| CATEGORII DOCUMENTE |
| Alimentatie nutritie | Asistenta sociala | Cosmetica frumusete | Logopedie | Retete culinare | Sport |
FIZIOLOGIA APARATULUI DIGESTIV

FUNCTIA DE DIGESTIE SI ABSORBTIE
1. Formatiuni intestinale adaptate pentru absorbtie:
Valvulele conivente: reprezinta pliuri de forma semilunara, ocupand 1/2 sau 2/3 din lumenul intestinal. Ele apar in duodenul descendent, au densitate maxima in jejun, diminueaza spre ileonul terminal. Se gasesc in numar de aproximativ 800 si tripleaza suprafata mucoasei intestinale.
Vilozitatile intestinale: de-a lungul intestinului subtire, membrana mucoasei este acoperita vilozitatile intestinale - formatiuni in deget de manusa, cu lungimea de 0,5 - 1mm, acoperite de un singur strat de epiteliu si continand o retea de capilare si vase limfatice. Vilozitatile detin functiile de baza ale intestinului: digestia si absorbtia. Prezenta lor mareste suprafata intestinala de 30 de ori.
Microvilozitatile enterocitare: se gasesc in zona apicala a enterocitului sub forma unor prelungiri ale citoplasmei si membranei, constituind marginea in 'perie' sau platoul striat. Se gasesc in numar de 650 - 3000 pe celula, avand o lungime de 1mm si contribuie la marirea suprafetei intestinale de 70 - 600 ori. Intre microvilozitati si deasupra lor se afla glicocalix - filamente de glicoproteine generate de enterocit, implicate in digestia unor constituenti alimentari.
Celulele epiteliului mucoasei, in special de la nivelul vilozitatilor, contin enzime digestive:
- peptidaze care digera peptidele pana la aminoacizi;
- dizaharidaze: maltaza, izomaltaza, lactaza, zaharaza hidrolizeaza dizaharidele in monozaharide;
- lipaza intestinala hidrolizeaza grasimile neutre in acizi grasi si glicerol. Majoritatea acestor enzime se afla la nivelul marginii in perie a celulei epiteliale intestinale.
2. Digestia si absorbtia glucidelor
GLUCIDELE DIN ALIMENTATIE
Intr-o alimentatie obisnuita, glucidele reprezinta principala sursa de calorii. Sursa majora de glucide este amidonul vegetal, amilopectina, care are o molecula ramificata de monomeri de glucoza si o greutate moleculara mai mare de 106.
Zaharoza si lactoza sunt principalele dizaharide alimentare, iar glucoza si fructoza principalele monozaharide.
Celuloza nu poate fi hidrolizata in tractul digestiv al omului. Prezenta ei este necesara deoarece stimuleaza peristaltismul intestinal. Deoarece singura forma absorbita este cea de monozaharid, este necesar ca in procesul de digestie polizaharidele si dizaharidele sa fie hidrolizate la monozaharide.
DIGESTIA GLUCIDELOR.
Amidonul este un polimer al glucozei alcatuit din doua componente:
amiloza, in care resturile glicozil se leaga prin legaturi a-1,4 glicozidice terminale;
amilopectina, in care resturile de glicozil se leaga nu numai prin legaturi a-1,4 glicozidice, ci si prin legaturi a-1,6, conferind moleculei de amidon o structura foarte ramificata.
I. Digestia amidonului incepe in gura sub actiunea amilazei salivare (ptialina), care catalizeaza hidroliza legaturilor 1,4 glicozidice.
Actiunea amilazei salivare continua in stomac pana ce pH-ul scade la 4 prin amestecul bolului alimentar cu sucul gastric acid.
II. In duoden digestia glucidelor este reluata prin actiunea amilazei pancreatice. Ambele tipuri de amilaza isi incep actiunea de la periferia moleculei spre centru, desfacand exclusiv legaturi 1,4 glicozidice. Cum legaturile 1,6 si cele 1,4 din vecinatatea ramificatiilor nu pot fi desfacute de catre amilaza, produsii de digestie ai amidonului vor fi maltoza si fragmente oligozaharidice de dimensiuni variabile - dextrine. Dextrinele sunt hidrolizate ulterior, sub actiunea unei hidrolaze, amilo-1,6-glucozidaza, la maltoza.
Amilaza pancreatica este foarte activa. Aproximativ la 10 minute dupa ce ajunge in duoden, amidonul e transformat in maltoza, maltrioza si a-dextrine ce contin de la 5 la 9 monomeri de glucoza.
III. In continuare digestia oligozaharidelor este realizata de oligozaharidaze care se gasesc la nivelul microvililor enterocitelor..
Oligozaharidazele (dizaharidaze)
Lactaza scindeaza lactoza in glucoza si galactoza;
Zaharaza (sucraza) scindeaza zaharoza (sucroza) in glucoza si fructoza;
Maltaza a-dextrinaza) cliveaza legaturile 1,6 ale a-dextrinelor;
Glucoamilaza scindeaza malto-oligozaharidele in glucoza.
Activitatea acestor patru oligozaharidaze este mai intensa in duoden si jejunul superior si scade gradat in restul intestinluli subtire.
ABSORBTIA GLUCIDELOR
Absorbtia monozaharidelor, respectiv a glucozei, galactozei si fructozei precum si a unor pentoze implica doua mecanisme: transportul activ contragradient si transportul facilitat.
Duodenul si jejunul superior au cea mai mare capacite de absorbtie. Aceasta scade progresiv in jejunul inferior si ileon.
Glucoza si galactoza sunt transportate activ prin celula epiteliala cu microvili de o proteina transportoare numita SGLT1.
SGLT1 foloseste energia gradientului de Na+ la transportul activ al glucozei si galactozei. SGLT1 transporta 2 Na+ si o molecula de glucoza sau galactoza ce sunt competitoare la traversarea marginii in perie. Diferenta de potential a ionilor de Na este reglata de pompa de Na+/K+/ATP-dependenta prezenta in membrana bazolaterala a celulelor epiteliului intestinal.
Glucoza si galactoza parasesc enterocitele la nivelul membranei bazolaterale prin transport facilitat. Proteina transportoare responsabila pentru efluxul glucozei si galactozei este GLUT2. aceasta este prezenta de asemenea in ficat, rinichi si celulele pancreatice.
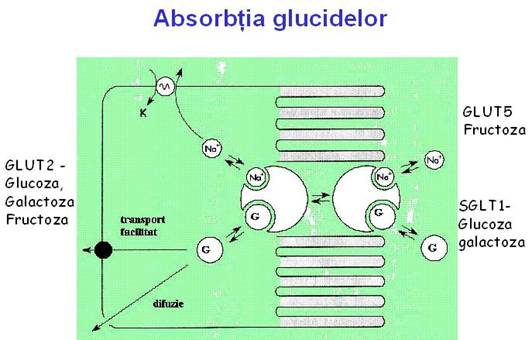
Transportul fructozei este facilitat de o proteina transportoare numita GLUT5, care este specifica pentru fructoza si nu este inhibat de glucoza, galactoza sau alte zaharuri. Fructoza traverseaza membrana bazolaterala a celulelor epiteliale intestinale cu ajutorul aceluiasi transportor GLUT2 folosit de glucoza si galactoza.
Aceste proteine trasportoare se gasesc in celulele epiteliale intestinale mature aproape de varful cililor, dar nu sunt prezente in celulele intestinale imature, in criptele Lieberkuhn.
Dupa traversarea barierei intestinale, monozaharidele sunt preluate de sangele venei porte si transportate in ficat, unde sunt depozitate sub forma de polizaharide (glicogen). Aceasta cantitate echivalenta cu aproximativ 150 g glucoza constituie o masa metabolica relativ constanta a ficatului. Atat depozitarea cat si eliberarea sunt dependente de hormoni (insulina, glucagon, adrenalina), spre deosebire de absorbtie, digestie sau reabsorbtia renala, care se realizeaza independent de actiunea hormonilor.
Monozaharidele si dizaharidele din alimente sunt complet absorbite in intestin. In schimb, aproximativ 10% din amidonul ingerat ajunge in colon, unde devine o sursa de carbonati pentru bacterii.
Malabsorbtia glucidelor are drept cauza deficienta unei oligozaharide de la nivelul microvilozitatilor enterocitare. Sindromul de malabsorbtie al lactozei este cea mai comuna tulburare de acest tip si se datoreaza absentei lactazei la nivelul duodenului si jejunului.
3. Digestia si absorbtia proteinelor
I. Digestia proteinelor incepe in stomac sub actiunea pepsinei. pepsina este secretata intr-o forma inactiva, de proenzima, este activata de acidul clorhidric si are o actiune optima la un pH cuprins intre 1,6 si 3,2. Proteinele sunt initial degradate de acidul clorhidric la acid -proteine, dupa care sunt supuse actiunii pepsinei, care cliveaza o parte din legaturile peptidice, ducand la formarea de polipeptide de diverse marimi. Actiunea enzimei este stopata in duoden si jejun datorita alcalinitatii sucului pancreatic si duodenal.
II. Pentru a-si putea exercita actiunea, enzimele pancreatice secretate sub forma inactiva trebuie activate in duoden. Activarea lor este indusa de enterokinaza intestinala (enteropeptidaza). In plus, tripsina pancreatica odata activata poate activa toate celelalte enzime proteolitice pancreatice, si are si o actiune autocatalitica de activare a tripsinogenului. Deficitul congenital de enterokinaza duce la imposibilitatea digestiei proteice si la instalarea malnutritiei proteice. Enzimele proteolitice pancreatice actioneaza la un pH alcalin si determina hidroliza proteinelor la polipeptide si aminoacizi(AA).
Tripsina si chimotripsina sunt endopeptidaze care determina hidroliza proteinelor la polipeptidaze
Elastaza este o endopeptidaza ca determina hidroliza elastinei
Carbopeptidaza este o exopeptidaza care determina hidroliza AA dela capetele carboxi si amino ale polipeptidelor, initiind eliberarea primilor aminoacizi, ca forme finale ale digestiei proteinelor
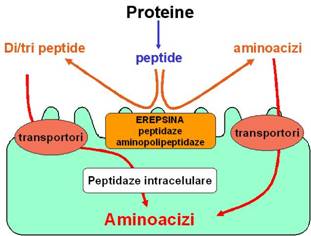
Digestia proteinelor este finalizata de peptidazele sucului intestinal, prezente la nivelul marginii in perie (microvililor) a enetrocitelor. Sub actiunea lor, peptidele sunt hidrolizate la aminoacizi, forma finala si absorbabila a digestiei proteinelor.
O parte dintre di- si tripeptidaze pot fi transportate activ in enterocite, unde sunt supuse actiunii hidrolitice a peptidazelor intracelulare.
Se poate considera ca formarea produsilor finali de digestie a proteinelor are loc la trei niveluri:
In lumenul intestinal, sub actiunea carboxipeptidazei pancreatice;
La nivelul microvililor, sub actiunea peptidazelor din sucul intestinal;
In enterocite, sub actiunea peptidazelor intracelulare.
O cantitate mica de proteine nedigerate ajung in colon, unde sunt supuse digestiei enzimatice a florei microbiene. Proteinele din materiile fecale nu sunt de origine aliimentara, ci rezulta din resturile bacteriene si celulare.
Absorbtia aminoacizilor
Absorbtia aminoacizilor din lumenul intestinal in enterocit se realizeaza prin cel putin 7 sisteme de transport diferite.
Absorbtia di- si tripeptidelor
Transportul acestora din lumen in enterocit se face printr-un sistem H+ dependent, spe deosebire de mecanismele de transport ale AA, care sunt Na+ dependente. In enterocit, prin hidroliza enzimatica, rezulta aminoacizi, care sunt transportati in sangele portal alaturi de AA absorbiti din lumenul intestinal.
Absorbtia proteinelor
Mici cantitati de proteine nedigerate pot dfi absorbite in enterocit printr-un proces de endocitoza si trecute apoi in sange prin exocitoza. Astfel se explica absorbtia anticorpilor din laptele matern. Dezavantajul acestui mecanism consta in posibilitatea aparitiei alergiei alimentare, deoarece trecerea in circulatia sanguina a proteinelor straine duce la formarea de anticorpi specifici fata de acestea.
4. Digestia si absorbtia lipidelor
DIGESTIA LIPIDELOR
Digestia trigliceridelor este realizata de lipaze.
Cea mai mare parte a digestiei trigliceridelor are loc in duoden, sub actiunea lipazei pancreatice. Lipidele emulsifiate in prealabil de catre sarurile biliare sunt hidrolizate de catre lipaza la acizi grasi liberi si monogliceride. Activitatea lipazei pancreatice se desfasoara la un pH alcalin (asigurat de HCO3).
In afara lipazelor pancreatice, exista o lipaza gastrica, mai putin importanta (la sugar), si lipaza intestinala.
Colesterolul alimentar este in cea mai mare parte esterificat. El este hidrolizat de sterol-ester hidrolaza pancreatica la colesterol liber.
Fosfolipidele alimentare sunt digerate de fosfolipaza A2 pancreatica la acizi grasi si lizo-fosfolipide.
In lumenul intestinal, produsii de digestie ai lipidelor formeaza impreuna cu sarurile biliare micele. Micelele sunt agregate cilindrice cu un continut lipidic diferit:
In zona centrala hidrofoba contin in general acizi grasi, monogliceride si colesterol.
In zona periferica contin sarurile biliare.
Rolul miceliilor este de a solubiliza lipidele si de a asigura transportul lor in enterocite.
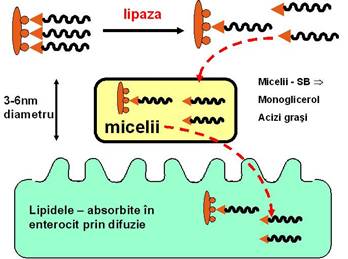
ABSORBTIA LIPIDELOR
La nivelul microvililor enterocitelor, lipidele se desfac din structura micelelor si trec in enterocit cu ajutorul unor carrier-i specifici. Sarurile biliare se desfac din structura micelelor si raman in lumenul intestinal pentru a forma noi micele. In prezenta lor se absorb 97% din lipide, pe cand in absenta lor, se atinge un procent de numai 50 - 60%.
In enterocit:
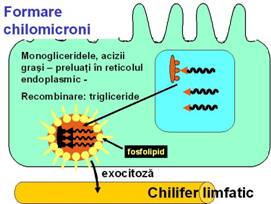
acizii grasi si monogliceridele refac rapid trigliceridele, mentinand un gradient de concentratie intre lumenul intestinal si celula favorabil absorbtiei. O parte din trigliceride sunt formate din glicerofosfatul rezultat din catabolismul glucozei.
colesterolul liber absorbit in enterocit este convertit la colesterol esterificat. Colesterolul se absoarbe lent in limfa.
Trigliceridele si cea mai mare parte a colesterolului esterificat formeaza impreuna cu fosfolipidele si niste proteine hidrosolubile specifice (apo-proteine) complexe denumite chilomicroni. Chilomicronii sunt eliberati pe la polul bazal enterocitelor prin exocitoza si trec in limfa. Datorita marii cantitati de de chilomicroni din limfaticele intestinale, acestea se mai numesc si vase chilifere, iar limfa - chil. Din limfa, chilomicronii trec in sange, uinde persista aproximativ 6 ore.
Acizii grasi cu lant scurt ca cei din lipidele din unt, sunt direct absorbiti la capilarele sanguine din vilozitati, de unde trec in sangele portal, spre deosebire de acizii grasi cu lant lung, care sunt convertiti in trigliceride in enterocit.
In lipsa enzimelor pancreatice, ca si in cazul lipsei sarurilor biliare, digestia si absorbtia lipidelor este alterata si deficitara; lipidele se elimina in scaun, determinand aspectul steatoreic al acestuia. Aparitia steatoreei se mai poate datora inhibitiai lipazei pancreatice in prezenta unui exces de secretie gastrica acida, care reduce semnificativ pH-ul intestinal (ca in cazul tumorilor secretante de gastrina).
5. Absorbtia apei, mineralelor si vitaminelor
 ABSORBTIA
APEI
ABSORBTIA
APEI
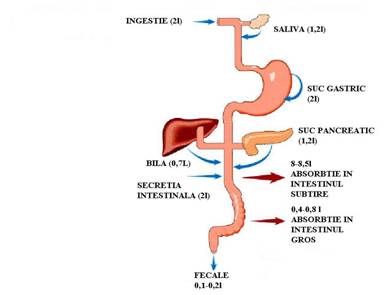
Turnoverul zilnic al apei in tractul gastrointestinal este reprezentat de aportul lichidian de aproximativ 9l/zi (2l din lichidele ingerate si 7l din secretiile digestive) si eliminarea a mai putin de 200ml/zi prin fecale. Deci, apa este absorbita in procent de 98%. Apa este transportata conform legilor osmozei, pentru a realiza izoosmolaritatea chimului cu plasma. Se realizeaza prin porii paracelulari de la nivelul jonctiunilor intercelulare.
|
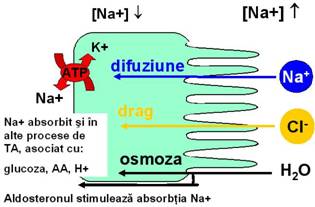
ABSORBTIA SODIULUI Sodiul trece din lumen in enterocit pe baza gradientului electrochimic, iar apoi este pompat activ in spatiile in sange prin zona bazo-laterala a enterocitului. Sodiul este absorbit si in alte procese de transport activ, asociat cu glucide, AA, ioni de hidrogen. Aldosteronul stimuleaza absorbtia intestinala a sodiului, mai ales la nivelul colonului. ABSORBTIA CLORULUI Clorul este absorbit pasiv, secundar absorbtiei sodiului. |
|
|
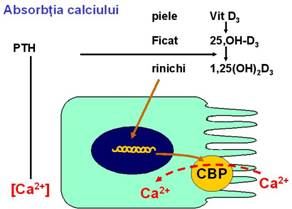
ABSORBTIA CALCIULUI Calciul este absorbit activ prin intermediul unui carrier de la nivelul polului apical CBP (proteina fixatoare de calciu). Sinteza acestei proteine se realizeaza in enterocit si regleaza rata absorbtiei intestinale in functie de controlul exercitat de vitamina D si de parathormon. |
|
ABSORBTIA FIERULUI
(vezi curs sange - eritrocit)
ABSORBTIA VITAMINELOR
Vitaminele liposolubile (A, D, E, K) se absorb similar lipidelor
Vitaminele hidrosolubile se absorb prin transport pasiv, cu exceptia:
Vitamina B1: transport activ Na+ dependent
Vitamina C: transport pasiv + transport activ Na+ dependent
Vitamina B12: Complex cu factorul intrinsec -absorbtie mediata de receptor in ileonul terminal
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 5310
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved