по- голямо видово разнообразие на висшите растения и широк географски ареал.
Флората на моретата и океаните се формира от халохидатофити и халохидрофити. Има огромно разнообразие от водорасли, висшите растения са около 30 вида обединени в 2 семейства:
С-во Potamogetonaceae
С-во Hydrochatitaceae
У нас Zostera marina образува подводни ливади.
Екологични групи растения : хидрофити
1) Потопени във вода – същински хидрофити и аерохидрофити. При някои видове цветовете са над водата.
А) Невкореняващи се – p Orticularia (мехунка), роголистник, някои видове водна леща, алдруванде (насекомоядно)
2) С плаващи стъбла – плаващи аерохидрофити
А) Невкореняващи се лейка Slvinia natans, водна лещажабешка водянка.
Б) вкореняващи се – род Nymphaceae, Nympha, Potamogeton.
Водните растения имат основни приспособления за живот във водна среда.
Увеличена повърхност на листата при неголям обем, това води до увеличаване на свободната повърхност за поглъщане на слабо интензивна радиация, разтворените в-ва , минерални в-ва и СО2. При редица видове листните пластинки са разсечени на дялове – мехунка, род Cekatophyllum Листните пластинки са тънки състоящи се от 2 или 1 слепени клетки. Потопените листа нямат диференциация на палисадна и гъбчеста т/н на мезофила и нямат устица. На отделни места има групи клетки с тънки клетъчни стени наречени хидропоти – за поглъщане на вода и минерални в-ва.
Хетерофилия (разнолистие) – По –голяма пластичност на листата в зависимост от режимите на средата. При някои амфибиини растения на едно и също стъбло има различни преходи от потопени до въздушни листа. Например Sagitalia sagitifolia (стрелолист). При други видове Salvinia, Nympha потопените листа рязко се отличават рязко от тези на повърхността. При някои видове роголистник причина за хетерофилията е температурният режим – през зимата по- широки и твърди листа.
Централно разположение на механичната т/н, това е свързано с по- голяма гъвкавост и устойчивост на движение на водната среда. Например водораслите на р Macrocystis u Nereocystis имат талус до няколко десетки метра . Централната му част се състои от клетки със задебелени стени – механична функция. Те устоявят на океанските бури.
Редукция на кореновата система при невкореняващите се или загуба на основната и функция при вкореняващите се. При едни видове служи за прикрепяне към субстрата, слабо разклонени без коренови власинки. При родове Nymphea u Nympha се формират мощни коренища главно за отлагане на хранителни в-ва и за вегетативно размножаване
Липса на транспирация при потопените видов. Възходящ воден ток при подводно- подводни видове се осъществява чрез интензивна транспирация на въздушните листа. Устицата се затварят само през нощта.
Водните животни приемат елементи от околната среда чрез консумация на фитомаса или осмоза като съществува непрекъснат водо и солеви обмен между тях и околната среда. Редица водни организми имат избирателна способност да натрупват някои елементи в тялото си. Средно вбиомасата на водните животни има хиляда пъти повече, спрямо воднатa среда, P, Si u Zn; 100 пъти повече S, Fe, Cu u I и 10 пъти повече K, Br u F.
Някои водорасли натрупват до 20% от теглото си Fe. На І са богати коралите, червени и кафяви водорасли–те се използват и за промишлен добив на І
Така водните организми изменят химизма на водоемите и натрупват органични утаечни материали – глини. Подържането на необходимото за обмяната на в-вата осмотично налягане при животните се осъществявя чрез регулация на водно- солевия обмен. По характера на солевия обмен 2 типа животни:
Болшинството от морските безгръбначни са слабо хипертонични: повече соли в организма спрямо средата. Има и хипотонични видове ракообразни. Изотонични са мешестите и иглокожите.
Хомеоосмотични – пресноводни безгръбначни и всички гръбначни.
При пресноводните видове запазването на по- високото осмотично налягане става чрез непрекъснато отделяне на вода. Напълно се обезводняват в солени води следователно не съществуват амфибии.
Концентрацията на солите се регулира от отделителната система с урината. При рибите урината е по- хипертонична от кръвта. Чрез хрилете поглъщат или отделят вода. Така се подържа необходимата соленост на тъканите. За акуловите риби е характерно подържането на изотония чрез задържане на урината в кръвта.
Хомеоосмозата при много животни е свързана с развитие на непроницаемост на покривните тъкани – хитин, рогови образувания, слизести в-ва. Високата соленост на морската вода е причина за достигане на големи размери и по- малка подвижност. Син кит 33 метра, акули 15метра, морски калмари 18 метра. При различна соленост се наблюдават различни размери. При рака Artemia salicha се наблюдава различен строеж на търбухчето при различна соленост. При някои риби се наблюдава увеличаване на броя на прешлените при повишаване на солеността.
В зависимост от толерантноста на организмите към разликата в солеността:
Режимът на овлажнение плюс температурният режим имат лимитиращо значение за структурата, функцията, географското разпространение и сезонната ритмика в развитието на организмите и съобществата им. Наземните организми са по- малко зависими от водната среда. Протичането на жизнените им процеси се извършва само във водна среда – клетки и междуклетъчно пространств, т.е. е вътрешна среда. Затова непрекъснатият водообмен с околната среда е жизненено необходим процес осигуряващ стабилна вътрешна среда. Водообменът зависи от режима на овлажнение в отделите екосистеми и за негвото протичане съществуват разнообразне адаптивни способности. Режимът на овлажнение на средата се определя от баланса на влагата – приход (валежи и кондензация) и разход (изпарение от почви, реки, водоеми и транспирация на фитоценозата) количеството на валежите зависи главно от пятя и характера на преместване на въздушните маси. Духащите от вътрешността на континента сухи ветрове не носят валежи. Така в крайбрежните райони могат да съществуват полупустини и пустини.
Важен фактор е разпространението на валежите по сезони. За България валежният максимум е през май – юни, а минимума август- септември. Важна количествена характеристика е месечната/годишната сума на валежите в мм.екологично значение за наземните групи и съобщестма има влажността на въздуха; изпарението и транспирацията зависят от температура , вятър, валежи.
Местните модификации на режима на овлажнение са свързани с условията на релефа, почвообразуващата скала и почвите. Тези екотопични условия са причина в един район да се срещат различни типове екологични системи. Когато при движението си влажните въздушни маси срещат планински масив, те се охлаждат и изкачват по склоновете и падат обилни валежи. На противоположните склонове количеството на валежите е ниско и съществува т.нар. дъждова сянка. Източните и западните склонове на остров Мадагаскар - върху източните склонове има горско тропична растителност, а по западните саванни екосистеми.
Върху местният режим на овлажнение оказва влияние и формата на релефа. Най- малко влага в горните, а най- много в понижените части на релефа. Склоновете с южно изложение имат по- неблагоприятен режим на овлажнение спрямо тези със северно.
Почвообразуващата скала оказва влияние върху баланса на влагата. Песъкливи и варовити скали по- добре пропускат дъждовната вода в сравнение с масивни силикатни.
Ако валежите са обилни, но подпочвената скала не задържа вода, то за екосистемите режимът е неблагоприятен.
Режим на овлажнение на почвата, валежи, повърхн. де/алувиален сток, дълбочина и разположението на подпочвените води, механичният състав на почвата, наличието на органика, биологично потребление на вода.
Основни форми на вода в почвата:
Гравитационна – стича се по големите почвени пори и подхранва подпочвената вода.
Непромивен (непермациден) – почвите не се навлажнявят в цялята си мощност. Между овлажненият от валежите почвен слой и подпочвените води има слой почва с нищожна влажност близък до увяхването т.нар. мъртъв хоризонт на изсушаване с дебелина от няколко dm до няколко m
Изпотяем (ексудационен) – по капилярен път сеовлажняват горните почвени хоризонти с подпочвена вода.
Водозастоял – подпочвените води са близо до повърхността такива са безоточните или слабооточните заблатени равнини.
Режимът на овлажнение е изменчив през годината и през много гдини. В зависимост от динамиката му през вегетационния период – следните местообитания.
Динамиката на овлажнение през годината, разликата в режимите на овлажнение на отделните почвени хоризонти са свързани с явлението екологична съвместимост на видовете според овлажнението т.е. съществуването в една биоценоза на растения приспособени към различни режими на овлажнение. Има 4-ри типа екологична съвместимост:
Понятието екосистема е въведено от Волтерен и Тенсли 1935г. Най- пълно е дефинирана от Одун 1971г.:” Екосистема е всяко природно единство, включващо всички организми (т.е. съобщества/биоценои)на даден участък, което взаимодейства с физичната среда по такъв начин, че протичащата енергия създава определена трофична структура , видово разнообразна и определя тип кръговрат на в-вата между биотичната и абиотичната част на това единство.” Т.е. според Одун екосистемата се състои от 6 основни компонента:
Понятието биогеоценоза, което се смята за синоним на екосистема е въведено от скандинавско – руската школа от Сугачев1940г.: “ Биогеоценозата е съвкупност от еднородни, на известно разстояние от земната повърхност природни явления (атмосфера, почва, подпочвена скала, хидрологични условия, растителен и животински свят и микроорганизми), имащи своя особена специфика на взаимодействие между съставящите я компоненти и между тях и други природни явления и характеризираща се с определен тип кръговрат на в-вата и величина и характер на протичащата през нея енергия, представлявящото вътрешно диелектрическо единство в постоянно движение и развитие”. Според Сукачов основните подсистеми на екосистемите са:
Екотопът включва цялата материална система на мъртвата част на биогеоценозата с всички свойствени на нея процеси и взаимодействия с биологичната част.
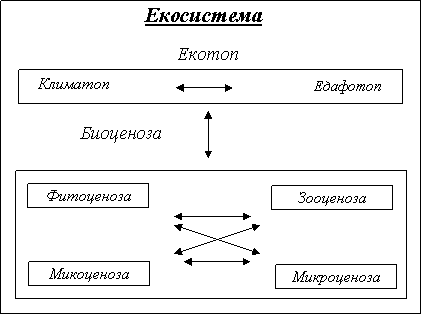
Както се вижда от определенията екосистема и биогеоценоза са много близки.
Биогеоценозата има конкретен характер. Системата в този случай е с определени граници, тип кръговрат и величина на енергията. Основателят и е изследвал главно наземни единства където структурните особености и връзката структура – функция са най – добре изразени.
Привържениците на понятието екосистема са изследвали главно водни екосистеми, където по- добре се разкриват функционалните връзки и по- слабо морфологични структури. Като екосистеми могат да бъдат разглеждани системи с различна големина: отделни малки водоеми, океан, биосфера – глобална екосистема. Условия за екосистема - да съществува равновесие и да се извършва кръговрат на в-вата в резултат на протичането на енергия.
Екосистемите от съвременна гледна точка са макробиологични системи със слажна структурна организация, при което благодарение на съществуващите функционални връзки между двете подсистеми ( екотоп и биоценоза) и между формиращите ги основни елементи се осъществявя:
Осъществява се вътрешна саморегулация на системата (саморегулиращ се хомеостазис) на основата на множество намалявщи енергетични главно отрицателни обратни връзки
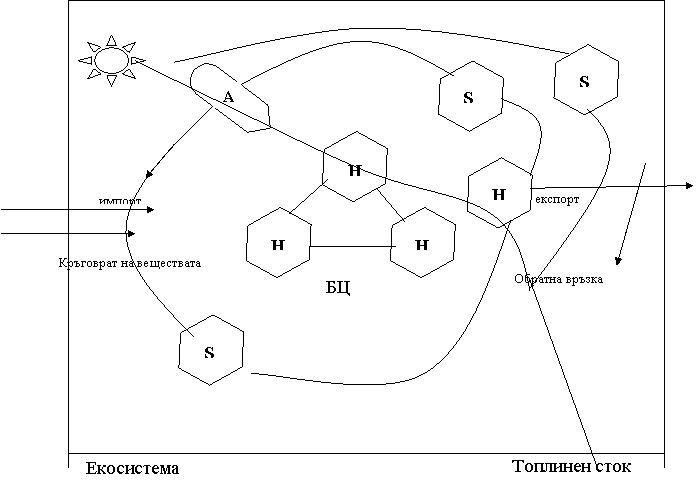
S – резервни хранителни в-ва.
Потокът енергия, минаващ през екосистемата параболоидно намалява, минавайки през хранителната мрежа от А и Н, тъй като част от енергията се включва в биоценозата катно енергия на хранителните в-ва. Този процес съгласно двата закона на термодинамиката е свързан с превръщането на по- голяма част от енергията в топлина и разсейването и в пространството чрез топлинниа сток, т.е. енергия може да се запасява, освобождава, да се включва в импорта и експорта, но биоенергията се използва еднократно – няма кръговрат.
S за разлика от Е могат да се използват многократно – кръговрат на в-вата. Ефективността на повторното използване, размерите на импорта и експорта зависят от типа екосистема и от ресрсите на местообитанието.
Различните екосистеми са разлино отворени т.е. за тях е жизнено необходимо постоянно постъпване и отдаване на енергия. За това при екосистемните изследвания е необходимо разглеждане на качеството и промените на средата – среда на изхода и среда на входа.
EC= IE+OE
IЕ – input env.
OE – output env.
Подобно разглеждане на екосистемите решава проблемите с очертаване на границите им. В много случаи те са условни, независими дали са естествени (бряг) или административни (град). Изменението на средата на входа и на изхода е различно и зависи от :
размер на екосистемата - колкото по- големи са размерите, толкова по- малка е зависимостта от средата.
При изследване и изучаване на екосистемите се прилагат два основни подхода:
Холистичен (holos, цел) – Измерва се постъпването на входа и отдаването на изхода на в-ва и енергия. Екосистемата се разглежда като единно цяло. Поведението на отделните екосистеми не се изследва.
Мерологичен (meros, част) изследват се свойствата на отделните елементи на системата, след което даннните се екстраполират за цялата екосистема.
Избирането на подход зависи от целите на изследване и степента на взаимодействие на елементите на екосистемта. Колкото по- силна е връзката, толкова регулациятя на системата е по- добре изразена следователно новите свойства по- добре се проявявят на ниво екосистема и не могат да се изразят чрез мерологичния метод. От друга страна едни и същи популации в различни екосистеми имат различна структура и поведение, което се пропуска обикновено при холистичния метод следователно двата подхода се допълват.
При екосистемите се използват експериментални методи. От функционална гледна точка е целесъобразно екосистемите да се изучават в следните направления:
Основната и най- важната функция на екосистемите е биологичният кръговрат. Основните движещи сили на биологичния кръговрат са енергийният поток и живите организми (биоценозите). В резултат на биологичния кръговрат на в-вата и потока на енергия през биосистемите с възникване на живота на Земята основно са сепроменяли съществуващите геосфери. Формираната биосфера се намира в равновесно състояние, подържана чрез многобройни регулаторни механизми основани на множество намаляващи енергетични обратни връзки В резултат на това се запазват благоприятните условия за съществуване, функциониране и еволюция на отделните биосистеми. Например забавяне на процесите на пълна хетеротрофия и утилизация и разлагане на продуктите на автотрофния метаболизъм в сравнение с процесите на тяхното създаване, обуславящи подържането в атмосферата на високи концентрации кислород и сравнително ниски концентрации на СО2, което е необходимо условие за съществуването на крупните аероби.
Биологичният кръговрат се осъществява чрез протичането на три основни групи процеси:
Продуктивността е два основни вида – първична и вторична.
Първичната продуктивност на икосистемите, на съобществата или на част от него е скоростта, с която се усвоява слънчевата енергия от продуцентите в процесите на фотосинтеза и хемосинтеза и се натрупват под формата на органично в-во. Първичната продуктивност е три вида:
Вторична продуктивност (асимилация) – скоростта на натрупване на енергия от консументите създадена от продуцентите. Част от енергията се изразходва за дишане , а друга част от енергията за собствените си тъкани.
Продукцията е количествена характеристика на свойството продуктивност. Продукцията е количеството синтезирано в-во от единица площ (при макропродукцията) единица обем при микропродуценти, микроконсументи или единица живо тегло при макроконсументи.
Продукцията се изразява в теглови единици – гр., кг., т. Или енергийни единици (J, KJ, cal) за измерването и могат да се използват теглови методи – макроконсументи и макропродуценти, газови и въглеродни – микропродуценти и микроконсументи. В основата на първичната продуктивност стои процесът на фотосинтеза, който от химична гледна точка се изразява чрез химична реакция от окислително- възстановителен тип.
![]()
![]()
![]()
За макропродуценти –зелени растения и водорасли А е О2, т.е. извършва се окисление на вода при освобождаване на О2, а СО2 се възстановява до ВВ и се освобождава вода.
При бактериалната фотосинтеза Н2А може да е неорганично съединение (H2S – зелени и пурпурни бактерии - серни) или органично съединение (пурпурни или кафяви несерни бактерии)Не се отделя кислород. Бактериалнатафотосинтеза играе незначителна роля в продукцията на органично в-во. Продукцията на серните бактерии е 34% от годишната продукция на органично в-во в целия обем. Екологичното значение на тази група е свързано с това, че бактериите могат да функционират при условия неблагоприятни за зелените растения. При тези условия участват в кръговрата на в-вата и играят важна роля за очистване на замърсени водоеми.
Съществуват три основни пътища за възстановяване на СО2 до въглеводороди.
Цикъл на Калвин (ПФЦ) - при С3 растенията съществува при сродни условия – голяма конкурентна способност в смесени съобщества и преобладаване в биоценози. Тази група създава по- голямата част от първичната продукция на земята.
Цикъл на трикарбоксиловите киселини – С4 растения. Видовете от семейство Poaceae и редица видове от пустинната, степната и тропическата растителност.
САМ метаболизъм – характерен за сукулентните растения (у нас Crassulaceae)
Хемосинтезиращите бактерии (хемолитотрофи) използват енергия за възстановяване на СО2 от хем, окисление на неорганични в-ва - NH3, NO3-, S2-, Fe2+, Fе3+. Редица автори смятат, че те участват при вторичното ползване на на въглерод – не са продуценти. Значението им е свързано с обогатяване на субстрата с минерални елементи и ползване на енергия недостъпна за консментите. Повечето са аеробни и съществуват при отсъствието на светлина.
Висока първична продуктивност се налага там, където са благоприятни и особено при наличието на допълнителна енергия, постъпваща в системите, което се нарича енергиина субседия. Всички източници на енергия, които намаляват загубите за самоподържане на системите и увеличават стойностите на продукцията се нарича енергиина субсидия. Например енергията на вятъра и дъжда в тропическите гори, енергията на приливите (естествена субсидия), енергията на изкопаемото гориво, енергията на мускулната сила (изкуствена субсидия) – за агро екосистеми.
Първичната субсидия е неравномерно разпределена както на сушата, където лимитиращи фактори са режим на овлажняване и топлинен режим, така и в океана където зависи от дълбочината (светлинен и топлинен режим) и режим на хранителни в-ва. Според общите изчисления годишната продукция на земята е 1.1018 kcal/m2. Разпределението на тази продукция по типове екосистеми е:
между 10 – 25 kcal/m2/у за естуарие, коралови рифове, равнинн екосистеми на алувиални почви и някои агро екосистеми.
между 3 – 10 kcal/m2/у – за влажни горски съобщества, ливадни, плитки езера и повечето агроекосистеми.
между 0.5 – 3 kcal/m2/у – за пасищни, някои селскостопански култури, дълбоките езера, високо планинските горски съобщества, водата на континенталния шелф.
Под 1.103 kcal/m2/у открит океан
Под 0,5.103 kcal/m2/у – пустинни съобщества.
По- голямата част от синтезираните органични в-ва не се консумират веднага, а постепенно преминават в постилката и почвата или постепенно се утаяват като мъртво органично вещество във водните системи, създавайки енергетичен базис на детритната хранителна верига. Детрит в геологията са продуктите от разрушаване на горните В областта на екологията под детрит се разбира органично в-во, подложено на разлагане, протичащо по детритните хранителни вериги. В детрита се включва неразтворимото органично в-во и разтворимото органично, което се измива или се извлича от сапрофитите от живите и мъртвите тъкани. Разлагането на органичната материя е резултат както на абиотичните процеси (степни и горски пожари), така и на биотични процеси, при които в атмосферата се връщат СО2 и други газове, а в почвата – минерални елементи.
За някои екосистеми от огневи тип (тип чакарал, степни) пожарите са важен регулаторен и лимитиращ фактор, тъй като микроорганизмите – деструктори не успяват да разложат органичното в-во. Механичното раздробяване на органичните остатъци също може да се извърши от абиотични фактори – ниски температури, вода, но и от микроконсументи и копрофаги. Окончателното разлагане се извършва основно от хетеротрофни микроорганизми – сапрофити. В широк смисъл разлагане означава биологично окисление, при което се отделя енергия. Съществуват два основни типа окислителни процеса:
Аеробно дишане, прикоето газообразният кислород е окислител/акцептор на електрони. Този процес е обратен на фотосинтезата, при който органичните в-ва се разлагат до вода и СО2. Освободената енергия е необходима на биологичните системи за включване на част от в-вата в клетките и тъканите. Всички висши растения и животни и Monera – Bacteriophyta & Cyanobacteria & Protista получават енергия по този начин.
Често окислението не протича до край – образуват се органични в-ва съдържащи част от енергията и доокисляващи се от други организми.
Desulpho vibrio – възстановява SO42- go газообразен H2S в Черно море. Сяроводорода в повърхностните слоеве може да се използва от фотосинтезиращи бактерии.
Много бактерии са факултативни анаероби. Продуктите при аеробното и анаеробното окисление са различни и освободената енергия при анаеробнто оксление е значително по- малко.
В бактериалните клетки и гъбния мицел съществува набор от ензими, необходими за протичането на специфични биохимични реакции. Те се отделят в мъртвото органично в-во . някои от продуктите на разграждането се използват от сапрофити, други остават в средата като енергиен източник на други сапрофити. Пълното разграждане на в-вата се осъществява от различни видове сапрофити, действащи впоследствие.
Интензивността на разлагане зависи от:
Разлагането протича през три основни етапа:
І) Раздробяване на детрита и освобождаване на разтворено органично в-во. През този етап основна роля играят малките животни, формиращи мезофауна – първаци, почвени кърлежи Nematoda, Polychaeta, личинки на насекоми. Те не могат да разлагат целулозните компоненти, а се хранят с детрит, получават енергия он микрофлората. Екологична роля:
Внасяне в средата на белтъци, хормони и други стимулиращи развитието на микроорганизмите в-ва.
ІІ) Сравнително бързо образуване на хумуса и освобождаване от сапрофитите на допълнително количество разтворено в-во. Хумусът и хумусоподобните в-ва са устойчиви на по- нататъшно разлагане. Те са задължителна част на екосистемите и играят важна екологична роля. Бавните темпове на разлагане на хумуса имат важна екологично значение – натрупване на СО2 в атмосферата – поради забавяне на процесите на разлагане и освобождаване на СО2 в сравнение.
Физичните свойства и химичният състав на хумуса малко се различават в различните екосистеми. За сега съществува само предположението, че морските хумусни в-ва имат различен строеж поради различния си произход.
Хумусът е тъмно, жълто – кафяво аморфно колоидно в-во с недостатъчно добре описан състав. Продуктите от кондензацията на ароматните съединения (феноли) с продукти от разлагане на белтъци и полизахариди. Бензолното ядро от фенолен тип и дългите му странични вериги са причина за устойчивостта му на микробно разлагане. За разлагането му са необходими специфични ензими – оксигенази, които отсъстват при широко разпространените почвени и водни сапрофити. Редица антропогенни замърсители – хербециди, пестециди, промишлени отпадъци са производни на бензола, трудно се разлагат и са силно токсични. Най- вероятно разлагането на подобни съединения е възможно в резултат на абиотични химични процеси и дейността на специализирани сапрофити.
Освен разлагане в природата може да се извършват и процеси на фосилизация на органичните в-ва, които протичат в два етапа:
Много бавен етап на карбонификация при анаеробни условия, водещ до последователното образуване на лигнитни, кафяви и каменни въглища, при което концентрацията на въглерод се увеличава.
Детритът и хумусните в-ва играят важна роля при формиране на плодородните почви. Присъствието на тези продукти в умерени количества е важно за структурата на почвата и подържането на благоприятен за продукцията режим на хранителни в-ва. Органичните в-ва могат да образуват комплекси с минералните елементи, които в някои случаи облекчават усвояването им, а в други намаляват токсичното им действие – например в резултат на хелатирането – образуване на комплекси с йоните на металните се намалява токсичното действие на неорганичните соли на тези метали.
Разлагането на органиката е дълъг и сложен процес, чрез който се контролират седем важни функции на екосистемите:
Производство на енергия за последователните родове в детритната верига.
Преобразуване на инертните тела на земната повърхност и формирането на почвата като биокостно тяло
Преносът на енергия в екосистемете от автотрофите през различните организми на основата на взаимната консумация се нарича хранителна верига. При всеки пореден пренос на енергия по- голямата част от потенциалната енергия се губи (средно между 80- 90%) под формата на топлина, следователно колкото по- къса е хранителната верига толкова повече е количеството, достъпна за популациите енергия. Хранителните вериги са два основни типа:
Двата типа хранителни вериги са тясно свързани, формирайки хранителни мрежи. Организмите, получаващи енергия от слънцето, преминала през еднакво количество степени, заемат едно и също трофично ниво (група от организми). Например зелените растения заемат първо трофично ниво – продуценти. Тревопасните животни второ трофично ниво – първични консументи. Първичните хищници заемат трето трофично ниво – вторични консументи и т.н.
Всяка популация може да заема едно или няколко трофични нива в зависимост от използваните източници на енергия, тъй като при преминаване на енергията през всяко трофично нива около 80% от нея се разсейва като топлина, достъпната енергия за всяко следващо ниво намалява. В резултат на това намалява и броя на организмите, които могат да се подържат от тази енертия т.е. намалява числеността и, а се увеличават размерите на отделните индивиди. По- големите размери са свързани с по- нисък интензитет на обменните процеси и следователно с по- малко количество необходима енергия. Например най- простите наблюдавани хранителни вериги са тези в тундровите съобщества. Те са с малко видово разнообразие – кратък вегетативен период и ниски температури. Първо трофично ниво – Cladonia, Poaceae, Junkaceae, Salix – фитофаги – карибо и северен елен, тундрови патици и леминги.
Трофични нива на хищници – северни вълци, полярни сови и човек. Основна храна на вълците и човека са елените, а на полярните сови - лемингите. Всички промени в числеността на популациите на елените и лемингите се отразяват върху функционирането на всички останали нива нива. Първичната продукция и числеността на продуцентите се регулира чрез есетествена миграция на стадата от карибо и северен елен. Миграцията на северния елен е направлявана от човека. Интродукцията на северния елен в аляска води до деградацията на пасищата и намаляване на достъпната първична продукция.
Във водоемите най- често продуценти са планктона и водораслите. Фитофаги – зоопланктон, безгръбначни животни, риби и бозайници; хищници – безгръбначни и риби.
Колкото по- дълги са хранителните вериги, толкова по- малко вторична продукция може да се използва от човека поради намаляване на енергията от ниво на ниво. Затова често при изкуствени зарибявания се предпочитат къси хранителни вериги – 2-3 вида растителноядни риби.
Дължината на хранителните вериги зависи от:
при които има недостатъчно първична продукция и дълги вериги.
Същиствува хипотеза за четния и нечетния брой трофични нива.: Ако една хранителна верига е с бедно функционално разнообразие и се състои само от две трофични нива – продуцентите и фитофагите са освен това с малко видово разнообразие. Количеството първична продукция ще се лимитира от фитофагите, а числеността им от достъпнат първична продукция. Други регулаторни механизми не съществуват и няма възможност за формиране на дълги пасищни вериги.
Ако хранителната верига се състои от три трофични нива – продуценти, фитофаги и хищтници с изразено видово разнообразие то числеността, то числеността на фитофагите ще се регулира от хищниците, а първичната продукция ще зависи от ресурсите на местобитанието. Това дава възможност за формиране на по- дълги хранителни вериги при налично видово и функционално разнообразие.
Количеството първична продукция ще бъде почти постоянна величина въпреки промените в режимите на средата поради видово разнообразие на продуцентите и различна екологична валентност и пластичност на отделните видове.
В резултат на протичането на енергия по хранителните вериги всяко съобщество придобива определена трофична структура, която често се използва за характеристика на типа екосистема. Например горска, езерна, речна, тревна. Трофичната структура обикновено се изгражда графична във вид на екологична пирамида. В основата е първо трофично ниво, а следващите формират етажите и върха. Екологичните пирамиди отразяват количествено отношението между биологичните компоненти в екосистемите или в отделни звена на трофичната вериа – хищник-жертва и т.н. В зависимост от използваните параметри за характеристика на тези количествени отношения, екологичните пирамиди са три основни типа:
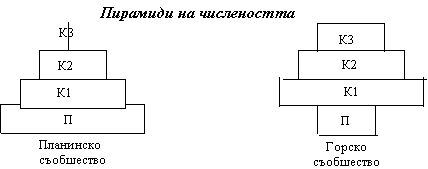
Формите на пирамидите на числеността са различни за различните съобщества в зависимост от това малки (планктонни, тревни) съобщества или големи (горски) са размерите на популациите. Числеността на популациите на различните на трофичните нива се различават обикновено с цял порядък. Това създава трудности при построяване на екологични пирамиди в един мащаб за цялото съобщество. Пирамидите на числеността общо взето не дават реално представа за функционалната роля на отделните трофични нива и на елементите, които ги формират (популации или части от популации). Пирамидите на биомасата по- добре отразяват ролята и съотношението на отделните функционални групи. В съобщества с много малки продуценти и крупни консументи те могат да бъдат обърнати, въпреки че през нивото на консументите минава по- голямо количество енергия. Например в езерата и моретата биомасата на продуцентите е много по- малка от тази на вторичните и третичните консументи. Това е свързано с бързия оборот на биомасата на планктона, даващ големи продукции на малко биомаса.
Съществува екологично правило: При използване на данните за числеността се надценява функционалнат роля на малките организми в една екосистема, а при използването на данните за биомасата – ролята на едрите организми. Числеността и биомасата във функционалната екология не са подходящи за оценката на ролята на силно различаващи се по отношението интензивност на метаболизма/размери на организмите.
Пирамидите на енергията отразяват скоростта на енергииния поток по хранителните вериги. Те не се влияят от други закономерности и винаги и винаги имат правилна форма съгласно втори закон на термодинамиката. Чрез тях могат да се съпоставят различни екосистеми и да се оцени функционалната роля на едни и същи и на различни популации в тях. Трофичната структура е фундаментално свойство на всички съобщества и екосистеми, благодарение на което то се намира в равновесие със средата и участва по определен начин в кръговрата на в-вата и утилизирането на слънчевата енергия. При хронични стресови въздействия върху екосистемите, трофичната структура може да се промени до известна степен съобразно с приспособленията на биотичните компоненти към стресовия фактор. Създава се ново равновесно състояние, което не е така устойчиво. При поява на този стресов фактор може да се стигне до деградация на съобществата.
Основни пътища при протичането на енергия през екосистемите от различен ранг могат да се отразят от така наречения унивирсален модел. Универсален е защото модел може да бъде приложен към всички живи компоненти – продуценти, консументи, популаия, трофична група. Съединени по между си подобни модели отразяват протичането на енергия по трофични вериги или енергетиката на цялатя екосистема. Популацията ня един и същи вид заема най- често различни трофични нива. Универсалният модел може да отразява енергетиката на една популация. В този случай той има вид на схема на хранителна мрежа с отбелязани видове, енергетично свързани с разглежданата популация. Възможно е чрез този модел да бъе отразявана енергетиката на отделно трофично ниво. В този случай в биомасата и на белязаните ще се включат всички популации или техни части подържани от един енергиен източник т.е. моделът ще отразява протичането на енергия по дадена хранителна верига. Елементи на универсалниа модел са:
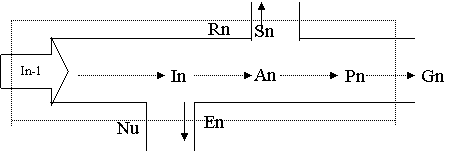
Bn – биомаса на разглежданата екосистема или неин компонент изразен в енергиини единици.
In – общо постъпване на енергия – светлина (автотрофи) органични в-ва (облигатни хетеротрофи) или и двата източника на енергия в определено съотношение (бактерии, водорасли, лишеи, безгръбначни)
In-1 – достъпна енергия
Nu – неизползвана енергия, тази част от светлината или храната, която преминава през листата или храносмилателния тракт без да се асимилира. Тази енергия е достъпна и попада в In-1
An – асимилирана енергия – общата продукция асимилирана при автотрофити или метаболизирана при хетеротрофите.
Pn – продукция – първична/вторична. Енергията, която се превръща в нова или пиринадлежаща на друга биологична система органично в-во , която е достъпна за други биологични системи.
Gn – енергията за повишаване биомасата на екоситемата
En – енергията за отделяне при жизненост чрез ексодати, екскременти или извличана от биосферата
Sn – запасна енергия – мазнини скорбяла. Енергия за по- късно използване.
Обратният поток Sn се нарича обратна връзка, чрез който се обезпечава енергията в бъдеще и се съхранява седемте елемента и функционалното състояние на биосистемата. Например може да се използва за повторно намиране на жертва.
Rn – дишане. Енергията, която се окислява и се губи под формата на топлина. Отношенията между продукция/дишане и биомаса/дишане силно варират и имат важно биологично значение. Енергията, използвана при дишане, т.е. за подържане на структурите на системата е висока за популациите с крупни организми и в съобществата с голяма биомаса. Дишането се увеличава при увеличаване температурата на средата и при стресови въздействия. Продукцията е повишена за активните популации на малките организми – бактерии, водорасли, за бързо растящите съобщества или в системи получаващи енергиини субсидии (естествени илиизкуствени)
Отношенията между величините на основните енергиини потоци в дадено трофично ниво или в различни трофични нива на хранителната верига, изразени в проценти се нарича екологична ефективност. Величинаа на енергиините потоци се изразява в cal or kcal. Най- често използваните ефективности отразяват:
![]()
КПД: q- средна калорийност на фитомасата kcal/g
Y- годишна продукция g/cm2/y
Q- достъпна слънчева радиация 380-720nm (FAR or TR) kcal/cm2/y
![]()
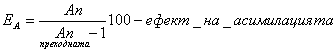
![]()
![]()
![]()
![]()
Ефективността на продуцирането е над десет пъти по- малък при топлокръвните животни в сравнение със студенокръвните, т.е. голяма част от асимилираната енергия се при дишането и подържането температурата на тялото.
Чрез изчисляване на основните типове ефективности се моделира потока на енергия. Разработени са модели за основните типове екосистеми. На базата на сравняване на моделите могат да се направят изводи за функционирането наекосистемите:
Заедно с намаляване на количеството се увеличава качеството на енергията, преминаваща по хранителните вериги. Качеството на енергията се измерва с изминатото от слънчевата енергия разстояние по трофичните вериги, т.е. калориите слънчева енергия, които трябва да се разсеят, за да се получи една калория по- висококачествена енергия. Например, за да се синтезире една калория енергия, включена във фитомасата, трябва да се изразходват няколко стотин килокалории слънчева енергия. За формирането на една килокалория енергия, включена – в биомасата на фитофагите- 1000 килокалории слънчева енергия, за една килокалория включена в биомасата на консументите около 10000 килокалории слънчева енергия, т.е. качеството на енергията за всяко следващо трофично ниво се увеличава съответно сто, хиляда и десет хиляди пъти, а количеството и намалява съответно 100, 1000 и 10000 пъти.
Термина биосфера за пръв път е използван от австрийския геолог E. Suss в трудът му “Ликът на Земята”. Като учения академик Вернадски – биосферата е една от земните обвивки, в нея са съсредоточени живите организми и резултата от тяхната дейност. Биосферата включва цялата хидросфера, тропосфера (долната част на въздуха, геосфера – ¾ от въздуха), долната част на стратосферата (включва озоновия екран), стратиосферата (слоестата горна част на литосферата- утаечната повърхност). Биосферата достига 20-25 километра в литосферата до 2-3 километра, 1-2 километра по дъното на океана, а в хидросферата 11 километра.
Биосферата не еднородна – на места е съсредоточено по- голямо количество живо в-во и продукти от обмяната. Тази част от биосферата, в която е съсредоточена по- компактна част от живото в-во е биострома (Лавренис) (слой на живата епигенема, фитогеосфера, витасфера). Биостромата се формира там, където се допират трите геосфери. Три типа:

Сухоземната биостома заема пространство от няколко сантиметра до няколко десетки метра, рядко до 150 метра над земната повърхност и няколоко 10-ки метра (10-30); максимум 1700 метра – нефтени находища под земната повърхност. Биостромата е на дълбочина 350- 400 метра под водата. Биостромата се подделя на отделни функционални единици- биосистемите. Биостромата има огромно значение за биосферата- това е най- активната и част. Там се фиксира слънчева енергия, извършва се биологичен кръговрат и се отделят необходимите за живота газове в трите геосфери. В резултат на функционирането на биостромата коренно са се изменили и се изменят съществуващите геосфери. Изменя се и газовият състав на атмосферата- съдържал е: N,NH3, CO2, CH4, водни пари. Отсъствал е кислород. Сега атмосферата съдържа: N, O2, CO2, Oзонов слой
Сумарната биомаса на организмите на Земята е 2,4.1012 тона, като около 99,87% е съсредоточена на сушата. Биосферата е възникнала преди около 3,4- 4,5.106 г. и е в непрекъснато развитие.еволюцията и протича под влиянието на алогенни (външни) фактори- геологични и климатични промени и аутогенни (вътрешни), обусловени от активността на живите компоненти. Еволюцията е преминала през следните етапи и периоди:
І Период на биогенеза
ІІ Ноогенеза
Това е период на разумно регулиране на взаимоотношението човек/природа. Основна движеща кила на еволюцията е разумът. Постепенното превръщане на биосферата в ноосфера е закономерен етап в развитието й. С появата на човека според Вернадски се формира антропосфера, социосфера и техносфера, които са земните компоненти на преобразуваната от човека биосфера. Ноосферата е най- висшият стадий в еволюцията на биосферата. Обхваща земната биосфера и част от космическото пространство. Въз основа на развитието на науката е техниката човек ще може да формира и проектира природо- техническа среда. Поставят се въпросе как да се избегнат необратимите последствията от антропогенната дейност, т.е. да се предотврати разпадането на биосферата и човека да дочака окончателната еволюция в ноосферата. Тъй като не е възможно да се спре развитието на средата, социалния и научно- техническия прогрес на човешката общност, то следва и така да се управлява процесите на взаимодействие между човека и биосферата, че да са взаимоизгодни- чрез постигане на балансираност между положителните и отрицателните обратни връзки между обществото и средата. С възникване на ноогенезата е свързано и възникването на науката ноотеника- наука за взаимоуправление на взаимодействието общество- природа. Главна задача е “изправяне” на нарушенията в биосферата в резултата на човешката дейност и предотвратяване на такива въздействия в бъдеще.
Атмосферата (газовата обвивка на Земята) е с маса 5,9.1015t и слоиста структура. Състои се от няколко сфери: тропосфера, стратосфера, мезосфера, термосфера (йоносфера), екзосфера. Между тези сфери са разположени и преходни сфери- паузи. В отделните сфери се изменя количествено и качествено състава на въздуха и температурата.
Над 4/5 от въздуха е съсредоточен в тропосферата (най- долния). Дебелината и е 10-12 километра в средните, 16-18 в екватора и 7-10 киламетра в северните ширини. Тропосферата се характеризира с неустойчиви температура, налягане, влажност; голямя подвижност- вертикално и хоризонтално движение във връзка с неравномерното нагряване на Земната повърхност. В резултат на йонизация молекулите на въздуха в стратосферата на височина 24-40 киламетра се формира озоновият слой. Атмосферата е надеждна защита от късовълновата радиация и метеоритен дъжд. Ако няма атмосфера на всеки км2 от Земята на 3-4 дена ще пада по един метеорит. Без атмосфера температурните амплитуди ще бъдат 200оС. Въздухът е необходим и за нормалното протичане на нормалните жизнени процеси при повечето организнми. Човек използва за едно денонощие повече от 500 литра кислород, пропскайки над 10000 литра въздух. За това време той приема около два килограма храна и вода. В процеса на еволюция повечето живи организми са изработили различни приспособления срещу токсичните в-ва, попадащи в храносмилателната система. Такива приспособления срещу отровните газове без цвят, аромати и вкус (с технологичен произход) няма. Промишлените замърсители на атмосферата са 2 групи: първични и вторични.
Първичните непосредствено постъпват в атмосферата от различни източници, а вторичните се образуват при фотохимичното окисление и други процеси на първичните. Вторичните могат да бъдат и по- токсични от първичните. Съществуват много и разнообразни класификации на замърсителите.
Организираното промишлено изхвърляне е чрез газопроводи, тръби и комини.
Неорганизираното изхвърляне постпва в атмосферата като ненасочени газови потоци.
ІV. Според локализацията на замърсителите: от основното производство, от спомагателното.
V. Според температурата на замърсителите: нагрети и студени
VІ. Според степента на очистване:
VІІ. Според характера си:
Химични –SO2, СО, FeO
Източниците на замърсители се делят на две основни групи: естествени (природни) и изкуствени (антропогенни)
Природните биват:
Извънземни- космически прах, при изгаряне на метеоритите 2-5.106 t/y
Естествената замърсяване включва природната прах, която е постоянна съставна част на земната атмосфера. Състои се от твърди частици с радиус 10-3 – 10-4 сантиметра и кондензирани ядра с радиус 5.10-6 сантиметра. В природната прах се включва и аеропланктон (полен, спори, плесен, микроорганизми, бактерии, продукти от гниенето и разлагането на органиката). Атмосферният въздух на океаните включва кристалчета соли на Mn, Na, и други, образуващи се при изхвърляне на капчици. Атмосферен прах- значение за химията. е формиране на валежите, поглъща пряката слънчева радиация или я разсейва, следователно има защитен ефект. В резултат на биологичното разлагане се отделят големи количества въглеводороди, амоняк, въглеродни окиси, поподащи в атмосферата. Естествените замърсителе общо взето не застрашават функционирането на екосистемите, биоценозите и биосферата. Тревожни са кратковременните локални последствия (изригване на вулкани).
Изкуствените (антропогенните) биват:
Всеки от тези източници отделя специфични замърсители. Постъпващите в масови количества замърсители са малко на брой. Транспортът и промишлеността са основни източници на замърсяване, тъй като при непълното изгаряне на топливото се отделят токсични замърсители. Автомобилните газове съдържат около 200 различни в-ва – канцерогенни въглеводороди и продукти от непълното им окисление и оловен тетраетил. Всеки бензинов двигател замърсява 25000 километра пробег консумирайки 4350 килограма кислород, а изхвърля 325 килограма СО2, 530 кг. СО, 93 кг. въглеводороди и около 27 кг. азотни окиси. На всеки автомобил се падат около 10 килограма “гумен прах” (Газовете, отделяни от ТЕЦ-овете съдържат S и N- окиси, прах, соли, метали. Средно се отделят до един тон SO2) 10 тона изгорели въглища от промишлените предприятия. Металургичните предприятия отделят прах, SO2, функционални съединения и метали. Установено е, че при получаването на един тон чугун се отделят около 5 килограма прах, 27 кг. СО2, 0.5 кг. Мn, As & Pb-ни съединения, Р, Hg, и редки метали.
Нефтодобиващите и преработващите предприятия- въглеводороди, SO2 и задушливи газове- ацетон, толуол, изопрен.
Химична промишленост- прах, SO2, H2S, CS2, Cl2, Pb, азотни окиси, железни съединения.
Изпитването на атомни оръжия, АЕЦ, недостатъчно ефективното обезвреждане на атомни отпадъци води до повишаване нивото на радиоактивността
Своеобразна форма на замърсител е шумът – слабите шумове (по- ниски от 20 dB) успокояват. Над 90-120 dB увреждат слуха и нервната система. Топлинното замърсяване е най- силно изразено над големите градове и промишлените зони- различен от регионалния климат.
Последствия от замърсяване на атмосферата. Най- голямо участие имат въглеродните окиси, съединенията на сярата, азота, въглеводородите и прахта. В земната атмосфера ежегодно се изхвърлят повече от 200 милиона тона въглеродни окиси, 20 милиарда тона въглероден диоксид, 150 милиона тона SO2, 53 милиона тона азотни окиси, 250 милиона тона прах, 120 милиона тона пепел, 50 милиона тона въглеводороди. Глобално последствие от научно-техническата революция е насищането на атмосферата с въглеродни оксиди.
Промените на състава и замърсяване на атмосферата е свързано с нарушение на енергииния баланс на Земята, нарушаване на нормалното функциониране на екосистемите и влошаване здравето на хората. Основни фактори, водещи до изменение на енергииния баланс са:
Албедото се променя чрез култивиране на определен тип растителност, унищожаването на климаксните съобщества, при хидро мелиоративни мероприятия (напояване/засушаване) и запрашаване. Хигиенния стандарт допуска допуска обща запрашеност от около тон и половина на хектар, но в определени райони може да достигне до 60 тона на хектар. Повишената запрашеност води до повишено количество на отразената слънчева радиация – намалява температурата на въздуха. Частиците прах попадащи на повърхността на ледниците поглъщат слънчевата светлина и способстват за разтапянето им. Основна роля за променената прозрачност на атмосферата има и повишаването на въглеродния диоксид средно с 5-10.109 тона на година. За последните 10 години количеството му се е повишило с повече от 10%. Той пропуска късовълновите лъчи и поглъща отразените от земята инфрачервени лъчи, това води до образуването на парниковия ефект и непосредствено влияе върху топлинния баланс.
Прозрачността зависи и от съдържаниета на водни пари – антропогенният ефект е повишаването им. Повишаването на водните пари води до намаляване защитната роля на озоновия екран, а съдържанието на водни пари в тропосферата води до промени в климата.
Разсейваната по време на горенето топлина оказва влияние върху топлинния баланс. Ежегодно се отделят 14,2.106 KJ топлина при изгаряне на топливо. Температурата на въздуха над големите градове е с 1-2 градуса по- висока във връзка с отделянето на техногенна топлина. Изменението на топлинния баланс може да доведе до затопляне или застудяване на атмосферата. Затоплянето на атмосферата с 1-2 градуса ще предизвика разтопяване на ледените шапки в Гренландия и Антарктика, това ще доведе до заливане и тектонически последици. Застудяването би довело до ледников период.
Количеството на озона в атмосферата е 2.10-6 % от обема на атмосферата, но озона има важна биологична роля. Окислението му от някои в-ва, продукти от горенето на самолетни и ракетни горива ще доведе до увеличаване радиацията на Земята. Тези дози ще бъдат увеличени с разрушаването на озона.
Изчислено е, че 3% от наличните запаси кислород биха се използвали за изгаряне на всички запаси от изкопаемо гориво. Темповете на използване на кислород изпреварват отделянето му. Една лека кола за 1000-1500 км изгаря необходимото годишно количество за човек.
Същественото намаляване на кислородните запаси оказва замърсяване на световния океан с Hg, ДДТ, пестициди и други унищожаващи продукти. Отделеният кислород от фотосинтезата на планктона е 30-70% от кислорода в атмосферата. При ускорените темпове на техническия прогрес, кислородът ще намалее с една трета за около 200 години. Човешкият организъм е чувствителен при намаляването му с 1%.
Поддържането на нивото на кислорода в атмосферата е важно не само за биологичните организми, но и запазване газовия баланс и стабилността на атмосферата. Под действието на слънчевата радиация в атмосферата се извършват реакции, в които участват кислород, азот, въглеродни и азотни окиси, при което кислорода може да встъпи в реакции с въглеводородите и да се получат вторични замърсяващи в-ва – алдехиди, формалдехиди – фотохимичен смог.
Един от съществените екологични проблеми е замърсяването на атмосферата с серни съединения. Според някои по- замърсено е северното полукълбо, където 90% от сярата е с антропогенен произход. SO2 оказво вредно действие върху продуцентите. Постъпвайки в листата угнетява клетките и предизвиква хлорози и те изсъхват. SO2 и други серни съединения имат имат дразнещо действие върху дихателните пътища и слизестите обвивки на очите. Продължителното им действие в малки концентрации води до появата на хроничен бронхит, ларингит, гастрит, рак и на други болести. Газобразният SO2 в атмосферата се окислява до SO3 от озона или от водородния прекис. Процесът се контролира от следи на метали Mn. Друг начин е образуването на SO42- и H2SO4. В сравнение с SO2, SO42- имат по- силно изразен биологичен ефект при равни концентрации. В зависимост от влажността на атмосферата SO2 може да съществува от няколко часа до няколко дни. Отношението SO2/SO42- намалява с височината, на малка височина преобладава SO2. Строителството на високите комини е свързано с преноса и разсейването на SO2 във връхните слоеве на тропосферата, това води до самоочистване на атмосферата. Това обаче увеличава времето на престой на SO2 в тропосферата и вероятността му за превръщане в SO42- и H2SO4 и падане на кисели валежи.
Според някои автори от 1955 година киселиността на дъждовната вода се е покачила над 40 пъти. Отбелязани са случаи на валежи с рН по- малко от две над промишлени райони – руши сградите. Киселинните дъждове оказват силно изразен отрицателено влияние върху стабилността и функционирането на земните и водните екосистеми. Установено е, че при постоянното им въздействие се задържа растежа, намалява се продуктивността на растителните видове на селскоктопанските култури. За съхраняване на равновесието в биотопа във водоемите е предложено варуване на 2-3 години, но това е много скъпо.
При SO42- в атмосферата се образуват мъгли и се намалява прозрачността на въздуха.
(NH4)2SO4 и NH4HSO4- - праховидни частици с размери 0.1 –1 микрона, които са силно хигроскопични. Те задържат влагата и разсейват светлината. Те оказват отрицателно влияние върху човешкото здраве. Световните запаси от сяра са почти изтощени. Разработване на методи за улавяне на SO2 и превръщането му в сяра и сулфати. Най- много SO2 се отделя от топлоенергията, черната и цветната металургия, химическата и нефтохимическата промишленост и транспорта.
Естествен начин на очистване на атмосферата от замърсители е способността и на самоочистване чрез разсейване и утаяване на замърсителите или чрез неутрализирането им от природни съединения, които реагират с тях. Природните способности за самопречистване са по- малки в сравнение с обема и скоростта на натрупване на промишлени битови и транспортни замрсители. Във връзка с това широко се прилагат различни локални и глобални мерки за очистване.
Праховидните частици могат да имат различна форма и размери в зависимост от използваните суровини. Поради голямата си сумарна повърхност те имат многа по- голяма химична и биологична активност от изходните материали. За механизмите и устройствата е от значение диаметърът на частиците – диаметър на кръг, чиято скорост на утаяване и плътност са равни на тези при частиците. Диапазона на дисперсност се определя от най- малкия и най- голямия размер на частиците в прахта. От свойствата на прахта, параметъра на газовия поток и необходимата степен на очистване зависи типът на прахоуловителят. Те могат да бъдат:
a) Сухи - гравитационни, инерционни, центробежни
b)
c)
a)
b)
Върху процесите на очистване чрез използване на прахоуловители влияние оказва влагата. При 20% съдържание е възможна кондензация и образуване на утайка и запушване на филтрите. Изборът на прахоуловител зависи и от температурата на отделяните газове. Например тъканни филтри се използват при температури до 35 градуса. Обикновено мокрите филтри са подходящи за очистването на газовете от запалителни и отровни примеси. За улавяне на прах с неорганични в-ва соли и други, Подходящи са електрически и механични мокри. За очистване на въздуха от прахови частици с размери по- малки от 5 микрона се използват електрофилтри, тъкнни и мокри. За очистване от прах с органична компонента се използват механични прахоуловители. Разнообразните методи за очистване от химични замърсители са обединени в две групи:
Некаталитични методи - извеждат се вредните примеси чрез кондензация или чрез поглъщане от течни или твърди в-ва. Те се разделят на: абсорбционни, хемосорбционни и адсорбционни.
Методите за химическо очистване се подбират на базата на химико-технологични процеси.
Хемосорбционните методи широко се използват за очистване на газове от CO, CO2, азотни окиси, SO2, H2S. Отделяните химически замърсители реагират с течни хемосорбенти – разтвори на минерални и органични в-ва, суспензии и т.н. Например очистването на газовете от SO2 се извършва с хемосорбенти варено мляко, Na2SO3, NaOH, MgO + MgSO3, ZnO, Na3PO4.
Адсорбционните методи се използват за очистване на газове с малка концентрация. Газовете се поглъщат и задържат от твърди порести в-ва – физична адсорбция, химична адсорбция. Адсорбцията протича в три фази:
Регенерирането на адсорбентите се извършва чрез десорбция с пари, температура. Степента на очистване е висока, но се губи допълнителна енергия за охлаждане и неколкократно връщане на газовете.
Недостатъци: могат да се пречистват само сухи, незапрашени газове, които да преминават през адсорбента с малка скорост. Очистването се извършва в адсорбенти, които могат да бъдат с неподвижни, с движещи се или с неподвижни или псевдо нагорещен слой от сорбента. С периодично или непериодично действие. За адсорбция на СО2 се използва активен въглен, калциев карбонат, дуламит, активен магнезиев окис.
Каталитичните методи са на основата на химичните превръщания на токсичните в нетоксични компоненти на повърхността на катализатора. На подобно очистване се подлагат газове не съдържащи прах и катализаторни отрови.
Преминаване към централизирано отопление в населените места и оборудване на топлоцентролите с
Газификация, електрификация и т.н.
Въпреки извършваните дейности па опазване и очистване на атмосферата, все оше са на лице съществени екологични проблеми – киселинни дъждове, э малко биоразнообразие и влошаване здравния статус на населението и т.н.
Световните запаси от вода са 1,9.1011км3 – течна (солена и прясна), твърда и газообразна (пресни). Запасите на земята се разпределят сътветно: около 97,2% в световния океан, около 0,64% на континента. За промишлеността и бита се използва само прясна вода. Най- големи запаси от прясна вода има в ледниците – 28,5.106 км3 и подземни питейни води 8. 106 км3 – не се използват.водите за плзване са в нищожни количества. Потреблението на вода в промишлеността и в селското стопанство е в огромни размери. Според специалисти безвъзвратното водопотребление е около 1% от повърхностното, което е повече от 150 км3, а общото потребление на прясна вода е 3103 км3/у. В селското стопанство около ¾ от използваната вода се губи безвъзвратно. За един тон пшеница, ориз, памук се изразходват около 1500, 7000, 10000 тона вода за вегетационния период. В промишлеността водата е важна суровина с различно потребление – разтвор, охлаждане, нагряване, очистване на разтвори и газови смеси. За добиване на един тон продукция се изразхадва вода в кубични метри. За стомана и каучук – 15/20, сярна киселина – 20/25, коприна – 300/400, синтетични влакна – 500, Cu – 500, пластмаси 500/1000, синтетичен каучук – 2000/3000. За работата на един ТЕЦ с мощност 300000 kW са необходими повече от 300 кубични метра.
Три милиона човека денонощна изразходват два милиона кубични метра, а годишно три милиона кубични метра.
Високите изисквания към качеството на водата водят до създаването на сложни технологични процеси за водоподържане и водоподготовка.
Качеството на водата, която се използва за производствени и други цели се установява във всеки конкретен случай в зависимост от предназначението и изискванията на технологичния процес. Качеството на водата е съвкупност от химични, физиологични, бактериологични, биологични показатели, обуславящи пригодността на водата за ползване на вода.
Следователно прясната вода е една от най- важните суровини на Земята, не може да се замени недостига от прясна вода на Земята – около 1/3 от населението. В някои страни има импорт, а в други експорт на вода. Основни причини за недостига са:
Намаляването на дебита на реките е свързано с изсичането на горите, разрушаване на естествените тревни и ливадни съобщества, пресушаване на блата и т.н. Глобалното използване на тези мероприятия е довело до увеличаване повърхностния оток – стичащата се към морето прясна вода и намалява количеството вода подхранващи подпочвените води.
За борба с намаляване на речния дебит се:
Голяма загуба – при ползване. За предотвратяване филтруването на водата през напоителните канали – ципи.
Замърсяването на водоемите с промишлени и битови води също аяожаяо водите на замърсените реки и езера са непригодни за битови нужди. Замърсяването на водоемите се характеризира с:
Замърсяването на водите може да е естествено, например с биологично активни в-ва, отделяни при цъфтежа им, продукти на ерозията и изкуствено (анропогенно).
Замърсителите се делят по характер на механични, химични и физични. Основни източници на замърсяване на природните води са:
преработващата промишленост.
При технологичните процеси се образуват отточни води.
Най- силно замърсяват повърхностните води целулозно- хартиените заводи, химическите предприятия, нефтопреработващите заводи, текстилните, металургията транспортните средства. Смята се, че във водоемите постъпват повече от 500000 различни в-ва. Основните замърсители на водата са нефта, детергентите, пестицидите, изкуствените торове и други. Замърсяването с нефт и неговите продукти е проблем на вътрешните водоеми и световния океан. В световния океан се изхвърлят 12/15 милиона тона нефт годишно, което е един проценто т транспортниа нефт. Общата замърсена с нефт площ е около 1/5 от общата акватория на световния океан. Същността на отрицателния ефект от нефтеното замърсяване е образуването на тънк слой на водната повърхност, който нарушава газобмена, възпроизводството, проникването на светлина и променя албедото. Смята се, че всеки тон нефт покрива 12 квадратги километра от водната повърхност и замърсява до един милион тона морско вода. Океана има огромна роля в газообмена. Значителна част от първичната продукция е резултат от планктона. Замърсяването с нефт може да предизвика глобални последствия върху климата и водата. Ежегодно загиват големи количества планктон и животни. Половинат риба в замърсените води не се използва за храна, заради придобития мирис и вкус. Плаващите нефтени острови се носят от теченията към бреговете и замърсяват плажовете.
Попадналите във водата полициклични ароматни въглеводороди нямат канцерогенно действие. Сред замърсителите много опасни са детергентите. Те образуват слой пяна със значителна дебелина и могат да възпрепятстват достъпа на кислород. Установено е, че един милиграм на литър предизвиква масова гибел на планктон, три милиграма на литър предизвиква масова смърт на дафнии, пет – понижава рибните ресурси. Замърсените с други замърсители води плюс детергенти трудно се очистват.
Пестициди, мъртва органика, минерални торове. Замърсяването с пестициди води до натрупванито им в планктона, така се предава по хранителната верига и при останалитетрофични нива. Те предизвикват различни функционални нарушения в отделни органи на различни организми. Особено токсични са органо хлорнити съединения те трудно се подлагат на биологично окисление.
Мъртвата органика – неконсумирана храна, урина, тор и други често попада във водните басейни иводи до еутрофикация. В отточните води, носещи органика се съдържат голямо количество биогенни елементи N u P. В резултат масово се размножава фитопланктон, синьозелени водорасли, кафеви водорасли и висши водни растения – цъфтеж. Това води до силно увеличаване на общата маса на органиката, намаляване на количеството кислород и преобладаване на анаеробни процеси и непълно разлагане на органиката. Тези процеси се наричат еутрофикация. Тя е свързана с промяна във видовия състав на водните биоценози, поради промяна в режимите, с увеличаване на биологичната продуктивност. В резултат на натрупването на биологични елементи, в резултат на антропогенни или естествени фактори. Еутрофикация може да настъпи и при измиване на азотните и фосфорните торове от културните площи. Битовите отпадни води съдържат различни органични и неорганични токсични и нетоксични в-ва, патогенни микроорганизми и други. Те са причина за редица инфекциозни заболявания и епидемии. При замърсяване с тежки метали те активно се натрупват по хранителната верига, а крайно звено е човека. Наблюдават се масови отравяния с Hg, Cd, и други при консумиране на риба от замърсени водоеми. При замърсяване на водите с радиоактивни в-ва се наблюдава биологичното им акумулиране по хранителната верига. Източници на такива замърсители са заводите, пречистващи уранови руди, АЕЦ, контейнерите с радиоактивни отпадъци, провежданите опити с атомни и водородни бомби. Изхвърлянето на контейнери с радиоактивни в-ва във водните басейни е голяма опасност за биотата в тях, тъй като е по- бързо разпадането на контейнерите от полуразпада на в-вата (Cs- 3 милиона години). В подобни басейни се наблюдава радиоактивност на планктонни организми 1000 пъти повече от тази на водата, а в зоомасата на някои риби, крайни звена в хранителната верига може да е 50 пъти по- голяма концентрацията. Животинската и растителната биомаса дълго време могат да запазят радиоактивността си. В резултат планктонът в чистите води може да е радиоактивен. “Заразени” рибни ята в резултат на миграцията си могат да попаднат в чисти води и да разпространат “заразата”.
Особено замърсяване е топлинното замърсяване – изхвърляне на топла вода. Това води до промяна на топлинния режим, режима на аерация и промени във видовия състав. Замърсяването на хидросферата като цяло от екологична гледна точка, е много по- опасно от замърсяването на атмосферата поради:
Естественото самопречистване на водата се извършва от микроорганизми, бактерии, гъби, водорасли. При смесване на замърсители с повече маса чиста вода се извършва естествена минерализация на органични в-ва и унищожаване на патогенните микроорганизми. Ниските температури забавят самопречистването. Основен принцип на самопречистването е многократното смесване на замърсени и чисти води. Така се самопречистват морските води до големите градове. Отпадъчните води се изхвърлят далеч от бреговете и на голяма дълбочина. Те се смесват с голямо количество чиста вода и така се извършва самопречистване. При голямо замърсяване не става да се прилгат съответните методи и средства за очистване. За предотвратяване на замърсяването на световния океан с нефт е сключена международна конвенция – извличането на нефт трябва да е на повече от 50 метра от бреговете. В много страни е забранено изхвърлянето на отпадъчни води в морето. Пристанищата и водните превозни средства се оборудват с пречиствателни приспособления.
Предотвратяването на загуба на нефт и нефтени продукти при товарителните работе и транспорта е обезопасено чрез оборудване с нефтоулавящи приспособления.
Борбата със замърсявването от промишлени отпадни води е свързана с изграждане на пречиствателни съоръжения, усъвършенстване на технологията и производството и изграждане на съоръжения и инсталации за извличане на ценните в-ва и повторнот използване на водата.
Методите за очистване на водата са механични, химични, физикохимични и биохимични.
Механичните методи се отнасят към група методи за предварително пречистване. Химичните и физикохимичните методи се използват отделно или в съчетание с механични и биохимични. Най- добър ефект на очистване се постига чрез комбинация на няколко метода. Методите се делят на:
Методите за очистване и конструктивно оформяне се избират съобразно факторите:
Ефективността на различните методи за очистване на отточните води е: при използване на механични методи 50-70%, химични 80-90%, физикохимични 90-95% и биохимични 85-95%. Преди подаване на отточнато вода в съоръженията тя постъпва в усреднители. Отпадъчните води, които се очистват с тези методи се подлагат на термично обезвреждане или се събират в резервуари.
Механични методи – за отделяне на твърди примеси с решетки, сита, мрежи, масло и нефтоуловители. По- малките частици чрез утаяване и филтруване. За интензифициране на тези процеси се прибавят коагуланти или се въздейства с центробежни сили, хидроциклони и други.
Химичните методи се прилагат за отделяне на разтворими примеси. Използват се различни реагенти, които встъпват в химични реакции с примесите. В резултат се получават малкоразтворими в-ва или слаботоксични в-ва, отделящи се като утайка. Най- чести реагенти са: Cl2, O2, O3. Някои в-ва се неутрализират с варено мляко. Пречистването може да се извърши по електричен път – електролизьори.
Физикохимичните методи се прилагат за отделяне на суспендирани и емулгирани примеси и на разтворени органични и неорганични в-ва. Коагулация, флотация, йонен обмен, адсорбция, екстракция, обратна осмоза, ултра филтрация, кристализиране, дестилация и други.
Биохимичните методи са основни за очистване на водите от органични замърсители, които се окисляват от микроорганизми. Биохимичното очистване може да протече в естествени условия върху специално подготвени участъци – поливни полета. Изгражда се прежа от главни и разпределителни канали, по които се разливат отпадъчни води. На база филтруването им през почвения слой те се очистват. Почвен слой с мощнос 80 сантиметра ги очиства достатъчно. Могат да се използват и изкуствени водоеми с дълбочина 0,5-1 метър. Обикновено стъпаловидно се разполагат 4-5 водоема през, които последователно преминава замърсената вода. Биохимичното очистване в изкуствени условия се извършва в биофилтри, аеротанкове или биобасейни. След очистването водата дезинфекцира с течен хлор или хлорна вар.
За отделяне на токсичните газове се извършва десорбция с инертен компонент при нагряване или добавяне на реагенти. Очистването се счита за ефективно, ако се провежда до необходимата пределно допустима концентрация. Колкото е по- голяма степента на замърсяване и по- голяма ефективността на очистване, толкова по- големи са разходите. Намаляване на разходите за очистване и увеличаване на икономическата ефективност на производството може да се постигне при:
Видът не е прота сума от индивиди, а многообразие от групировки. Видът е диференциран- сложен по състав. Представлява система с вътрешна структура и е съставен от структурни елементи. Вътревидовите групировки на всеки вид микроорганизъм, растение или животно са най- различни, но основната групировка, чрез която видът се утвърждава в околната среда, на нивото на която се приспособляват към постоянно променящите се условия и която функционира като единно цяло е популацията (populus – гр. народ, население). В такъв широк смисъл с изразена терминологична неопределеност се използва и в биоогията. За пръв път се използва от Йохансен 1903г. Той не признава влиянието на външната среда за формирането на белези и предлага популацията като противопологен термин на чистата линия (съвкупност от еднакви по наследствени белези индивиди), популация (група от генетично разновидни индивиди). Следователно всеки индивид от популацията самостоятелно реагира на средата и популацията е прост аритметичен сбор от индивидите на даден вид. Днес това понятие е с друго значение.
Популационният живот на организмите е централен живот на проучаване в екологията. Единно схващане за същността на популацията няма. Някои учени на преден план поставят проблемите на наследствеността, еволюцията; систематиците и палеонтолозите поставят на преден план генетичните особености. Други учени оставят на преден план териториалните белези – привързаност и начин на използване на териториалното значение на биологичните сигналти полета при използване на територията. Според други учени на преден план е функционалното използване на популациите– механизмите поддържащи тяхната цялост, хомеостаза единство със средата. Фитоеколозите акцентират на връзката на всяка популация с другите компоненти на биоценозата, което намира отражение в понятието ценопопулация. Според това понятие популацията е съвкупност от индивидите на един вид, живеещи на територия границите на която обикновено съвпадат с границите на биоченозата.
Популацията е естествено възникнала или под влияние на човека съвкупност от индивиди на даден вид, обединени от обща територия и известно генетично родство и отнасящи се към една или няколко екосистеми. Индивидите на популациите се характеризират с еднотипни външни признаци, сходни адаптации, единна специфична реакция на въздействие на факторите на средата, характеризират се със специфичен тип на динамика на числеността си, със своя териториална и демографска структура, с общи биологични сигнали, полета които осигуряват популационния хомеостазис, самостоятелно и съществено развитие в продължение на дълго време, съпроводено със съответно взаимодействие със средата на местообитание.
Популациите се разглеждат не само като вътревидови групировки, възникнали исторически в резултат на действието на естествения отбор, но икато групировки, които могат да имат антропогенен произход. Популациите могат да се образуват например от интродукцията на определен брой индивиди в дадено място, където са успели да се приспособят, размножат и разсеят.
Популацията е централен въпрос в екологията на животните и в по- малка степен в екологията на растенията. При съпоставяне на закономерностите на популационния живот на растенията и на животните изпъква тяхното сродство, а в някои отношения и пълна еднаквокст, което позволява много от тези закономерности да се използват като общи екологични. Въпреки това популациите на растенията и животните имат специфични особености. Дълбоки различия има например между отделните популации само от групата на растенията или от групата на животните. Например различни са популациите на прикрепените и на подвижните животни. Степента на подвижност оказва огромно въздействие. Големи различия се наблюдават и при популации, различаващи се по нивото на развитие на нервната система. Степента на изменчивост на популациите на растенията и на животните е различна. В зооекологията последните десет години е направено повече от колкото във фитоекологията. Структурата и функционирането на животинските популации са несравнимо по- сложни. Работе се много повече с зоологичен материал и най- вече с безгръбначни животни, а от гръбначните се работи с дребни – гризачи. Птиците са най- добре изучени.
Популациите са предмет на изучаване не само в екологията, а оше и в генетиката, систематиката, еволюционното учение и в редица други биологични и нибиологични науки. В генетиката – популационна генетика.
Видове популации.
Географска – тя е най- голяма по- заемана територия. Най- голямата териториална групировкана вида след подвида. Съставен е от една група индивиди на един вид, които населяват територии с еднообразни условия, но разнообразни в биоценологично отношение. Достатъчно ясно са ограничени и са относително изолирани. Размерите на географската популация зависят от биологията на вида и от условията на територията – обширни географски пространства изолирани с географсти прегради. Parus major (голям синигер) заема обширни ареали, но е представен само от пет географски популации. Географските популации се различават една от друга по отношението си към климатичните фактори, срокове на размногаване, брой поколения за година, морфологични и физиологични белези, поведение, периодичност на миграциите. Могат да са наследствено предадени.
Екологична – тези популации са от по- нисък ранг. Наричат се още биотопични, местни, ценопопулации. Състоят се от организми от един и същи вид обитаващи един и същи биотоп, привързани към конкретна биоценоза. Например катерицата (Scilurus vulgaris) обитава различни типове гора – дъбови, елови, борови популации. Тези популации са слабо изолирани за разлика от геогпафските и между тях често се осъществява генетичен обмен. Те са по- малко постоянни по състав.
Елементарна
При разглеждането на популацианната структура трябва да се има предви, че видовете реално съществуват чрез своите екологични популации и не езадължително да имат географски и елементарни популации. Броят на популациите на един вид може да е различен – от един до хиляди. Колкото ареалът на вида е по- обширен, толкова по- разнобразни са условията и следователно толкова повече са популациите.
Формирането на многo популации може да се наблюдава и на сравнително ограничено пространство. Например в планинските местности, където условията на малка територия, могат да бъдат твърде разнообразни и да съществува физична изолация на една популация от друга. В подобна обстановка възникват не само екологичните популации, но може да протече формиране на видове и родове. Броят на екологичните популации на даден вид не се определя само от големината на ареала и разнообразието на условията, които той предоставя. Популационната структура на вида зависи в много по- голяма степен от неговите биологични особености и подвижността на индивидите. Обикновено по- слабо подвижните видове образуват повече и по- малки пространствени групировки, а по- подвижните – по-малко, но по- големи пространствени групировки. Затова на една и съща територия при едни видове се образува цяла редица от популационни групи, а при други нито една. Такива големи групи от индивиди на даден вид, които населяват непрекъснато огромни територии и не се разпределят на по- малки териториални комплекси се наричат супер популации (супра популации). Например в еднородна площ с благоприятна структура на почвата Lumbricus terrestris образува единна популация върху обширен район. Ако в пределите на супрапопулациите могат да се доловят обособени групи те се наричат субпопулации.
Популациите могат да заемат площ с размери на големи части от континети или на цели континенти. Такива са популациите на северната лисица. Те също така могат да бъдат разположени и на няколко квадратни метра – много земноводни, влечуги, охлюви. Териториалните граници на популациите не са статично явление. Те могат да се изменят във връзка с колебанията на числеността – насекоми, гризачи, за които е характерно появяването им в огромни числености – каламитети и масово напускане на пределите на постониия ареал. В годините на депресии, измиране и изчезване. При процъфтяване на вида микропопулациите могат да се превърнат в екологични популации и дори в супра популации, а след това да претърпят регресия до микропопулации. Броят на индивидите, от които са изградени популациите е видово специфичен, но е есилно зависим от условията на средата. При тревистите вастения, много насекоми броят може да достигне до стотици хиляди. Популациите на едрите дървесни видове, едрите животни са по- малочислени и включват (при едрите хищници) до няколко десетки.
Популациите се разделят също по способността им за възпроизводство и времето за съществуване. Делят се на:
Основни свойства на популациите.
Генетична характеристика – всяка популация има своя генетична характеристика. Едно от основните свойства на всяка популация е генетичното родство на съставящите я индивиди. В една популация не могат да бъдат обединени, които са генетически изолирани. Популациине са взаимнокръстосващо се единство, но в природата има немалко животни и растения, които се размножават чрез самооплождане, партеногенетично или по безполов начин (делене, пъпкуване, спорообразуване) и не могат да се определят като взаимно кръстосващо се единство. Такива популации са системи от клонове или чисти линии, които използват средата, затова са относително мономорфни. Самооплождането, партенегенезисът и различните форми на неполово размножаване не са винаги първичен начин на размножаване. В повечето случаи са вторично явление. Например при самоопрашващите се растения периодично се извършва кръстосано опрашване. За покритосеменните растения, които се размножават вегетативно е задължително и половото размножаване. Партеногенезисът при много безгръбначни има периодично редуване с кръстосаното размножаване. Задължителната агамност е характерна за някои бактерии и синьо-зелени водорасли, но не е абсолютна. При тях няма полов процес, но се предполага, че се пренася генетична информация чрез вируси. Следователно такива популации се отличават от панмиктичните само по интензивността на наследствените изменения. Това се отразява на структурата им, но не е принципно тяхно отличие. Не трябва да се счита, че всяка група индивиди, обединени чрез панмиксия са една популация. В повечето случаи между съседни популации се осъществява обмен на индивиди. Благодарание на разселването на новородените (приплодите) при животните на неголеми разстояния, непрекъснато се извършва смесване на генофонда на различни популации. Смесването на генофонда определя тяхната генетична близост, но различията между тях се запазват поради фенотипичните им особености. Фенотипичните различия на организмите не се намират в съответствие с техните генотипични различия. Характерни фенотипове се формират под влиянията на конкретни условия. Тази закономерност се изразява в наличието на морфологични и физиологични различия между групи индивиди, живеещи при не еднакви условия. Различията засягат: скоростта на нарастване и развитие, размери, тегло, плодовитост, оцветяване и т.н. Затова и в такива случаи, когато генетичните общности на две съседни групи индивиди на даден вид не предизвикват съмнение, фенотипичните им особености са индикатои за различията в условията на съществуване на сравняваните видове, следователно не могат да се смятат за една популация индивиди, които не се различават генетически, но се отличават по своя фенотип.
Механизми за изграждане на териториална структура
Обикновено борбата е само система от ритуални пози и завършва като правило с победа на собственика. Пришълеца се изгонва, дори когато е по- силния. В поведението на животните, които са на своя територия преобладавт рефлексите на активна отбрана, а тези които са на чужда територия - ориентировъчни рефлекси. Обикновено собственика веднага се нахвърля върху пришълеца и го преследва до границите на своята територия. Ако при преследването се увлече и попадне на неговата територия, ролите им се сменят. Обикновено животните рядко попадат в конфликти, т.е. избягват попадането в чужда територия. Обикновено се използва маркировка, тя е много разнообразна. При много птици има акустична маркировка – показват заетостта на територията чрез песен и се задържа там, където е най- вероягтно навлизането на съсед. По границите на индивидуалния си участък мечките и дивите котки правят дълбоки драскотини по корите на дърветата, които редовно възстановяват. По- височината, на която са разположени драскотините се съди за размера на животното и за силата на живитното. Много често се използва олфакторно маркиране (чрез урина или изпражнения). Например маркиране чрез урина използват семействата на котките и кучетата. По разположението и специфичната миризма животните получават разнобразна информация за този, който ги е оставил – възраст, физиологично състояние, пол, полова активност и т.н. Използването на това олфакторно средство за информация през размножителниа период рязко се увеличава. Аналогична роля играят екскрементите. При някои животни за маркиране и информация служат секрети от специални жлези наречени телегрони или феромон. Това са секрети на анални, гръбни, коремни, околоочни идруги жлези.
При някои видове се отбранява целия индивидуален, семеен или групов участък, при други само част от него – около “биологичния център” – мястото на скривалището, гнездото. Част от териториалните участъци могат да си припокриват, там инстинктът за агресия изчезва – територия за общо ползване. Много животни имат индивидуален участък през целия си живот – дребни мишевидни гризачи. Повечето обаче имат териториален участък само през определен период от жизнения си цикъл – по време на размножаване и др. В постоянните семейни групи териториалните отношения между членовете могат да бъдат много сложни. Например тигрите са полигамни, семейството е от един мъжки и 2-3 женски и млади индивиди. Размножаването на териториалния участък зависи от числеността на жертвите. Мъжките постоянно обхождат територията и я маркират чрез екскременти или чрез урина. Участъците на младите индивиди се припокрива отчасти с тези на женските. Някои части се посещават само от мъжки. Всеки член си има любима територия за лов. Младите напускат територията на 3-5 години.
При животни със скитащ начин на живот няма индивидуални участъци. Те имат силно развита ориентация за положението на съседните. Например при стадата от копитни се подържа определено средно разстояние между тях, така че да неси пречат.
Пространствената структура при животните е възможна благодарение на две основни свойства:
Придвижването може да е пасивно и активно. Пасивно е при почти всички растения и някои неподвижни прикрепени животни. Пасивните движения се осъществяват чрез вятъра, водата, живите организми.
Общуването се осъществява по различен начин – най- древно средство е хим. чувств. Характерна е за всички видове оргонизми. За растенията, микроорганизми и някои нисши животни е единствено средство за общуване. Химическата чувствителност се усложнява с появата на обонятелни и вкусови органи.
За подържане на пространствената структура се използват и оптични сигнали. Сигнално значение има формата на тялото, оцветяването, жестове, мимики. Съществува език на цветовете, на жестовете, на формите. Всички звукове, които съпътстват движенията, изяждането на храната и други жизнени актове и техните производни от специални звукови органи са система за общуване, която е по- добре развите при по- висшите групи животни членестоноги, риби, земноводни, птици и бозайници. Акустичните параметри на гласа и звука при индивиди от един и същи вид са съгласувани. При някои птици са установени диалекти – видови и популационни. Звуковете издавани от индивиди на различни популации не са идентични и служат за общуване в рамките на популацията. Прилепите и делфините използват ехолокация. Електрична сигнализация при някои риби.
Пространствената структура е различна при различните видове. Възможно е един и същи вид да използва различни системи на пространствена организация. Един вид използва различна пространствена сруктура през различните сезони или в различни части на ареала си. В една част от ареала в един сезон са възможни възрастови и полови различия на пространствената структура. Например гора от един вид – в началото индивидите се разпределят групово, а после в резултат на конкуренцията за светлина гъстота е по- малка и дърветата са на еднакво разстояние. Водният плъх през зимата в северните части на ареала си води единичен начин на живот, а в южните групов. В групи прекарват зимата и някои насекоми (калинки), много риби, земноводни, птици, бозайници, влечуги, които през топлите месеци водят единичен начин на живот. Някои птици през топлите месеци водят единичен начин на живот, а малките като се излюпят – групов. При дивите свине мъжките водят единичен, а женските с малките групов. Някои птици некрофаги – C. Corax, врана образуват рехави ята със слаба връзка между индивидите. Водят семеен начин на живот но отделните индивиди се следят във въздуха и се струпват там, където един индивид намери храна.
Биологично значение.
В зимните струпвания на мирните риби се намалява нивото на обмяната на в-вата. Златните рибки, отлеждани единично, употребяват повече кислород, отколкото ако са няколко.
При овца извън стадото пулсът и дишането се учестяват (повече кислород), при вида на стадото се нормализира.
Снижаването нивото на обмяната при струпвания спомага изразходването на резервни в-ва – единично зимуване приели са много по- ниско ниво на обмяна от това в група, това води до повишаване загубата на енергия, изтощаване, смърт. Същото е и при земноводни, влечуги и бозайници.
Груповият начин на живот при много птици и бозайници способства за по- успешно намиране на храна, по- добра защита от неприятели. Вълците ловуват по- успешно обединени в глутници. Пеликаните гонят рибата групово към плитчините.
Много струпвания са свързани с размножаването. Колониалните патици не могат да се размножават ако колонията силно намалее по численост. За африканските слонове стадата трябва да са не повече от 25 индивида. При северния елен най- продуктивни са стадата от 300-400 индивида.
Груповият ефект не трябва да се смесва с т.нар. масов ефект, който се получава при преуплътняване и е с отрицателен ефект за популацията. За всеки вид е свойствен и специфичен минимален размер на популациите, под който те не могат да съществуват. Това се нарича принцип на минималния размер. При повечето популации смъртността е минимална при малки размери на популацията, а при други при промеждутъчен размер. Тогава е вредна пренаселеността и недонаселеността. Това се нарича принцип на О ш.
Възрастова структура - съотношението на съставящите популацията възрастови групи индивиди. Това е много важна характеристика на популацията, защото определя репродуктивните и възможности в момента и показва какво може да се очаква в бъдеще. Прието е данните за за разпределение на възрастовите групи да се представят във вид на възрастови пирамиди. В тях броят на индивидите в различни възрасти или техният дял в проценти от индивидите на цялата популация се означава с ширината на хоризонтално ориентирани и последователно подредени във вертикална плоскост фигури. Отдолу се намират най- младите индивиди, а най- отгоре най- старите.
Различават се три вида възрастови пирамиди – А, В, С.
А
В
С
При разделнополовите видове за женските и мъжките се правят отделни пирамиди. За интерпретация на данните за възрастовата структура се отчита и продължителността на различните възрастови стадии. Различават се три екологичне възрасти:
Продължителността им е различна при различните видове. При различни условия в един вид също е различна.
При съвременния човек тези възрасти са почти еднакви (по1/3). За много растения и животни е характерен дълъг предрепродуктивен период. При някои животни (насекоми) е прекалено дълъг, репродуктивният е къс, а пострепродуктивнят период може да липсва. Например еднодневките – предрепродуктивният период е 1-2 години, репродуктивния – няколко дни до една седмица, а пострепродуктивен период няма. Пострепродуктивният отсъства при растенията с еднократно плодоношение, включително и едногодишнште и при някои с многократно плодоношение. При такива видове с различна продължителност на екологичната структура, при стабилна възрастова структура отношенето между отделните възрастови групи няма да е еднакво.
Особености на възрастовата структура при растителни популации
Популация, която в даден момент е представена само от семена и от млади растения се нарича инвазионна. Популация, която няма млади индивиди, а има само сенилни и субсенилни, или от цъфтящи, но не образуващи семена индивиди се нарича регресивна. Тя не може да се самоподдържа и зависи от внесените семена от вън. Популация, която включва индивиди от всички или от почти всички възрастови групи е нормална. Нормална популация, която има индивиди от всички възрастови групи е пълночисленна, а ако има от почти всички – непълночисленна
Особености на възрастовата структура на популации на живатни
Такъв вид, който се размножава един път в живота и популацията му се състои от няколко генерации е майският бръмбар. Имагото снася яйцата през пролетта и възрастните загиват. Ежегодно това се повтаря. От яйце до имаго развитието продължава 4 години следователно едновременно в популациите присъстват 4 генерации. Всяка година една генерация завършва своя жизнен цикъл и се появява нова. Възрастовите групи в такива популации са ясно разграничени. Едновременното съществуване на няколко генерации при видовете с еднократно размножаване маже да се обуславя също от разтеглено във времето снасяне на яйца, нарастване и полово съзряване на индивидите. Това става в резултат на наследствена нееднородност на индивидите в популацията и под влияние на различия в микроклиматичните и други условия. Например домашната муха зимува като ларва и като имаго. През лятото се развиват 4-5 генерации. Едновременно се срещат представители от няколко поколения, но винаги едно от тях (най- скоро създалото се) преобладава.
Много по сложна е възрастовата структура в популации с видове, размножаващи се многократно. Възможни са две крайни ситуации:
Биологично значение на възрастовата структура.
Биологичното значение на възрастовите различия в устройството, функциите и поведението е огромно, тъй като така се усложняват и обогатяват връзките между организмите и средата и се откриват възможности за по- пълното и използване.
Възрастовата структура не е статично явление. Тя непрекъснато се променя – поради промяна на външната среда или в самата популация. Въпреки, че е динамично явление, за всяка популация е характерна известна нормална/стабилна възрастова структура, към коята всяка популация се стреми.
Полова структура – съотоншението между половите групи. При повечето видове полът се определя в момоента на оплождането – генотипно. Съотоншението на зиготите по пол е 1:1, но не следва такова съотношение за популацията въобще. Мъжките и женските индивиди са със значителни физиологични, екологични и етологични различия и за това не е еднаква смъртност, преживяемост на индивидите от различните полове е широко разпространено явление. Нееднаквата смъртност е налице не само при онтогенетичното развитие, но може да се прояви още през ембрионалното развитие. При ондатрата в много райони сред новородените малки са 1.5 повече. Женските новородени преобладават при вълците, златките и други хищници. При повечето животни е обратно сред приплодите преобладаван мъжките. Раждането на повече женски има компенсаторен аооао. Женските са по- жизнеспособни, а смъртността при мъжките е по- висока. По- голямата жизнеспособност на женските за сега не е е изяснена. В основата и стоят физиологични, биохимични и други особености. Това явление е с важно значение, тъй като възпроизводството на популациите в голяма степен зависи от женските индивиди. Мъжките индивиди са по- подвижната част на популацията, по- често са извън убежищата, по- често стават жертва на хищници и неблагоприятни условия. Подвижността и смъртността на възрастните мъжки е голяма, най- много през размножителния период и затова в края му често преобладават възрастните женски – при повечето гризачи, фазани, зеленоглава патица. Вероятно в много случаи развитието на полигамията исторически е обусловена от по- голямата смъртност на мъжките индидивиди. При женските индивиди смъртността е най- голяма през зимата, тъй като основните усилия за отглеждането на малките се падат на тях и през зимата са по- изтощени. Така дялът на женските при някои видове прилепи след зимния сън пада до 20%. В края на размножителния период във възрастната част на популцията преобладават женските, а през зимата съотношението се изравнява. Половата структура се определя не само генетично, но зависи и от условията. При кафявата горска мравка температурата определя пола. От яйцата, снесени при температури под 20оС се развиват мъжки, а от яйцата снесени при по- високи температури – женски. Мускулатурата на семеприемника на матката, където се задържа семенната течност след оплождане се активира само при високи температури и яйцето се опложда – развива се женски индивид. От неоплодените се развиват мъжките.
При някои видове полът по начало се определя от факторите на средата. Това са видове, чиито зиготи са генетично еднополови – еднодомни растения, някои червеи, охлюви, ракообразни и насекоми. Например от големите, с много хранителни запаси клубени на Arisema japonica израстват екземпляри с женски цветове, а от малките – с мъжки. Водната бълха (Daphnia) при нормални условия се размножава партеногенетично, а при по- ниски или по- високи температури се появяват и мъжки индивиди. При листните въшки появата на мъжки индивиди е съпроводеа с понижаване температурата на въздуха и продължителността на светлия ден.
Динамика на половата структура.
Биологично значение.
Наличието на полови групи увеличава полиморфизма но популациите. Освен по физиологични признаци двата пола се различават и по екологични особености, които се изразяват в особеностите на хранене. При много видове мъжките и женските през определен период са и териториално разделени това дава възможност за по- пълноценно използване на средата.
Индивидите, които образуват една популация са в известна степен качествено не еднородна – в морфологични или поведенчески особености. Тя е основа за възникването между контактуващите индивиди на отношения на съподчинение, които се характеризират с доминирането на едни индивиди и подчинението на други – и при растенията и при животните.
При растенията и при прикрепените животни отношенията се регулират чрез промени на физичните особености и химизма на вътре популационната среда. Много по- сложна е йерархичнат структура на популациите на подвижните животни. Тази структура се нарича етологична, тъй като в основата и лежат особености на поведението. Поведението на животните в границата на популацията се определя от това дали за вида е характерен единичен или групов начин на живот. Системата на доминиране е различна при различните видове, но може да се сведе до три типа:
![]()
Всеки индивид е подчинен на предходния и доминира на следващия. Последните са най- безправни. Животните от последния ранг проявяват признаци на покорност пред останалите – хранят се последни, заемат най- неудобните места, женските не достигат за тях.
Подобни отношения възникват при спрегнатите кучета. Могат да се образуват и по- сложни отношения – паралелни редове на съподчинение – при женски и мъжки (при някои маймуни – павиани, мъжките доминират над женските, а те над младите.)
Йерархичната структура е извънредно динамича. Съставът на групите се променя – прорастват приплодия, някои индивиди загиват, пришълци, емигранти и т.н. Тази динамика има положително зночеие, защото структурата на популацията се привежда в съответствие с условията на средата.
Биологично значение.
Съгласуваното поведение има значение за защита от хищници, защита на малките, миграция при неблагоприятни условия. Например при гладуване загиват първо най- слабите, подчинените индивиди. Така се постига подържане на по- висока жизненост.До женските имат достъп най- силните и жизнени мъжки и така се запазват най- добрите качества.
Всяка йерархична група е свързана с определено физиологично състояние на организма. При бозайниците подчинените животни са с хипертрофия на кората на надбъбречната жлеза. Най- силно е изразено при индивидите на най- ниските стъпала, при доминантните индивиди няма хипертрофия. Стероидните хормони на надбъбречната жлеза оказват голямо влияние на процесите на обмяна на в-вата, които лежат в основата на промените на поведението.
Ролята, която дадена популация играе в биоценозите и екосистемите зависи не само от видовата принадлежност, но и от това колко са и на каква площ са разположени, т.е. от числеността и пътността.
Численост на популацията е общият брой на индивидите в популацията. Тя е непостоянна величина и зависи от съотношението раждаемост и смъртност, от емиграцията и имиграцията.
Плътността на популацията е броят на индивидите или биомасата на единица площ или в единица обем. За почвените и водните индивиди – на единица обем. Плътността на популацията изразена чрез броя на индивидите/биомасата е показател е напълно достатъчен за оценка на състоянието на популацията. За оценка на ролята на популациите в биоценозите и екосистемите броя на индивиди/биомасата не са най- надеждният критерии. Когато размерите и интензивността на метаболизма на индивидите на сравняваните популации не са различни или много малко се различават, броят на индивидите е напълно задовобяваща мярка, но когато не са еднакви, е на лице правилото: ”данните за числеността довеждат до преувеличаване значението на малките организми, а данните за биомасата – преувеличаване значението на едрите организми.”
Следователно тези критерии (численост и биомаса) са непригодни за сравняване функционалната роля за популации различаващи се по размери и интензивност на метаболизма на съставящите ги индивиди. Най- надеждният показател за сравняване на един компонент на дадена биоценоза с друг е потокът на енергия, т.е. количеството на фиксираната енергия на единица площ за единица време в даденото трофично ниво.
Плътността е изменчива величина, но не безкрайно. Съществуват определени най- ниски и най- високи стойности за плътността за всяка популация, зад които не може да съществува. Горната граница на плътността се определя от мястото на популацията в трофичната верига. По принцип (с много изключения) колкото нивото е по- високо, толкова възможната горна граница плътността е по- ниска и обратно. Долната граница не е ясно изразена и се определя трудно.
Видове.
Основни групи методи за определяне на плътноста
Метод на извадките – отчитане или претегляне на организми от дадена популация от известен брой извадки – обемни и пространствени и чрез интерполация се изчислява плътността на популацията в цялата екосистема. Най- често използвани методи. Голямо внимание се обръща на пространствената структура на изследваната популация, от която много зависи големината и броят на извадките. Стандартните статистически методи се базират на случайното разпределение. При равномерно разпределение броят на извадките е най- малък, а при групово най- голям. Колкото числеността е по- голяма, толкова с по- малко на брой и по- малки по обем или площ проби може да се достигт.не до задоволителни резултати. При по- дребните организми броят и обемът на пробите може да е по- малък, а при по- едрите по- голям.
Методи на улавяне и белязане със следващо повторно улавяне. При по- подвижни животни. Ако една популация се състои от N индивида, от които (а) индивиди са уловени и маркирани и след това пуснати, след известно време се прави повторно улавяне – (b) индивиди. От тях (с) са маркирани и се получава:
a/N = c/b следователно N = a.b/c
Прилагането на този метод е възможно когато популацията е стабилна, няма емиграции и имиграции, раждането и смъртността са равни и когато първото улавяне не променя поведението на популацията. Не се прилага по време на размножителния период. Големи грешки се допускат при ниска плътност на популациите (N,a,b,c са ниски).
Косвени методи Само при животни. За плътността се съди по видими белизи от жизнената дейност на животните – дупки, къртичини, хралупи, обилие на екскременти, брой убити животни, по остатъците в храната на хищниците числеността на жертвите по погадки.
Корелацията между получените резултати и реалната плътност обикновено е незначителна, следователно приложението на тези методи е незначително – за приблизителна оценка на популациите.
Промятата на числеността на популацията се определя от две противоположни явления: раждаемост и смъртност, а също и миграции и емиграции.
Раждаемостта е способността на популацията да се самовъзпроизвежда. Терминът раждаемост се използва за означаване честотата на появата на нови индивиди в популацията независимо дали се раждат, излюпват или се образуват чрез делене или пъпкуване.
Има два вида раждаемост:
Двата вида са от своя страна:
Максималната раждаемост на всяка популация е максимална величина. Да се знае тази величина е наложително, защото е критерий за сравняване на реализуемата раждаемост, т.е. критерий по който се разбира в каква степен дадена популация, в дадена способност реализира своята максимална размножителна способност. Понеже е относително постоянна величина, може да се използва за прогнозиране големината на популацията. Например очевидно е, че обикновеното коприварче със своите 4-6 яйца, не може да се размножи в необикновено големи количества за определено време. Брашненият червей с 570 яйца на женска, ако условията са брагоприятни, ще възникне популационен взрив. Максималната раждаемост не се определя лесна на практика. За практически цели може да се установи приблизително, чрез експериментални методи. Например най- висока средна продукция (на люцерна), получени при серия експерименти извеждани при най- благоприятни условия. Ако се премести неголяма група чехълчета в нова неблагоприятна среда, то достигнатата скорост на размножаване може да служи ката максималана раждаемост. Добрият специалист може да се възползва от този метод и при полеви условия, доколкото в природата също се създават такива екологични ситуации, при които действието на лимитиращите фактори е временно силно понижено.
Фактори влияещи върху раждаемоста.
Върху раждаемостта влияние оказва и друг фактор – пространствена структура. Липсата на индивидуален участък при видовете, водещи единично- семеен начин на живот е свързан с откъс от размножаването.
Реакцията на популацията към факторите има приспособителен характер. Чрез промяна на раждаемостта се постига оптимизиране на числеността към състоянието на средата, но пълна оптимизация не се постига, защото тя се управлява и от други механизми. Раждаемостта може да е положителна или 0, но никога не и отрицателна.
Раждаемостта е понятие, което се отнася към цялата популация, а не към отделен индивид. За мярка на раждаемостта се приема средната, а не най- малката или най- голямата индивидуална репродуктивна способност.
Основно свойство на всяка популация, което в известна степен се явява антитеза на раждаемостта е смъртността. С нея се отбелязва честотата на отпадане на индивиди от популацията поради смърт. Смъртността бива:
Смъртността бива:
Причините за смъртността могат да бъдат най- различни. Много от тях за конкретни индивиди могат да имат случаен характер, но при популацията смъртността има закономерен характер. Нейната величина закономерно се изменя в зависимост от:
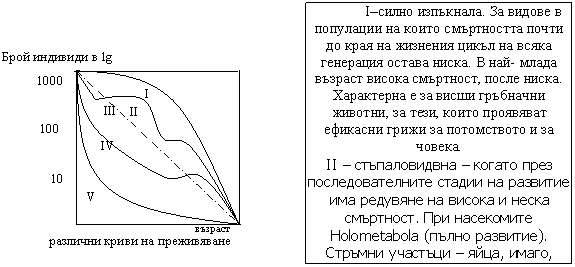
--->предкакавиден стадий. Полегати участъци – ларви и какавиди – по- малка смъртност.
ІV – за много видове птици, дребни гризачи, зайци смъртността е малко по- голяма при младите, а при възрастните е по- ниска и по- постоянна.
V – при висока смъртност в младите стадии. Характерна е сравнително устойчива малка смъртност на възрастните индивиди. При много от растителните и животинските видове.
Числеността е променлива величина. По начина на промените във времето, колебанията на числеността могат да се разделят на:
Най- интересни са промените на плътността несвързани нито със сезонни, нито с годишни колебания на условията, но сясно изразена периодичност през няколко години. Например в северните зони при бозайниците, птиците, насекомите, рибите и растенията. Класически пример са 9-10 годишните цикли на заека и риса в Канада. По- кратки 3-4 годишни цикли са характерни за много северни мишевидни гризачи и техните хищници. Характерни са и за много насекоми. Например боровата педомерка, гъсениците на която се хранят с листата на бора, средно на 10 години се появява в големи числености. Известни са годишни многочислени колебания на някои скакалци – Logusta migratoria – на 40 години се появява в голяма численост и мигрира. Налице е популационен взрив.
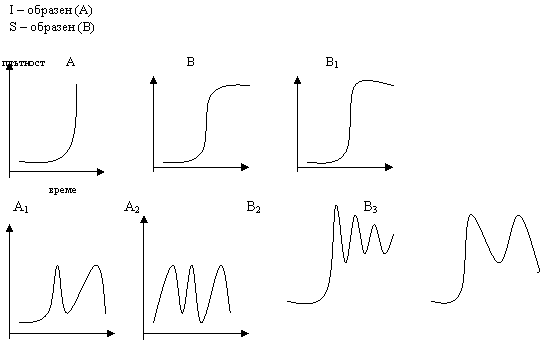 При I –
образния тип
При I –
образния тип
S – образен тип. Увеличаването на популацията отначало става бавно, после по- бързо, но от дадедн момент отново става бавно, накрая нарастването се прекратява. В крайна сметка се достига и се подържа повече или по- малко устойчива равновесие. За разлика от I – образната крива, при която лимитиращити фактори въздействат едва в края на нарастване на популацията, пре S – образната крива действието на лимитиращите фактори се увеличава постепенно с увеличаване плътността на популацията. Такъв тип (S) има при много микроорганизми, растения и животни. Идеална S – образна крива имат немного организми. Въздействието на отрицателни фактори увеличава линейно при повишавени плътността само при популации на организми с прост жизнен цикъл (дрожди, развиват се в ограничено пространство). В популации с по- голяма организация на живот, при растения и животни със сложен жизнен цикъл и дълги периоди на индивидуално развитие характерът на колебанията на числеността се модифицира в една или в друга степен. В тези случаи S – образната крива е по- вдлъбната, т.е. необходим е по- дълъг период, за да се почувства ефектът на увеличената раждаемост. Освен това популацията почти винаги “прескача” над своя предел на нарастване ипреди числеността и да се установи на този предел, числеността претърпява редица колебания.
риложение на S- образните криви. Следователно отношението между увеличаването на популациите и плътностите им се мени невинаги при достигане на висока плътност се наблюдава отрицателна корелация. Само при човека тя винаги е положителна. Изключение: популациите на някои видове растения и животни при някои ситуации може да се изплъзнат от регулаторното влияние на плътността и да се получи взрив на плътността.
еории за колебанията на числеността.
І) определящо влияние за колебанията на числеността играят климатичните фактори, на биотичните фактори се отдава второстепенно значение. Застъпници на тези теории са еколози от области с много неблагоприятни условия и такива, които изследват дребни организми, за които е характерен къс жизнен цикъл – планктон, насекоми и т.н.
ІІ) числеността се управлява само от екологични фактори. Тези теории се поддържат от еколози, работещи в места с оптимални условия, с едри обекти с продължителен жизнен цикъл – птици, бозайници, дървесни видове.
ІІІ) междинна позиция – решаващ може да е биотичният или абиотичният фактор
Групират факторите на:
фактори независещи от плътността на популацията – климатични, физикохимични, някои биотични. Отричането на физикохимичните условия на средата значи да се отричат доказните фактори – температура, светлина и други влияят върху продължителността на живота, плодовитостта и други. Засушяванията, наводняванията, силните студове предизвикват значителна и избирателна плътност, която довежда в някои случаи до рязка промяна на плътността. Друг фактор е например мястото за гнездене – брой на достъпните места. Известно е у нас, че числеността на гълъба хралупар Columba .. намалява рязко поради изсичане на стари гори и липса на хралупести дървета за гнездене.
Действието на климатичните фактори може да се отрази върху популациите не веднага, а с известно закъснение. Например при благоприятно лято иглолистните гори в Северна Европа обилно плодоносят на следващата година.
Сокерицата при наличие на обилна храна през текущатата година снася през слдващата пролет средно не 3, а 4 яйца. Следователно благоприятното стечение на климатичните фактори и увеличаванито на числеността при този вид са разделени от дву годишен период.
Частично зависещи са - взаимодействията между популациите, а изцялозависещ е само един фактор – вътревидовата конкуренция. Отрицателно взаимодействие в широк смисъл е всяко взаимодействие между популациите на различни видове, което се отразява вредно върху увеличаването на числеността поне на една от тези популации. Към отрицателните взаимодействия се отнасят – влияния, възникващи не само в средствие на намаляването на ресурсите – конкуренция, но и непосредствено взаимодействие – хищничество, аразитизъм, заболяванияот различни инфекциозни агенти, влияние на различни вредни в-ва. Междувидовите химични посредници се наричат алелопатични в-ва (антагонистични аломони).
Ролята на хищниците, паразитите и болестотворните агенти като ограничаващ фактор са очевидни. Когато числеността на жертвите се увелии, плодовитостта на хищниците също нараства. Ако величината на увеличаване на хищниците напълно съответства на увеличаванто на жертвите, хищникът е в състояние да ограничи числеността на жертвата. В противен случай хищникът от даден момент може да се превърне в неефективен, т.е. популацията на жертвата се изплъзва от ограничаващото влияние на хищника. И така той се превръща във фактор не зависщ от плътността.
Степента на паразитизъм също расте до определено ниво на плътност, а след това може да започне да намалява. Разпространението на много инфекции става по- лесно и по- бързо в популации с по- голяма плътност.
Използването на знанията за факторите, зависещи от плътността имат голямо плактическо значение – биологична борба в селското и горското стопанство.
За разлика от междувидовата конкуренция (хищници и паразити), вътревидовата конкуренция изцяло зависи от плътността. Тя може да се прояви в различни проявления – като борба за храна, за жизнено пространство, като канибализъм. Вътревидовата конкуренция може да доведе до физиологични и поведенчески промени на популацията известни като шокова болест. Например при голяма численост гъсениците на една пеперуда загубват активността си, хранят се слабо, заболяват. Имагото им е също по- слабо активно, не е склонно към разсейване и отлага яйцата си в близост до мястото на имагиниране. Основавайки се на медицинска теория за стреса се натрупват доказателства, че с увеличената численост при висшите животни се увеличават надбъбречните жлези, което е симптом за нарушена нервно- ендокринно равновесие, което се отразява на поведението, репродуктивните възможности, устойчивостта към заболяване и други. Шоковата болест довежда до значително намаляване плътността на популациите.
Следователно и двата типа фактори: частично и пълно зависими и независимите от плътността фактори са свързани и постоянно си взаимодействат. В районите със сравнително устойчив и благоприятен за размножаване климат основна роля много често играят биотичните фактори, а в райони с по- неблагоприятен климат (зимен период) определяща е ролята на климатичните фактори. Колебанията на плътността на популацията зависи и от степента на сложност на екосистемата. Колкото е по- сложна (с повече компоненти), толкова по- устойчиви и независими от външни условия са популациите. Бързи и значителни колебания на числеността са свойствени за простите екосистеми – с малък брой видове и къси хранителни вериги. И най- малкото променяне на едно трофично ниво силно се тразява на следващите. Такива екосистеми са характелни за тундровите области – обратно е в тропичните области. Природните екосистеми в умерените области заемат промеждутъчно положение.
Популациите са екологични макросистеми, които не съществуват самостоятелно, а са интегрирани в макросистеми от по- висок ранг – биоценози. Синоними са асоциация и съобщества.
Биоценозата е комплекс от организирана група взаимосвързани, взаимодействащи си популации на микроорганизми, растения, животни и гъби, обитаващи съвместно пространство – територия/акватория с повече или по- малко еднородни условия.
Биоценозата е въведена 1877 година от Карл Мьобиус, той е изучавал стридни съобщества в Северно море. Установил е, че струпването от миди не е сучайно, а е сложно организиран комплекс от индивиди, управляван, оформен в хода на еволюцията под действието на естествения отбор и борбата за съществуване.
Това понятие биоценоза е изиграло роля за насочване вниманието на изледователя към взаимодействието на организмите като основа за съществуване на екологичните съобщества. Различни автори му придават различен смисъл:
Най- важните особености на биоценозата като система на организация на живота са:
Биоценозата е комплекс от популации на различни видове, които вземат участие във функционирането на една екосистема не само постоянно, но и периодично. В биоценозите са включени не само микроорганизмите, растенията и животните, постоянно обитаващи, но и тези, които прекарват само част от жизнените се цикли. Например много ларви населяват водоеми, а после са сухоземни. Много птици гнездят в гориу а се хранят в ливади и пасища – участват в няколко екосистеми едновременно. Много видове бозайници, прелетни птици. При изучаване на биоценозите те се разчленяват на отделни компоненти: микробоценози, фитоценози, зооценози. Използват се и термини с по- тясно съдържание – ентомоценози, орнитоценози и т.н. Това подразделяне на биоценозите е изкуствено, тъй като е неправилно да се отделят от единния природен комплекс, защото не могат да съществуват самостоятелно. Но това подразделяне се прави и сцел техническо удобство при различни видове изследвания и се използва широко в практиката.
По начина си на възникване биоценозите се подразделят на две групи:
Изкуствени – възникнали под влияние на човека, или са повлияни от него. Всички агроекосистеми (агробиоценози), полезащитни пояси, язовири, пресушени блата.
Между изкуствените и естествените биоценози няма редки разлики, но между тях, но между тях съществуват разлики в структурата и функционирането. Естествените биоценози обикновено са богати на видове, а изкуствените имат много по- малко видове, но са в голяма численост. В естествените биоценози количественото съотношение на индивидите на различните видовете е взаимно обусловено. В тях действат механизми регулиращи това съотношение. В естествените биоценози има динамично равновесие, поддържащо най- изгодни количествени пропорции. В изкуствените биоценози такива механизми няма и човек е поел напълно грижата да поддържа съотношението между видовете.
Всяка биоценоза има наименование. Няма точни правила за образуване на имената. Най- често имената се дават на структурна основа по името на най- важните видове – доминанти, или не едификаторните видове. На сушата на растителни видове. Имената на животни се използват по- рядко за водни биоценози (съобщество на стридни банки). В много случаи видовият състав на съобществата се променя. Например при видове с къс жизнен цикъл – планктонни съобщества и наименованието може да се даде и по друг, набиващ се в очи и устойчив биогенен или абиогенен фактор. Например съобщества на бързо течаща вода, на пясъчното дъно.
Същност на екосистемите
Всяка биоценоза има определен биохронологичен ранг – заема определена територия/акватория с еднакви или почти еднакви условия наречена биотоп. Климатичните условия в нея образуват т.нар. климатоп, а почвата едафотоп, водата – хидротоп. Системата биоценоза + биотоп се нарича екосистема. Биоценозата е органичната компонента на екосистемата, а биотопа – неорганичната.
Понятието екосистема е предложено от Тенсли 1935 г., а биогеоценоза от Сукачов 1940 година.
Двата термина са почти напълно идентични и се използват като синоними. Терминът екосистема има някои предимства.
Не всяка комбинация от организми и среда е екосистема. Съобществата от организми са свързани с ж с тесни материално- енергиини връзки. Растенията могат да съществуват само за сметка на вода, кислород, СО2, минерални соли. Хетеротрофите живеят за сметка на растенията, но се нуждаят и от кислород и вода. След като във всяко конкретно място запасите от неорганични съединения непрекъснато се използват, то те биха се изчерпили бързо ако не се възобновяваха. Те се възобновяват непрекъснато. Връщането на самите биогенни елементи в средата става от самите организми- дишане екскреции, дефекации и след смъртта им разлагане. Така съобществото от организми образува с неорганичната среда система, в която потокът от елементи получен от жизнедеятелността на организмите, показва тенденция на движение в кръг. Екосистемата е такова обединение на организмите с неорганичната среда, което се характеризира със стабилност и има ясно обособен функциониращ вътрешен кръговрат на в-вата. От количествена гледна точка екосистемата може да се счита такова обединение на оргнизми и неорганични в-ва вътрешният обмен на в-вата, екосистемният кръговрат, на което е повече от външния. Външен елемент винаги съществува, тогава всички екосистеми са открити системи. От тях се изнасят и в тях се внасят в-ва. Възможно е в някои екосистеми външният кръговрат да е по- голям от вътрешния т.е. изнасянето на в-ва да е толкова голямо, че стабилността на екосистемата да се поддържа за сметка на такова количество в-ва отвън от други екосистеми. Такива екосистеми са например реките, ручеите и други. Със значително по- пълен кръговрат на в-вата и относително по- устойчиви са горските екосистеми, ливадните и други, но нито една екосистема няма напълно затворен кръговрат. За да се поддържа кръговрат на в-вата в екосистемата, трябва да са на лице и трите функционално различни групи организми – продуценти, консументи и редуценти. По принцип кръговрата на в-вата може да се поддържа и без консументи, но такива екосистеми са изключение. Екосистемите са естествени (природни, първични) и изкуствени (вторични, антропогенни). В съвременния свят под влияние на човека природните екосистеми намаляват своя дял.
Названията на екосистемите се образува като и при биоценозите. На сушата – растителни видове, доминанти, едификати. Във водата – доминантни животински видове , може и по постояненен абиотичен фактор.
Структура
Закономерностите в отношенията и връзките на частите и структурата на биоценозите е многопланова.
Влизащите в състава на биоценозата видове и техните отношения по численост или биомаса. Има бедни и богати на видове биоценози. Бедни са биоценозите в арктическите райони и тундрата- дефицитни на топлина, в пясъчните пустини, замърсените водоеми – един или няколко фактора силно се отклоняват от оптималното за живот ниво. Не много видове могат да се приспособяват към такива условия. Бедни са и биоценозите, които често са подложени на катастрофални въздействия – ежегодно заливане, разораване, пестициди.
Там, където условията на абиотичната среда са близки до оптималните за живот биоценози са богати на видове – например влажни тропически гори, коралови рифове и други.
Видовият състав зависи и от продължителността но живот, от историята на биоценозата. Младите биоценози имат малък набор от видове в сравнение със “зрелите” съобщества. Биоценозите, създадени от човека са по- бедни от сходните природни екосистеми. Това еднообразие и беден видов състав на агроценозите човек поддържа съзнателно. И най- бедните биоценози включват няколко десетки видове от различни систематични групи.
В една пшенична агроекосистема, освен пшеница има и плевели, насекоми вредители, мишевидни гризачи, безгръбначни в почвата, патогенни гъби, хищници и други.
Почти всяка сухоземна и повечето водни биоценози включват в състава си микроорганизми, растения и животни. Съществуват и случаи, в които липсва някой от компонентите- например в пещерите или дълбоките слоеве на водите липсват растения. Съществуват биоценози само от микроорганизми – при анаеробни условия.
Видовият състав на биоценозите зависи и от разнородността на средата за обитание. Влиянието на разнообразието в условията върху броя на видовете се изразява особено ясно чрез т.нар. граничен ефект в екотоните. Границите на биоценозите рядко са резки. По такъв начин между съседните екосистеми се създава по- тясна или по- широка преходна зона със свой температурен режим, налягяне, влажност и осветяване. В тазе зона се преплитат типичните условия на съседните екосистеми. Тази зона се нарича екотон. Във всеки екотон има видове от двете гранични биоценози. Във всеки екотон има специфични за него видове, следователно екотонът е с много по- богата биоценоза. Това явление се нарича граничен ефект. Колкото повече са различията на двата биотопа, толкова по- разнообразни са условията на границата им по- силно се проявява граничният ефект. Видовото богатство е особено голямо в местата на контакти на силно различаващи се биоценози – горски и тревни, водни и сухоземни и т.н. Разнородност на средата се създава както от абиотични фактори, така и от самите организми. Всеки вид в биоценозата създава условия за живот и на други видове, които са свързани с него чрез трофични и топични взаимоотношения. Например лалугерите, заселили се в едно ниво, привличат хищници, които се хранят с тях и донасят около 50 вида паразити и стотици видове обитатели на техните дупки. Допълнителна разнообразност на средата за животните създава растителността. Колкото по- силно е развита, толкова по- разнообразни са микроклиматичните им условия и по- голям е броят на видовете животни.
Освен броят на видовете в биоценозите за характеризиране на видовата структура от значение са количествените съотношения на видовете. Във всяка биоценоза съществуват няколко (малко) видове с висока численост и множество видове с ниска численост. Най- многочислените видове, които се явяват определящи за функционирането на биоценозите като цяло са доминантните видове. Те господстват и образуват основните ядра на съобществата. Те се наричат основни видове. За да се определят доминантите на всяко съобщество е необходимо за всеки компонент на биоценозата да се определи степен на доминантност – отразява процентното участе на всеки вид от общата численост на всички видове. ж да се определи плътноста на популацията на всеки вид и се изчислява в процент какъв дял и се падат от общатата плътност.
Тъй като във всяка биоценоза числено преоблаават най- малките форми – микроорганизми, при сравнение на различни по размери видове показателят за доминиране не е в състояние да отрази истинската роля на компонентите в съобществото. За това този показател се изчислява не за съобщество, а за отделни групировки, в рамките на които разликата между размерите на индивидите може да се пренебрегне. Такива групровки се различават по различни признаци – систематични (птици, насекоми, бозайници), морфологични (еколого- морфологични), непосредствено по размери – микрофауна, мезофауна
Доминантите господстват в съобществата, но не винаги играят еднаква роля. Сред тях се отделят видове, които със своято жизнедеятелност в най- голяма степен създават среда за съобществата и без които съществуванета на други видове не е възможно. Такива доминантни видове се наричат едификатори (строител). В сухоземните екосистеми едификатори обикновено са определени видове растения. Например в степите едификатори са чиместите тревни видове. В смърчова гора- смърч, в елова гора- ела. В някои случаи едификатори са животни. Например на територия заето от мармоти, характера на ландшафта, микроклимата се определят от ровещата дейност на тези животни. Освен доминантни в биоценозата влизат и много други видове с ниска численост и редки видове. Те също са важни за живот в биоценозата. Те създават видовото богатство, увеличават разнообразието на биотопните връзки и са резерв за попълване състава на доминантните видове. Те придават устойчивост и надеждност за функционирането на биоценозите в различни условия. Колкото е по- голям резервът от такива видове, толкова по- голяма е вероятността между тях да се намерят такива видове, които до станат доминатни при промяна на средата.
Колкото е по- голямо видовото разнообразие на една биоценоза, толкова по- стабилна е тя.
Пространствената структура се определя преди всичко от растителни видове. Тя може да бъде вертикална и хоризонтална. При съвместно обитание на различни по височина растния, фитоценозата е с ясно изразен слоест строеж. Асимилиращите наземни органи и подземните им части са в няколко слоя/етажи, където използват по различен начин и променят средата. Надземните етажи при растенията се отделят по това къде е разположена основната маса от асимилиращи органи. Етажността е особено ясно изразена в области с умерен климат. Например в една широколистна гора 5-6 надземни етажа.
Най- високите етажи – Іви етаж
по- ниски дървета – ІІри етаж,
храсти – ІІІти,
високи треви – ІVти,
по- ниски треви – V,
най- ниски треви – VІ.
По- проста е етажността в иглолистните гори. При една елова гора:
Іви етаж – дървета
ІІ – храсти
ІІІ – треви
ІV – мъхов етаж
Етажът от храсти и треви може и да липсва.
Тревните съобщества – ливади, степи, савани имат по- малко и необособени етажи.
Етажите могат да бъдат и с различна дебелина: няколко метра – дървета,няколко сантиметра – мъхове. Всеки етаж е приспособен към определен комплекс от условия, но и сам участва в образуването на такъв комплекс отусловия. Например в иглолистните гори растенията от етажите на тревите и храстите се намират при по- слабо осветяване, по- изравнени температури, слаби въздушни течения, висока влажност и съдържание на кислород. Така етажите на дърветата и етажите на дърветата и храстите се намират в различна екологична обстановка. ж
Подземната етажност е свързана с различната дълбочина на разположението на корените. В горите могат да се наблюдават да 6 подземни етажа.
Растенията от всеки етаж създават специфична среда, към която са привързани различни видове микроорганизми и животни. За това всяка микробоценоза и зооценоза имат слоест строеж. Дори птиците се придържат към някой етаж. Всеки етаж в биоценозата се различава не само по височина, но и по състав на организмите и по ролята, която играе в живота на съобществото.
В биоценозата има винаги и извънетажни организми – водорасли, лишеи, лианите, животни.
Хоризонтална структура
Разчленеността на биоценозите в хоризонтално направление е известна като мозаечност и е характерна за всички биоценози. Образуваните в хоризонтално направление структурни части на биоценозата се наричат синузии. Те са групировки, които се различават по видов състав, по количественото съотношение на видовете, по продуктивността и други. Мозаечността се обуславя от различни причини: нееднородност на релефа, различни свойства на почвата, различни абиотични фактори. Стимулират и в резултат на дейността на животни или човек – разораване от животни или човека, изсушаване на гори. Мозаечността може да бъде резултат от различни природни бедствия. Когато към синузията се включи почвата, в която тя е обособена и климатът, тя се нарича парцела. Следователно за разлика от синузията, която е комплекс на органична единица, парцелата е комплекс от органични и неорганични елементи. Парцелата е една екосистема в екосистемата или микроекосистема.
Друг вид микроекосистема е консорцията. Тя е съвкупност от популации, жизнеността на които в границите на една биоценоза трофически и топически са свързани с популация на определен вид централен член на консорцията. Той се нарича детерминант, а ведовете обособени около него – консорти. Консорция може да се образува около всеки вид с някакво средообразно действие. Консорция е например борът с неговите микоризни гъби, с епифитните лишеи и мъхове, с множество населяващи го членестоноги. Полевката с нейните ектопаразити, хелминти, първаци и бактерии е също консорциум. Обикновено в ролята на централен вид са едификаторите. Тъй като всеки член на една голяяма консорция може да е център на по- малко обединение се различават консорции от І, ІІ и ІІІ порядък, следователно биоценозата е система от свързани по между си консорции и когато поради някаква причина някой член на консорцията е унищожен, особено ако е детерминант, цялата консорция се променя или разрушава. Това води до промяна на други консорции, а от там и на цялата биоценоза. Аналогични промени стават и при продукцията на нови видове. Всеки нов вид може да дойде с част от консортите си, да се включи в някоя консорция или да създаде нова.
Различните типове биоценози се характеризират с определенивзаимоотношения между екологичните групи от видове, което представлява екологичната структура на биоценозите. Биоценозите със сходна структура могат да имат различен видов сустав, така едни и същи екологични ниши могат да бъдат заемани от сходни поекология, но отдалечени в родствено отношение. Такива видове които изпълняват едни и същи функции в сходни биоценози се наричат викарииращи. Явлението еколог. викариат е широко разпространено в природата. Например една и съща екологична ниша заемат белката и златката в европейската тайга и самурът в азиатската тайга, бизоните в Сев. Америка, антилопите в саваните на Африка.
Конкретен вид за биоценозата е случайно явление, така биоценозата се формира от тези видове които се намират в конкретната среда. Но екологичната структура на биоценозите, образуваща се в определен климат и ландшафтни условия е строго закономерно явление. Например в биоценозите на различните природни зони, закономерно се изменя съотношението между фитофагите и сапрофагите. В степите, полупостините и пустините фитофагите преобладават над сапрофагите. В горските съобщества на умерения пояс обратно по-силно е представена сапрофагията.Основен тип хранене в дълбочината на океана е хищничеството, а в осветената повърхностна зона преобладават филтраторите използващи фитопланктонили със смесен тип хранене. Трофичната структура на такива биоценози е различна.
Екологичната структура е също съотношението на групи организми като хидрофити, мезофити и ксерофити при растенията и хидрофили, мезофили и ксерофили при животните. Разнообразието и численоста на представителите на една или друга екологична група характеризират биотопа в не по-малка степен отколкото точното измерване на физ. и хим. параметри на средата.Такъв подход към оценка на биоценозите, при който се използав общата характеристика на тяхната екологична, видова и пространствена структура, еколозите наричат макроскопичен - оценяваме биоценозата чрез подробно изучаване на всички детайли от екологията на всеки вид в биоценозата
Една от характерните особености на всяка биоценоза, съответно на всяка екосистема е изменяивоста – 2 типа:
Переодично повтарящи се – денонощни и сезонни промени в биоценозите. Например температурата, hv влияе върху денон. Промени в биоценозите. Едни от членовете на биоценозите са активни през деня, други през ноща (за жив. и раст. съотв.). При растенията денонощната изменчивост се проявява в течение на физиологичните процеси (цветовете на някои растения се отварят само нощем) При животните голямо значение имат денон. миграций – почвени обитатели – вертикално.
Сезонните разлики са по-съществени. Свързани са със сезонните цикли на природните условия. Смяната на условията влияе силно на съобществата. Наблюдават се периоди на цъфтеж, зимен покой, зимен сън, миграций, диапауза и т.н..
Поради разликата на тези условия през сезоните и през денон. се различават дневни и нощни стаций; сезонни стации, стации за размн. и хранене. Всички те могат да бъдат в разл. екосистеми. Стации – местообитанието на вида. Видовете които не напускат пределите на екосистемата хомохомоценни видове. Видовете с периодически миграции в друг биотоп са хетероцентни, а тези които не са привързани към определен биотоп – убиквисти.
При денон. и сезони изменения в биоценозите целоста им не се нарушава трайно. Те могат да бъдат подложени и на въздействия, които ги изменят съществено, трайно и необратимо или напълно ги разрушават. Тези изменения могат да бъдат предизвикани от нарушаването на противоречията между видовете, поради изменения в хранителния субстрат или поради изменения в абиотичния компонент на екосистемата и поради дейноста на човека. В такъв случай на мястото на дадена биоценоза се развива друга приспособена към новите условия.
1962г. Със заповед на “Главно управление на горите” се защитават дивите животни на територията на България.
1986г. Заповед на “Комитета за опазване на природната среда” – списък с ценни видове птици – за опазването им от изчезване.
1986г. Заповед на КОПС обявяват се известен брой защитени видове земноводни и влечуги.
1997г. Прието е постановление на министерския съвет и е определена тарифа за обезщетение на за причинени щети на природни обекти.
1972г. Конференция на ООН по проблемите на околната среда в Стокхолм. Приетата декларация е международен договор за екоогична защита на Земята.
Конвенция за опазване на световното културно и природно наследство. Има по- широк формат. Освен биологични формации има за цел опазване на различни исторически и културни паметници. В България се прилага от 1975г.
1973г. е открита за дописване конвенции по международна търговия със защитени видове от дивата флора и фауна. Страните членки се договарят да ограничат или спрат търговията със стоки или суровини от застрашени от изчезване видове. Списъкът на защитените видове се актуаизира на всеки три години. В България се прилага от 1991г.
1979г. Бернска конвенция – запазване на дивата европейска флора и фауна в природните местообитания. Голямо значение за запазването на биоразнообразието в Европа, но включва и неевропейските страни, в които видовете мигрират. Опазване на миграционите пътища за гнездене и размножаване, за зимуване на всички изброени видове от дивата европейска флора и фауна. В БЪлгария се прилага от май 1991г.
1979г. Бонска конвенция – за съхраняване на мигриращите видове диви животни. Страните се договарят да си сътрудничат при опазване на тези в-ва, д полагат стоги мерки за опазване и да се подготвят за подписване на специфични договори за някои видове. В България от 1979г.
1989- 1992г. Глобална стратегия за опазване на биологичното разнообразие. От експерти на цял свят с помощта на правителства, фондации и програма на ООН за опазване на околната среда. Глобалната стратегия отбелязва, че устойчивото използване на природни ресурси означава тяхното опазване и използване по начин, осигуряващ нормално съществуване на всички видове и функциониране на екосистеми.
1992г. “Среща на Земята” – подписана е конвенция за биологично разнообразие на Земята от членките на ООН. Признават се неприходните екологични, генетични, социални, икономични, научни образуват културната и естетическата стойност на биологичното разнообразие. Потвърждава суверените права на всички държави върху биологичните и ресурси. Общата воля на участниците е да създават системи от защитни територии да възтановят нарушените екосистеми. Предвижда финансови механизми и действия за въвличане на цялата общественост за опазване на биоразнообразието за бъдещите поколения. В България се прилага от 1996г.
Почвата е една от основните среди на живот и е продкт на живота. Основа за образуване на почвата е материнската скала. Тъй като скалите имат различен химичен състав това определя до голяма степен бъдещите свойства на почвата. Първите организми са нисши растения и микроорганизми. Със своята жизнена дейност те също предизвикват химични промени в скалния субстрат. След смъртта им те стават среда за живот на различни микроорганизми и ги разлагата до прости органични и неорганични съединения. Неорганичните съединения са включени в състава на скалната маса. Върху тази основа от скална маса примесена с органични в-ва и минерални елементи и заселена от микроорганизми, които продължават да минерализират органични съединения се заселват и по- висши растения и нисши животни, едноклетъчни и дребни безгръбначни. Процесът на почвообразуване продължава до достигане на динамично равновесие, при което почвата бавно променя качеството си като увелчава количеството и се засилва от все повече организми. Всяка почва се характеризира с редиц химични и физични свойства:
Физични свойства на почвата:
Тези свойства имат важно значение за организмите
Химични свойства: Химичен състав на неорганичните и органичните в-ва. В зависимост от вида на почвата минералните частици в нея са 50- 90%. Големината им варира от 1- 2 микрона до няколко сантиметра. В зависимост от размерите на частиците се различават
песачливи – 20 микрона – 0.2 nm
едрозърнести – по- големи от 0.2 nm
Органичните в-ва са размесени с минералните частици в почвата. Всяка почва се характеризира с дебелина, плътност, възможност за проникване на вода, газове, соли, светлина. Почвите имат различен набор от живи организми. Всяка почва е най- голямата в света зоологическа и ботаническа градина едновременно. Главни групи организми в почвата са: бактерии, гъби, водорасли, първаци, нематоди, насекоми и дъждовни червеи – играят голяма роля в изграждането и обработката на почвата.
Почвата е основен природен ресурс, който човек използва за свое облагодетелстване. Без нея е невъзможно съществуването му. Човешкато дейност е давала и дава дълбоки отражения върху почвата. За съжаление човек не винаги я използва най- целесъобразно и с мисъл за опазването и.
Като следствие голяма часто то почввите са или силно разорани и унищожени или са силно променени в неблагоприятна за плодородието насока. У нас ¼ от селскостопанските земи са засегнати от ерозията, коята е резултат от неправилното отношение на почвената обработка или поради изсичане на горите. Друга ¼ е застрашена от ерозия. Целите Източн Родопи ималки части от различни планинин са подложени на ерозия. България по време на средновековието е била покрита с гъсти дъбови гори. Тракийската низина и Идточните Родопи сега са оголени. През турското робство Витоша, София, склоновете на Странджа са били покрити с дъбови гори. Унищожаването на горите води до нарушаване на водния баланс на реките. Появяват се пороища и се засилва ерозията. Ветровете засилват действието си и така също засилват ерозията. Думата ерозия идва от латински – изронване. Това е процес, при които скалите или почвените частици се откъсват от първоначалното си място, пренасят се и се отлагат на ново място. Пряки причини са вятърът, водата, ледниците, земните свлачища, приплъзванията между земните пластове и други. Тя е естествена – под влияние на природни фактори. Нарича се геологична. Извършва се с изключително бавна скорост от образуването на Земята до днес. Планининте, равнините, долините се създават от водната или ветровата ерозия, действащи в течение на дълги периоди. С появата на човека започва изкуствената ерозия, която протича с много по- голяма скорост.
Друго отрицателно въздействие върху почвата е засолването. То се предизвиква от наличието на лесно разтворими соли главно на Na –NaCl, Na2SO4 и други. Наблюдава се главно в зони със сух климат, с недостатъчно атмосферно овлажняване. В такива почви поради изпаряването преобладава възходящият ток но вода, който донася в повърхностните почвени хоризонти голямо количество лесно разтворими соли. Затова непрекъснатото напояване в такива почви е вредно за повечето растения и животински организми на почвата. Засолените почви са главно два вида:
СОЛОНЧАЦИ – при тях Na често не надвишава 50% от разтворените катйони, рH<8.
СОЛОНЦИ – съдържат излишък от натрий рН до 9.
Оподзоляване – процесите протичат при хладен и влажен климат в покритите с горска растителност почви. Терминът оподзоляване е образуван от руски – под- отдолу, зола- пепел. В условията на влажен климат падат голямо количество валежи повече от 750 l/m2 годишно. Мъртвата горска постилка от листа, клонки се разлага от различни видове гъби. Проникващата надолу вода от изобилстващите валежи отмива разтворимите калциеви, калиеви и нитратни соли под зоната, в която биха били достъпни за корините – отиват до материнската скала.
Заблатяване
Върху почвата оказват влияние и някои мероприятия на съвременното земедеулие, които могат да доведат до почвена умора – силно намаляване на добивите и дори загуба на растенията при продължително безсменно отглеждане на култури. При това отглеждане се нарушава структурата и хранителният режим на почвата, натрупват се токсични в-ва на метаболизма на почвените организми.
Употреба на пестициди – около 40% от т.нар. устойчиви пестициди се задържат непроменени в песакливите и глинесто- песакливите почви поне 14 години след използването им. Употребата на органо- хлорни пестициди сериозно намалява почвената продукция – неблагоприятно за съществуването и дейността на почвата, N- фиксиращите и нитрифициращите бактерии.
За възстановяване на почвеното плодородие се предприемат следните мерки:
Сеитбообращения голямо значение за подобряване на почвеното плодородие и намаляване на ерозията. Това е редуване на културите върху една и съща почва.
Пряко унищожаване на почвите – застрояване върху обработваеми земи. Много почви се унищожават при добиване на полезни изкопаеми, при открити рудници, строителство на водохранилища, създаване на каменни кариери, в торфопреработваемите територии.
МЕРОПРИЯТИЯ ПРЕДПАЗВАЩИ ОТ ЕРОЗИЯ
Основни причини за ерозията са:
Мерките са съобразени с тези причини
Основни признаци и принципи са:
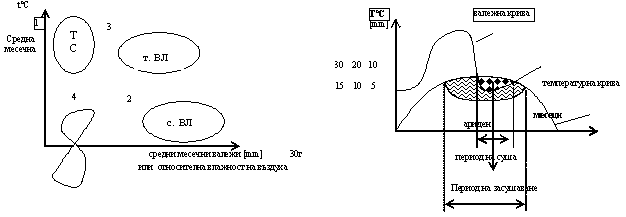
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 7797
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved