| CATEGORII DOCUMENTE |
| Alimentatie nutritie | Asistenta sociala | Cosmetica frumusete | Logopedie | Retete culinare | Sport |
Schimburile de substante prin membrana celulara
Procesul de traversare a substantelor prin membrana se numeste transport membranar. Mecanismul de transport depinde de permeabilitatea membranei pentru o substanta data, proprietate care este:
selectiva - membrana permite trecerea numai a anumitor substante;
asimetrica membrana este traversata numai intr-un singur sens;
dinamica - depinde de metabolismul celular si de conditiile de mediu.
In functie de dimensiunea substantei de transport se descriu sisteme de microtransfer (pentru micromolecule) si sisteme de macrotransfer (pentru macromolecule).
In functie de natura fortelor care asigura sistemele de microtransfer se descriu:
Transport pasiv: - difuziune simpla - prin dublu strat lipidic
- prin canale ionice
- difuziune facilitata
- osmoza
Transport activ - primar
- secundar
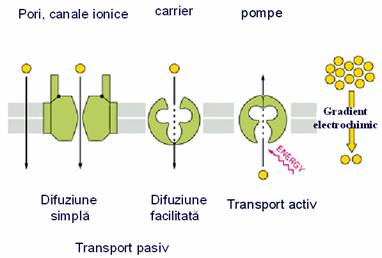
1.TRANSPORTUL PASIV
Caracteristici
- se desfasoara spontan, sub actiunea unor forte fizice
- fara consum de energie
- in virtutea unor gradiente pana la atingerea starii de echilibru:
Tipuri de transport pasiv
A. DIFUZIUNE SIMPLA
- presupune transportul in virtutea unor gradiente pana se instaleaza starea de echilibru (steady state).
- se desfasoara in ambele sensuri, gradientul de concentratie stabilind marimea difuziunii nete.
Se realizeaza:
Caracteristici:
sunt proteine integrale din structura membranei
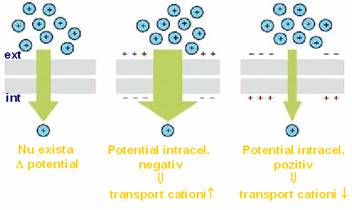
asigura transportul selectiv, unidirectional al anumitor ioni, cu diametrul < 8 nm, in sensul gradientului electrochimic al acestora.
intensitatea fluxului ionic este exprimata prin conductanta ionica, evaluata in Siemens. Canalele ionice au o conductanta de ordinul pS (1pS = 10-12 S), prin canal putand trece 1 milion ioni/sec.
In ansamblu, un canal ionic prezinta:
canal de trecerea al ionilor - porul sau canalul ionic propriu-zis;
filtrul de selectivitate - prin dimensiunile, forma si sarcina sa electrica asigura permeabilitatea selectiva a canalului;
una sau doua porti - configuratii moleculare tranzitorii datorita carora canalele ionice prezinta o dinamica ("gating mecanism");
un senzor - structura receptiva la actiunea unor factori de control, celulari si extracelulari, care comanda dinamica portilor.
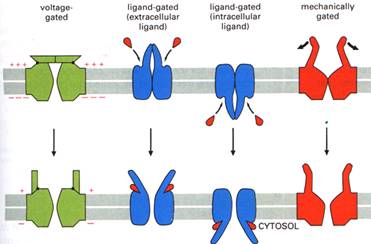
In functie de natura factorilor care controleaza dinamica canalelor ionice se descriu:
canale ionice voltaj-dependente;
canale ionice operate mecanic;
canale ionice operate de ligand.

sunt prezente in neuroni si in variate tipuri de celule musculare.
- dinamica lor este dependenta de valoarea potentialului transmembranar, iar fluxul de ioni determina, la randul sau, modificarea potentialului transmembranar.
- Se descriu: canale rapide de Na+, canale de K+, canale lente de Ca2+ si canale de Cl-.
a1. Canale rapide de na+
se gasesc in membrana neuronului, a fibrei musculare cardiace si scheletice.
se deschid in urma depolarizarii membranei celulare pana la valoarea potentialului prag si permit un influx masiv de Na+ cu efect depolarizant. Se inchid in momentul repolarizarii.
Canalul de Na+ prezinta o poarta de activare - notata m si una de inactivare - notata h. Portile prezinta urmatoarea dinamica:
la valoarea potentialului de repaus - canalul de Na+ este inchis, bariera de activare m este inchisa si cea de inactivare h este deschisa;
in faza de depolarizare - canalul de Na+ este deschis, poarta de activare m se deschide atunci cand se atinge o depolarizare cu amplitudinea de 10-15 mV si canalul permite trecerea curentului de Na+ spre interior;
in faza de repolarizare - canalul de Na+ este inactivat, poarta h se inchide lent, inactivand dupa un scurt interval (1 ms), curentul de Na+ rezultat in urma deschiderii portii m. Canalul in stare inactivata, are poarta m deschisa si poarta h inchisa si dupa 6 ms trece in stare de repaus.
a2. Canale de k+
se gasesc in toate structurile excitabile
- au rol in mentinerea potentialului membranar de repaus si in repolarizarea membranei care urmeaza dupa marile depolarizari din timpul potentialului de actiune.
- controleaza nivelul excitabilitatii celulare.
Aceste canale au o singura poarta de activare (n) care se deschide tardiv, la 6 ms dupa stimulare, si care permite un eflux de K+ cu efect hiperpolarizant.
a3. Canale lente de ca2+
sunt canale lente
- prezinta o permeabilitate selectiva atat pentru Ca2+, cat si pentru Na+.
- sunt activate in urma depolarizarii celulare si permit un influx de Ca2+ cu implicatii deosebite in excitabilitatea, contractia si secretia celulara.
- se descriu trei tipuri de canale de Ca2+, notate L, T si N. Dintre acestea, canalul de tip L se gaseste in fibra musculara cardiaca si neteda, avand rol in cuplarea excitatiei cu contractia.
- sunt sensibile la actiunea unor "blocante" utilizate in terapeutica, cum sunt Nifedipina, Verapamilul si Diltiazemul.
a4. Canale de cl-
se gasesc in celulele epiteliului tractului digestiv si celulele epiteliului cailor aeriene
- se deschid in urma depolarizarii, iar influxul de Cl- rezultat are un efect repolarizant sau hiperpolarizant.
- se activeaza la intinderea membranei celulare (stretch).
- Se gasesc in:
celulele musculare netede (pe sarcolema), asigurand un influx de Na+ si Ca2+ cu rol depolarizant.
celulele endoteliale
celule senzoriale. Ex: in celulele auditive din canalul cohlear, deformarea mecanica a cililor deschide canale ionice urmate de un influx cationic depolarizant, generator al potentialului local de receptor.
- sunt reglate prin modificari conformationale induse de actiunea unor substante biologic active extracelulare (ligand extracelular) sau a unui mesager intracelular (ligand intracelular).
c1. Canalele ionice operate de liganzi extracelulari - participa la formarea unor complexe structurale receptoare. Cele mai importante sunt prezentate in tabel:
|
Canal |
Receptor |
Ligand |
Localizare |
Efect postsinaptic |
|
Na+ |
nicotinic |
Ach |
Sinapse de tip excitator: placa motorie, SNC, SNV periferic |
Influx de Na+ Depolarizare |
|
Cl- |
GABA-ergic |
GABA |
Sinapse de tip inhibitor: SNC |
Influx de Cl- Hiperpolarizare |
|
Na+ K+ |
specific |
AA excitatori (glutamat) |
Sinapse de tip excitator: SNC |
Influx de Na+ Eflux de K+ |
c2. Canalele ionice operate de liganzi intracelulari -
prezinta in general situsuri de fixare a liganzilor la nivelul portilor de activare ale canalului.
Ex: - la nivelul celulelor epiteliale (epitelii de absorbtie),
- principalii liganzi intracelulari care moduleaza dinamica unor canale ionice de Na+, K+, Cl-, dar si de apa (acvaporine) sunt: Ca2+ citosolic, AMPc si GMPc.
c3. Canalele ionice operate de receptori prin proteine G - au o dinamica reglata de proteine reglatorii intramembranare ce opereaza direct sau prin mesageri secundari.
c4. Canalele intercelulare (conexonii)
se gasesc la nivelul jonctiunii permeabile dintre celule, numita gap jonction.
fiecare conexon este format din 2 hemicanale, apartinand celor doua membrane aflate in apozitie. Fiecare hemicanal are 6 unitati proteice care delimiteaza un por cu diametru de 1,5 nm, permeabil pentru molecule cu diametru mic, cum ar fi: apa, ionii, nucleotidele, vitaminele, produsii de metabolism celular si mesagerii secundarii.
acidoza intracelulara si cresterea Ca2+ citosolic suprima conductanta conexonilor.


Conexonii sunt numerosi la nivelul fibrei musculare cardiace si netede, functionand ca adevarate sinapse electrice ce asigura trecerea excitatiei de la o celula la alta.
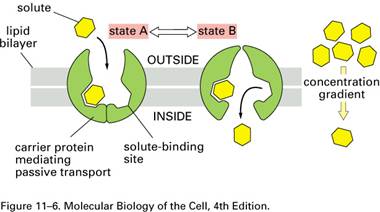
B. DIFUZIUNEA FACILITATA
- transportul pasiv al substantelor organice neionizate care nu pot traversa membrana decat cu ajutorul unei proteine "carrier" specifice. Substanta este fixata pe partea membranei unde aceasta este in concentratie mai mare, iar proteina "carrier", prin modificari conformationale care nu necesita consum energetic, o transfera pe partea membranei unde concentratia substantei este mai mica.
- viteza de transfer este mai mare in comparatie cu difuziunea simpla
- rata difuziunii depinde de gradientul de concentratie si de disponibilitatea transportorului. Astfel, difuziunea facilitata are un caracter limitativ - se poate desfasura pana la o valoarea maxima (Tmax).
- Capacitatea de transport poate fi influentata de interventia anumitor substante biologic active. De exemplu, difuziunea facilitata a glucozei creste de 10-20 de ori in prezenta insulinei, prin cresterea numarului de transportori specifici pentru glucoza.
Exemple: absorbtia fructozei si a unor AA la nivelul epiteliului intestinal

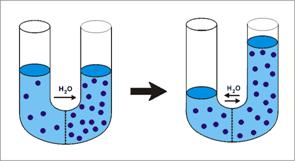
C. OSMOZA
- reprezinta procesul de trecere a apei printr-o membrana semipermeabila, dinspre solutia mai diluata spre cea mai concentrata (in sensul gradientului de concentratie al apei).
- In functie de permeabilitatea selectiva a membranei, gradientul de presiune osmotica determina sensul si marimea transferului de apa (solvent) si de particule osmotic active (solviti) - NaCl, glucoza, uree, proteine:
daca doua solutii cu osmolaritati diferite sunt puse in contact printr-o membrana permeabila, atat pentru apa, cat si pentru solviti, gradientul de presiune osmotica determina transferul solvitilor dinspre solutia cu presiune osmotica mai mare spre cea cu presiune osmotica mai mica, iar a apei in sens invers. Starea de echilibru osmotic este atinsa fara ca volumele celor doua solutii sa se modifice;
daca doua solutii cu osmolaritati diferite sunt puse in contact printr-o membrana semipermeabila, permeabila pentru apa, dar nu si pentru solviti, gradientul de presiune osmotica determina transferul apei dinspre solutia cu presiune osmotica mai mica spre cea cu presiune osmotica mai mare. Atingerea starii de echilibru osmotic este asociata cu modificarea volumelor celor doua solutii. In acest caz, gradientul de presiune osmotica asigura difuziunea neta a apei in sensul gradientului sau de concentratie, fie prin matricea fosfolipidica, fie prin canale de apa.

2. TRANSPORTUL ACTIV
Caracteristici:
se realizeaza contra gradientului
- este posibil numai in prezenta unui proteine "carrier" specifice. Proteinele transportoare prezinta un numar limitat de situsuri de legare pentru substanta pe care o transporta sau pentru un numar limitat de substante, inrudite chimic. De exemplu, glucoza si aminoacizii folosesc transportori specifici, dar mai multi aminoacizi inruditi chimic pot utiliza aceeasi proteina transportoare, participand la un transport activ competitiv.
- are un caracter limitativ, determinat de disponibilitatea transportorului (rata de transfer a substantei este direct proportionala cu concentratia acesteia pana in momentul in care situsurile sunt complet ocupate si se atinge rata maxima de transport (Tmax).
- se realizeaza cu consum de energie.
Transportul activ este principalul mecanism prin care se asigura diferenta de concentratie a substantelor intre mediul intracelular si extracelular. Prin mecanism activ sunt transportate molecule organice mari (monozaharide, AA, proteine) si ioni (Na+, K+, Ca2+, Fe2+, H+, etc).
Transportul activ depinde de energetica celulara, energia necesara fiind proportionala cu logaritmul raportului dintre concentratiile de o parte si de alta a membranei:
Energie (cal/osm) = 1400 log C1/C2
Conform acestei formule, transportul contra gradient a unui osmol de substanta, la o diferenta de concentratie de 10 ori, necesita 1400 calorii.
In functie de sursa de energie care asigura desfasurarea procesului, transportul activ poate fi primar si secundar.
A. Transportul activ primar
- se caracterizeaza prin faptul ca proteina transportoare este in acelasi timp si ATP-aza. Aceasta hidrolizeaza ATP-ul in ADP si fosfat anorganic (Pi) si elibereaza energia necesara transportului. ATP-azele implicate in transportul ionilor, contra gradientelor electrochimice, se numesc pompe ionice. Cele mai cunoscute pompe ionice sunt:
pompa Na+/K+ (Na+/K+-ATP-aza);
pompa H+/K+ (H+/K+ - ATP-aza);
pompa de Ca2+ (Ca 2+-ATP-aza);
pompele protonice (H+-ATP-azele).
Pompele ionice
Pompele ionice sunt ATP-aze transportoare specifice, contra gradient si cu mare consum energetic.
Cele mai importante pompe ionice sunt:
a. Pompa Na+/K+
- este cea importanta dintre ATP-azele de transport, Mg2+ dependenta.
- Structura: este formata din 2 subunitati a - catalitice si 2 subunitati b - reglatorii:
subunitatea a prezinta situsuri de fixare interne pentru Na+, ATP si Mg2+,
situsuri de fixare externe pentru K+ si ouabaina
situs de fosforilare - de fixare a gruparii Pi;
subunitatea b asigura translocarea pompei din citoplasma in membrana si distributia polarizata a pompei pe suprafata celulara.
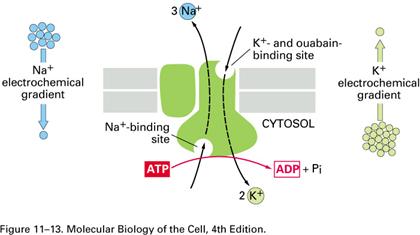

- Mecanism: transporta 3Na+ spre exterior si 2K+ spre interior, pentru fiecare molecula de ATP hidrolizata, respectand urmatoarele secvente:
fixarea gruparii Pi (sub actiunea unei kinaze) la nivelul situsului de fosforilare, urmata de cresterea afinitatii pompei pentru Na+;
utilizarea energiei pentru modificarea conformationala a proteinei, cu expunerea situsurilor de Na+ pe fata externa a membranei si disocierea Na+ in mediul extracelular;
ocuparea situsurilor externe cu 2 K+ ;
indepartarea gruparii Pi (sub actiunea unei fosfataze), urmata de cresterea afinitatii pentru K+ ;
revenirea proteinei la conformatia initiala, cu expunerea situsurilor de K+ pe fata interna membranei si disocierea K+ in mediul intracelular.
- Roluri
- mecanism principal de asigurare a homeostaziei celulare.
- mentine potentialul de repaus si participa la refacerea echilibrului ionic dupa repolarizare (fiind o pompa electrogenica, determina o incarcare electronegativa a membranei la interior). Activarea sa in exces determina hiperpolarizarea membranei, iar inhibitia sa determina o usoara depolarizare
- intretine gradientul de Na+ necesar transportul activ secundar.
Pentru realizarea acestor functii, pompa Na+/K+ consuma o mare parte din energia celulara, aproximativ 50% pentru neuron si nefrocit si 10% pentru fibra musculara si hepatocite.
- Factori reglatori:
- este activata de insulina, hormonii tiroidieni, catecolamine
- este inhibata de ouabaina si de tonicardiace (digoxin).
b. Pompa H+/K+
- este prezenta in celula parietala gastrica unde are rol in formarea HCl.
- scoate din celula 1H+ si introduce 1K+,
c. Pompa de Ca2+
- este caracteristica fibrei musculare:
la nivelul sarcolemei; rol: expulzeaza Ca2+ in mediul extracelular
la nivelul membranei reticulului sarcoplasmatic: asigura recaptarea Ca2+ in principalul depozit de Ca2+ intracelular.
- Rolul pompelor de Ca2+ este cel de scadere a concentratiei Ca2+ citosolic de la 10-5 M la 10-7 M, necesara relaxarii musculare.
d) Pompele de H+
- se intalnesc:
la nivelul membranelor mitocondriale, fiind implicate in transportul electronilor in timpul fosforilarii oxidative.
la nivelul polului apical al nefrocitului distal, asigurand eliminarea renala a H+, cu rol in mentinerea echilibrului acido-bazic.
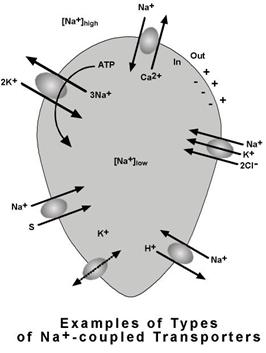
B. Transportul activ secundar
- asigura transportul unei substante cuplat cu transportul de Na+, in cadrul unui mecanism de transport care se numeste simport, in aceeasi directie (cotransport) sau in directii opuse (antiport sau schimbator ionic). Substanta transportata poate fi un ion sau o substanta organica.
Mecanism de transport prezinta urmatoarele caracteristici:
transportul de Na+ are loc intracelular si in sensul gradientului sau de concentratie mentinut de pompa Na+/K+ (scoate in permanenta 3Na+ si introduce 2K+ pentru 1 ATP);
transportul substantei cuplate are loc contra gradient si in functie de directia de transfer, acesta poate fi:
cotransport - se desfasoara in aceeasi directie cu transportul de Na+; de exemplu - Na+/glucoza, Na+/aminoacizi;
schimbator ionic (antiport) - se desfasoara in directie opusa cu transportul de Na+; de exemplu - Na+/Ca2+, Na+/H+.
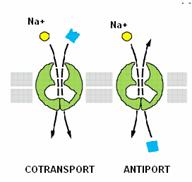

Cotransportorii
Cotransportorii asigura un transport activ secundar pentru ioni si substante organice. Transportul se desfasoara in aceeasi directie cu transportul de Na+.
Cotransportorii se clasifica in:
cotranportori ionici - Na+/Cl-, Na+/K+/2Cl-, 2Na+/PO34-;
cotransportori organici - Na+/glucide, Na+/aminoacizi.
Acest tip de transport poate fi:
electroneutru, cand nu modifica potentialul transmembranar. Ex: cotransportul Na+/Cl- este electroneutru deoarece transporta doua molecule cu sarcini electrice opuse, care se anuleaza reciproc.
electrogen, cand determina modificari de potential transmembranar. Ex: in cazul cotransportului Na+/glucoza, la fiecare molecula de glucoza transportata spre interior, mediului intracelular i se mai este adaugata o sarcina pozitiva suplimentara care genereaza o depolarizare.
Schimbatorii ionici
Schimbatorii ionici sunt proteine care cupleaza transportul a doi ioni in directii opuse, in cadrul unui transport activ secundar. In functie de natura ionilor implicati in transport, se deosebesc:
schimbatori cationici - Na+/H+, Na+/Ca2+;
schimbatori anionici - Cl-/HCO3-.
a. Schimbatorul Na+/H - electroneutru, scoate din celula 1H+ si introduce 1Na+, contribuind la mentinerea constanta a pH-ului intracelular. Acest aspect este foarte important in procese biologice cum ar fi: proliferarea, diferentierea celulara si sensibilitatea celulei la hormoni.
b. Schimbatorul Na+/Ca2+ - electrogen, scoate din celula 1Ca2+ si introduce 3Na+, fiind important mai ales pentru celula miocardica de lucru aflata in repaus, cand participa la mentinerea unei concentratii scazute a Ca2+ citoplasmatic.

|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 8634
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved