| CATEGORII DOCUMENTE |
| Agricultura | Asigurari | Comert | Confectii | Contabilitate | Contracte | Economie |
| Transporturi | Turism | Zootehnie |
BIOTEHNOLOGII AGRICOLE
REALIZAREA UNUI BIOPREPARAT PE BAZA UNUI CONSORTIU DE BACTERII FIXATOARE DE AZOT
CUPRINS:
CAPITOLUL I. INTRODUCERE
CAPITOLUL II. STADIUL ACTUAL AL CUNOSTINTELOR IN DOMENIU
II.1. Rolul azotului si al fixarii biologice
II.1.1. Tipurile de microorganisme fixatoare de azot
II.2.1. Incadrarea sistematica a rizobiilor
II.2.2. Specificitatea de gazda a rizobiilor
II.2.3. Efectele inocularii cu Rhizobium - formarea nodozitatilor
II.2.3.1. Structura nodozitatilor
II.2.3.2. Functionarea nodozitatilor
II.2.4. Fenomenul de quorum sensing la bacteriile fixatoare de azot din grupul Rhizobium
II.3. Genul Azospirillum
II.3.1. Specificitate si variabilitate
II.3.2. Efectele inocularii cu Azospirillum asupra plantelor
II.3.2.1. Modul de actiune asupra plantelor
II.3.2.2. Efectele hormonale ale bacteriei asupra cresterii plantelor
II.3.2.3. Fixarea azotului
II.3.2.4. Imbunatatirea dezvoltarii radacinii
II.3.2.5. Efectele inocularii asupra dezvoltarii radacinilor
II.3.2.6. Colonizarea radacinilor
CAPITOLUL III. MATERIALE SI METODE
III.1. Izolarea unor tulpini bacteriene din grupul Rhizobium
III.1.1. Urmarirea dezvoltarii culturii in mediul lichid
III.1.2. Estimarea numarului de microorganisme
III.2. Caracterizarea unei tulpini de Rhizobium cu microsistemul Biolog
III.3. Cresterea rhizobiilor pe diferite medii de cultura
III.3.1. Cultivarea bacteriilor din grupul Rhizobium pe mediu hidrogelifiat
III.4. Verificarea pastrarii caracteristicilor biologice ale rhizobiilor
III.5. Elaborarea unui model experimental pentru evidentierea fenomenului stingerii de QS la bacteriile PGPR
III.6. Caracterizarea unor tulpini de Rhizobium ce prezinta activitate ACC deaminazica si care noduleaza fasolea
III.7. Estimarea eficacitatii fixarii simbiotice a azotului prin determinarea hidrogenului
III.8. Cultivarea unor tulpini de Azospirillum brasiliense pe diferite medii de cultura
III.9. Testarea activitatii biologice a biomasei de Azospirillum
III.10. Tehnologia de realizare a inocului de laborator
III.11. Conditionarea biomaselor de Azospirillum si Rhizobium ca biopreparate aglomerate - Floculi
CAPITOLUL IV. REZULTATE SI DISCUTII
IV.1. Izolarea si cresterea pe diferite medii
IV.2. Caracterizarea unei tulpini de Rhizobium la Biolog
IV.3. Cultivarea rhizobiilor pe diferite medii
IV.4. Verificarea pastrarii caracteristicilor biologice ale rhizobiilor
IV.5. Evidentierea fenomenului de QS la bacteriile inoculante
IV.6. Caracterizarea unor tulpini de Rhizobium care prezinta activitate ACC deaminazica si care noduleaza fasolea
IV.7. Estimarea eficacitatii fixarii simbiotice a azotului prin determinarea hidrogenului
IV.8. Cultivarea unei tulpini de Azospirillum brasiliense pe diferite medii de cultura
IV.9. Testarea activitatii biologice a biomasei de Azospirillum
IV.10. Conditionarea tulpinilor bacteriene FL220 si SP001 sub forma de agregate - floculi
CAPITOLUL V. CONCLUZII
CAPITOLUL VI. BIBLIOGRAFIE
CAPITOLUL I. INTRODUCERE
Unicitatea microorganismelor, natura lor adesea imprevizibila si capacitatile biosintetice date de conditii specifice de mediu si culturale, le fac sa fie cei mai probabili candidati pentru rezolvarea anumitor probleme dificile atat in stiinta cat si in domeniile economice.
Diferitele cai pe care microorganismele au fost utilizate in ultimii 50 ani pentru progresul tehnologiei medicale, al sanatatii umane si animale, al ingineriei genetice, protectiei mediului, biotehnologiei agricole si a tratamentelor mai eficace ale reziduurilor agricole si menajere furnizeaza o extrem de impresionanta lista de realizari. Multe dintre aceste progrese tehnologice nu ar fi fost posibile prin utilizarea in exclusivitate a metodelor tehnice, chimice si fizice, sau daca ele nu ar fi fost practice sau economic fezabile.
Desi in ultimii ani tehnologiile microbiene au fost utilizate pentru solutionarea unor variate probleme agricole si de mediu, ele nu au fost larg acceptate de comunitatea stiintifica, deoarece adesea este dificil sa se reproduca considerabil efectele lor benefice. Microorganismele sunt eficiente numai cand se afla in conditiile optime pentru metabolizarea substraturilor specifice incluzand apa, oxigenul (in functie de natura microorganismelor), pH si temperatura mediului. Realizari semnificative au fost facute in sensul corelarii cu necesitatile pietei datorita noilor tehnologii.
Deoarece microorganismele sunt utile la eliminarea problemelor asociate cu utilizarea fertilizantilor si pesticidelor de sinteza, ele sunt acum larg aplicate in exploatatiile agricole in sistem organic sau durabil (Higa, 1991; Parr si col., 1994).
De multi ani, microbiologii si specialistii in ecologie microbiana au tins sa diferentieze microorganismele edafice in benefice si daunatoare, in functie de activitatile lor si de modul cum afecteaza calitatea solului, cresterea plantelor, recolta si sanatatea plantelor. Astfel, microorganismele benefice sunt cele care pot fixa azotul atmosferic, descompun resturile organice, detoxifica pesticidele, represeaza bolile plantelor si patogenii edafici, recicleaza si sporesc accesibilitatea nutrientilor plantelor, produc molecule organice simple pe care plantele le pot absorbi, complexeaza metalele grele, solubilizeaza nutrientii insolubili, produc polizaharide pentru imbunatatirea structurii solului si produc compusi bioactivi de tipul vitaminelor, hormonilor si enzimelor care stimuleaza cresterea plantelor.
Aceste microorganisme care maresc fertilitatea solului si care contribuie astfel la dezvoltarea plantelor au fost numite biofertilizatori si sunt frecvent utilizati ca inoculanti in practica agriculturii durabile.
Tipurile de microorganisme care produc compusi bioactivi de tipul vitaminelor, hormonilor si enzimelor sunt denumite fitostimulatori, si sunt studiate de asemenea pentru obtinerea de biopreparate.
Microorganismle daunatoare sunt acelea care pot induce boli la plante, stimuleaza patogenii de sol, inhiba germinarea semintelor, imobilizaza nutrientii si produc substante toxice si de putrefactie care afecteaza negativ cresterea si sanatatea plantelor. Anumite microorganisme sunt insa antagoniste ale acestora, si pot preveni infectiile. Mecanismul de prevenire consta in concurenta celor doua categorii pentru accesul la nutrienti si/sau producerea unor metaboliti secundari - enzime extracelulare, antibiotice si alte componente inhibitoare de crestere. Alte microorganisme din sol produc substante care activeaza mecanismele naturale de aparare ale plantelor impotriva patogenilor. Aceste categorii de microorganisme descrise mai sus au primit denumirea generica de biopesticide.
Higa (1991) a dezvoltat conceptul de microorganisme eficace (ME), care se refera la culturi amestecate de microorganisme benefice ce se gasesc in mod natural in mediu si pot fi aplicate ca inoculanti pentru cresterea diversitatii microbiene in soluri si plante.
S-a demonstrat ca, inocularea culturilor de ME in ecosistemul sol/planta, poate imbunatati calitatea si sanatate solului si poate duce la cresterea dezvoltarii productiei si calitatii culturilor. ME contin specii selectionate de microorganisme benefice care sunt mutual compatibile unele cu altele si pot coexista in cultura lichida.
Biotehnologia agricola, ca stiinta inovativa, duce la realizarea unor noi soiuri de plante, agenti de combatere biologica si tehnici de protectie a culturilor care optrimizeaza atat protectia cat si productia agricola, oferind mijloace complementare produselor si metodelor traditionale de cultivare a plantelor. Este important sa se ajunga la un echilibru intre mentinerea unor culturi sanatoase, profitabile si menajarea populatiilor care traiesc in terenurile cultivate si in jurul acestora. De aceea, produsele de protectie a culturilor rezultate in urma unor procese biotehnologice sunt cele mai adecvate, ele fiind lipsite de nocivitate pentru om si mediu atat in timpul productiei cat si al utilizarii.
Productia agricola este amenintata de declinul productivitatii terenului agricol ca rezultat al practicilor neconvenabile de exploatare a solului si apei si de eliberarea constanta a produselor chimice din ce in ce mai toxice pentru oameni si animale (Gregory, 1989).
O cale de evitare a acestui pericol o constituie utilizarea biofertilizantilor, o alternativa ieftina si sigura pentru ingrasamintele chimice conventionale. Aceasta consta din includerea in microbiocenozele agricole a populatiilor de fixatori de azot simbiotici si/sau asociativi din genurile Rhizobium, Bradyrhizobium, Synorhizobium si Azospirillum, pentru care s-a demonstrat o rata de fixare a azotului atmosferic de pana la 70% din necesarul de azot.
Biopesticidele, alaturi de biofertilizatori se constituie intr-o alternativa de mare insemnatate - control biologic - la utilizarea de pesticide si fertilizatori clasici de natura chimica.
Lucrarea a avut ca scop realizarea unui biopreparat mixt, cu actiune complexa, de nutritie, stimulare si protectia plantelor. Lucrarea a fost realizata in cadrul proiectului PN2 MIMOSA - "Inoculanti microbieni pentru sisteme de agricultura durabila", iar experimentele aferente lucrarii s-au desfasurat in laboratorul de organisme utile - bacteriologie de la Institutul de Cercetare si Dezvoltare pentru Protectia Plantelor, Bucuresti.
CAP.II. STADIUL ACTUAL AL CUNOSTINTELOR IN DOMENIU
II.1. Rolul azotului si a fixarii biologice
Azotul este un element care intra in alcatuirea proteinelor, acizilor nucleici si a multor alte molecule biologice fiind, deci, un element indispensabil pentru viata.
Studiile efectuate pe plan mondial au demonstrat ca fertilizarea contribuie cu circa 40% la cresterea recoltelor per unitate de suprafata. Dintre ingrasaminte, azotul ocupa rolul principal prin aportul sau in determinarea sporului de recolta si prin ponderea sa. Prin urmare, satisfacerea nevoilor alimentare ale populatiei globului in continua crestere, precum si furnizarea unora dintre materiile prime pentru industriile care si le iau de la agricultura, depind in mare masura, de asigurarea cantitatilor corespunzatoare de azot alaturi de celelalte elemente necesare metabolismului plantelor.
Sursele de azot, asa cum sunt cunoscute la ora actuala, sunt reprezentate de ingrasamintele minerale, ingrasamintele organice (gunoaie, namoluri si alte reziduuri) si formele alternative biologice (bacterii fixatoare de azot atmosferic simbiotice, nesimbiotice si asociative).
Agricultura durabila, strategia mondiala actuala a dezvoltarii agricole, presupune practici ecologic sanatoase si economic atractive. Fixarea biologica a azotului este o tehnologie care raspunde ambelor deziderate, oferind mijloace de reducere a costurilor si a inputurilor energetice.
Practica utilizarii leguminoaselor simbionte pentru imbunatatirea calitatii solurilor dateaza din timpurile prebiblice, dar a capatat valoare comerciala in secolul XIX, dupa descoperirea bacteriilor care produc nodozitati.
Pentru aprecierea importantei procesului biologic de fixare a azotului trebuie sa se tina seama de considerente majore, cum este cel al utilitatii pentru lumea vie (prin implicarea, alaturi de fotosinteza, in producerea de substanta organica primara, combustibil, fibre) si cel al utilitatii economice (reprezentand sursa naturala si preponderenta de azot accesibil plantelor de cultura, conservarea umiditatii solului, prevenirea eroziunii) (Bohlool, 1990).
Procesul de fixare biologica realizeaza reducerea azotului din atmosfera, unde se gaseste in cantitati mari, de aproape 77000 tone deasupra fiecarui hectar de sol, pe seama energiei solare inepuizabile, captata fotosintetic. Deci, procesul este neconsumator de energie fosila, supus autoreglarii sistemului in care se desfasoara si nepoluant pentru mediu.
In 1977 Hardy a facut urmatoarea estimare a evolutiei in timp si a raportului dintre fixarea biologica de azot si alte surse:
Tabel 1. Estimarea evolutiei in timp si a raportului dintre fixarea biologica de azot si alte surse.
|
Specificare | ||||
|
Tehnologii curente |
Tehnologii noi |
|||
|
Fixarea biologica | ||||
|
Fixarea industriala | ||||
|
Alte surse de azot fixat | ||||
|
Total | ||||
Pentru satisfacerea necesarului de hrana de la sfarsitul secolului XX-inceputul secolului XXI, se impune cresterea coeficientului de utilizare de catre plante a ingrasamintelor cu azot, elaborarea si implementarea unor tehnologii economice de producere industriala a ingrasamintelor chimice si cresterea eficientei biologice prin optimizarea proceselor naturale (fotosinteza si fixarea biologica de azot).
Capacitatea de a fixa azotul atmosferic este o insusire care apartine atat unor procariote (bacterii fototrofe sau heterotrofe, cianobacterii si actinomicete), libere sau asociate cu plantele, grupate sub denumirea generica de microorganisme diazotrofe, cat si, in pofida a ceea ce se credea pana nu de mult, unor eucariote din regnul vegetal.
Numeroase cercetari au semnalat prezenta si abundenta unor diverse grupe de microorganisme fixatoare de azot, in cele mai diferite conditii de mediu. Fixarea de azot de catre heterotrofele libere, in mediile naturale, nu este semnificativa decat in prezenta unor surse corspunzatoare de energie si carbon. Cele mai mari cantitati de azot sunt fixate in rizoplanul si rizosfera anumitor plante datorita exsudatelor lor radiculare.
Conditia obligatorie pentru ca un organism sa poata realiza reducerea azotului atmosferic, in conditii normale de temperatura si presiune, este detinerea unei informatii genetice pentru sinteza si controlul unui sistem enzimatic complex denumit "nitrogenaza". Sistemul enzimatic nitrogenazic este format din doua componente proteice diferite, care functioneaza asociat intr-un ciclu catalitic. Sinteza sistemului enzimatic nitrogenazic este guvernata de un grup de gene cunoscute sub denumirea de gene nif.
Desfasurarea procesului de fixare biologica a azotului este conditionata de numerosi factori endogeni si exogeni. Acesti factori pot fi clasificati, dupa gradul de participare, in factori primari, secundari si tertiari.
In categoria factorilor primari sunt cuprinsi factorii endogeni implicati direct in reactia centrala a fixarii: concentratia de nitrogenaza, gradul de saturare al nitrogenazei cu substratul de reactie si concentratia produsului rezultat - amoniul. Nivelul cantitativ al nitrogenazei diazotrofilor nu este factor limitativ pentru fixare la temperaturi in jur de 200C, dar devine limitativ cand temperaturile scad sub acest nivel (Hardy si Havelka, 1976).
Alaturi de factorii primari, alti determinanti ai procesului de fixare sunt reprezentati de caile de asimilare ale amoniacului produs, mecanismele de represie enzimatica, mecanismele de excludere a ionilor de hidrogen din apropierea nitrogenazei, sistemul enzimatic pentru conversia molibenului intr-o forma catalitic activa.
Factorii primari indispensabili pentru desfasurarea procesului de fixare a azotului trebuie priviti in stransa interdependenta cu un factor secundar, fotosinteza. In cursul perioadei de crestere, atat intensitatea fotosintezei cat si activitatea de fixare a azotului de catre nodozitati merg paralel si ating valori maxime inaintea infloritului. Dupa aceasta se instaleaza o competitie privind repartizarea produsilor asimilati intre organele reproducatoare si nodozitati, tradusa prin diminuarea progresiva a activitaatii de fixare a azotului.
Dependenta stransa a procesului de fixare a azotului atmosferic de fotosinteza se manifesta si prin variatii nictemerale. Analiza acestora indica faptul ca activitatea maxima coincide cu perioada de fotosinteza cea mai intensa.
Conditionarea fixarii azotului de fotosinteza, in cazul rizocenozelor diazotrofe, se face prin intermediul exsudarii prin radacini a unei parti din produsii de fotosinteza. Nivelul cantitativ si diversitatea calitativa a exsudatelor radiculare sunt determinate de tipul de fotosinteza a plantelor. Bogatia exsudatelor radiculare ale plantelor favorizeaza dezvoltarea bacteriilor diazotrofe in detrimentul altor grupe de microorganisme cu necesitati mai variate.
Factorii tertiari sunt reprezentati, mai ales, de factorii determinanti pentru procesul de fotosinteza alaturi de care intervin, prin mecanisme diferite, o serie de alti factori de mediu, unii generali, altii particulari. (Balandreau, 1975).
Factorii generali care actioneaza asupra fixarii azotului, prin intermediul fotosintezei, ating maximum de corelare in cazul simbiozelor de la leguminoase si sunt: intensitatea energiei luminoase, dimensiunea complexului fotosintetizator (densitatea plantelor, suprafata foliara totala), potentialul hidric, competitia intre maturarea plantei si necesitatile de functionare ale nodozitatilor, cresterea concentratiei in dioxid de carbon.
Importanta acestor factori a fost demonstrata (Pate si col. 1981) prin reducerea drastica si imediata a fixarii, ca urmare a diminuarii fotosintezei prin iluminare redusa sau dupa defoliere. Marirea capacitatii fotosintetizatoare, prin diverse mijloace, stimuleaza fixarea azotului. Iluminarea suplimentara a plantelor de soia determina o crestere a activitatii de fixare de la 125 kg la 165 kg N/ha/perioada de vegetatie, imbogatirea cu dioxid de carbon a atmosferei a marit de peste 5 ori nivelul de fixare a azotului prin dublarea masei si activitatii nodozitatilor, precum si prin extinderea fazei exponentiale de crestere a plantelor. In conditii de camp, este dificil de marit capacitatea de fotosinteza dar, trebuie acordat atentie afectarii, in sens negativ, a procesului de fixare de catre densitati ridicate ale plantelor si caderi ale acestora.
Reducerea experimentala sau eliminarea consumului asimilatelor catre varfuri vegetative sau boli, au drept consecinte canalizarea produsilor de fotosinteza spre nodozitati si indirect cresterea activitatii de fixare.
Importanta factorului lumina este primordiala, dar acesta devine limitant numai cand celelalte conditii, in special temperatura, sunt optime pentru fixare.
Dintre factorii de mediu generali, tensiunea de oxigen are un rol limitant, reactia de fixare desfasurandu-se numai in conditii de anaerobioza. Expunerea indelugata la oxigen inactiveaza, in mod ireversibil, sistemul enzimatic nitrogenazic.
Prezenta in sol a unei cantitati adecvate de molibden asimilabil reprezinta o necesitate generala pentru bacteriile fixatoare de azot libere sau simbiotice. De asemenea, insuficienta sulfului in sol restrange sinteza sistemului enzimatic in a carui structura intra acest element.
Un alt factor general care conditioneaza desfasurarea procesului de fixare a azotului atmosferic este nivelul azotului mineral din sol.
Efectele inhibitoare ale azotului mineral asupra simbiozei la leguminoase au fost demonstrate, dar s-au acumulat si dovezi practice care sustin ca doze mici de azot combinat, daca sunt corect aplicate in timp, pot stimula formarea de nodozitati si fixarea de azot (Obaton si col. 1982). O explicatie a acestei contradictii ar putea fi aceea ca, multe organisme cultivate, mai ales soiurile inalt productive, poseda un potential de crestere care nu poate fi satisfacut nici de cele mai eficiente tipuri de simbioza. Stimularea fenomenului de simbioza se poate produce indirect, prin favorizarea dezvoltarii plantei, iar stadiile din ciclul de dezvoltare care beneficiaza de fertilizare sunt reprezentate de faza anterioara instalarii simbiozei si faza de umplere a bobului, cand simbioza este in declin.
O alta explicatie, sustinuta de numeroase dovezi experimentale, atat in cazul lui Rhizobium, cat si in cazul unor diazotrofi liberi cum este Azospirillum, este reprezentata de selectia spontana a unor tulpini mutante, derepresate fata de amoniu, care ar putea avea implicatii practice deosebit de interesante pentru cresterea productivitatii culturilor.
Asupra sistemelor simbiotice, aciditatea solului exercita un efect complex, explicat printr-o actiune directa, insotita de multiple efecte colaterale. Actiunea directa, se manifesta la inceputul formarii nodozitatilor, in stadiul numit "acido-sensibil" si consta in inhibarea activitatii pectinazei sau formarii mucigelului, secretat de radacinile leguminoaselor. Mucigelul inglobeaza in mod specific bacteriile simbiotice si le protejeaza de pH-ul acid al solului. Efectele colaterale se adreseaza plantelor care, in mediu acid, sufera fenomene de intoxicatie prin absorbtie marita de metale grele. De asemenea bacteriile, in conditii de aciditate, pierd capacitatea de fixare a azotului molecular.
Necesitatile in calciu ale sistemelor simbiotice de la leguminoase sunt ridicate si sunt mult mai mari decat cele ale partenerilor analizati separat. Cerintele fata de calciu sunt maxime in faza de initiere a nodozitatilor, interventia sa specifica exercitandu-se asupra mitozelor.
Aciditatea, deficienta de calciu, excesul de aluminiu si mangan se coreleaza in anumite tipuri de soluri, mai putin propice pentru simbioza la leguminoase, cum sunt solurile podzolice din Romania. Amendarea cu calciu corecteaza o parte din aceste deficiente. Solurile cernoziomice asigura instalarea si evolutia unei simbioze eficiente in fixarea azotului.
Rolul jucat de temperatura in desfasurarea proceselor de fixare a azotului este inca discutat. Se considera ca fixarea este mult mai activa in regiunile tropicale si subtropicale comparativ cu regiunile temperate. S-a demonstrat ca, 24-30 0C constituie intervalul de temperatura optim la care simbioza este functionala in regiunile temperate. Acesta coincide cu temperaturile optime pentru dezvoltarea plantelor.
II.1.1.TIPURILE DE MICROORGANISME FIXATOARE DE AZOT.
Identificarea si
selectarea microorganismelor asociate cu plante si ameliorarea
genetica, sunt strategii alternative de obtinere a recoltelor care
beneficiaza de fixare de N![]() procariota. Fixarea de N
procariota. Fixarea de N![]() (fixare
fara formare de nodozitati) de la bacterii asociate este o
metoda corespunzatoare pentru exploatarea fixarii de N
(fixare
fara formare de nodozitati) de la bacterii asociate este o
metoda corespunzatoare pentru exploatarea fixarii de N![]() in
plante neleguminoase (Bitton, 2002). S-a recunoscut si folosul bacteriilor
N
in
plante neleguminoase (Bitton, 2002). S-a recunoscut si folosul bacteriilor
N![]() fixatoare
in bioremediere. Fixarea de N
fixatoare
in bioremediere. Fixarea de N![]() poate
diminua consumul de azot necesar consortiei bacteriene folosit pentru
degradarea combustibilului diesel.
poate
diminua consumul de azot necesar consortiei bacteriene folosit pentru
degradarea combustibilului diesel.
Proprietatea de fixare
biologica a N![]() conditionata de prezenta unui
sistem enzimatic specific: nitrogenaza, este limitata la cateva genuri de
bacterii (Bains, 2004).
conditionata de prezenta unui
sistem enzimatic specific: nitrogenaza, este limitata la cateva genuri de
bacterii (Bains, 2004).
Dupa modul lor de viata, microorganismele fixatoare de azot, se impart in doua grupe mari:
Microorganisme libere fixatoare de azot
a) aerobe: Azotobacter; Azomonas; Beijerinckia; Derxia; Azotococcus; Methylococcus; Methylosinus; Aquaspirillum peregrinum, A. fasciculus; Streptomyces thermoautotrophicus; toate cianobacteriile care formeaza heterochisti; Gloeocapsa.
b) facultativ anaerobe: Bacillus polymyxa, B. macerans; Klebsiella pneumoniae, K. aerogenes, K. oxytoca; Citrobacter freundii; Enterobacter cloacae, Erwinia herbicola; tulpini de Escherichia coli si de Salmonella typhimurium la care in conditii de laborator au fost incorporate gene nif.
c) microaerofile: Xanthobacter autotrophicum; Azospirillum lipoferum, A. brasilense, A. doebereinerae, A. largimobile, A. halopraeferens, A. amazonense, A. irakense; Thiobacillus ferrooxidans; numeroase specii de cianobacterii care nu formeaza heterochisti; Rhizobium japonicum (Bull, 2004).
d) anaerobe: Clostridium, Clostridium pasteurianum; Desulfovibrio; Chromatium;
Desulfotomaculum; Chlorobium; Propionibacterium; Thiopedia; Thiocapsa;
Ectothiospira; Rhodomicrobium;Rhodospirillum; Rhodopseudomonas;
Rhodophila; Rhodobacter; Heliobacterium; Heliobacillus, Heliophilum;
Methanosarcina; Methanococcus; Methanobacterium; Methanospirillum;
Methanolobus .
Dintre microorganismele libere fixatoare de azot aerobe cele mai importante apartin familiei Azotobacteraceae.
Reprezentantii acestei famili se prezinta morfologic sub forma unor celule mari, predominant bastonase, pana la oval, dar care isi pot schimba aspectul morfologic in functie de varsta culturii si de conditiile de cultivare. Celulele sunt grupate adesea in perechi si sunt mobile datorita prezentei flagelilor peritrichi sau polari. Exista si forme imobile. Nu formeaza endospori. Unele specii formeaza microchisti rezultati din ingrosarea peretelui celulei vegetative si acumularea unor substante de rezerva. Chisturile se formeaza in conditii de lipsa de substante nutritive si deshidratare, sunt rezistente si asigura o stare latenta partiala. In conditii favorabile chisturile germineaza. Cele mai multe tulpini prezinta o capsula mucilaginoasa care inconjoara una sau mai multe celule si care este cu atat mai mare cu cat continutul mediului de cultura in glucide este mai ridicat. Aceasta capsula inconjoara si protejeaza coloniile de celulele in sol. Unele tulpini produc pigmenti fluorescenti. Sunt microorganisme heterotrofe, capabile sa fixeze azotul molecular intr-un mediu fara azot, dar in prezenta unei surse organice de carbon (Szab, 1989).
Speciile din genul Azotobacter sunt cei mai importanti
reprezentanti ai bacteriilor fixatoare de azot obligat aerobe, saprofite,
libere din sol. Pot fi determinate in toate regiunile geografice si din
toate tipurile de sol. Microchistii de Azotobacter
sunt rezistente fata de deshidratare si de radiatiile
ultraviolete, dar nu si fata de caldura (peretii
nu contin acid dipicolic). Cea mai raspandita specie a genului
este Azotobacter chroococcum, care
prezinta o mare variabilitate morfologica. Celulele tinere au
aspectul unor bastonase groase de 2 m x 4 m, cu capetele rotunjite,
izolate sau in perechi, ciliate peritrich. Celulele adulte sunt ovalare sau cu
aspectul unor coci mari de 2 m x 1,2 m si contin numeroase
granulatii. Celulele batrane sunt aproape sferice, numarul
flagelilor scade si celulele isi pierd mobilitatea. Datorita prezentei
capsulei gelatinoase preparatele de Azotobacter
chroococcum au un aspect deosebit: celulele se gasesc intotdeauna la o
anumita distanta intre ele. Este aerob si facultativ
microaerofil. Formeaza la suprafata mediilor lichide o pelicula
colorata brun pana la negru, datorita melaninei. Temperatura
optima de dezvoltare este de 25-28![]() C
si pH-ul=7-7,5. Este Gram-negativ in stadiul tanar si
Gram-variabil la maturitate. Valorifica amidon si manita.
Fixeaza cel putin 10 mg N
C
si pH-ul=7-7,5. Este Gram-negativ in stadiul tanar si
Gram-variabil la maturitate. Valorifica amidon si manita.
Fixeaza cel putin 10 mg N![]() la 1 g de carbohidrati consumati.
Exigenta fata de molibden poate fi inlocuita cu vanadiu. Nu
prezinta activitate proteolitica. Este catalaz pozitiv. Ca sursa
de azot valorifica nitrati, amoniac si aminoacizi. Azotobacter chroococcum se intalneste
aproape in toate solurile, frecventa cea mai mare fiind in solurile
cultivate, fertile (Dunca si al., 2004).
la 1 g de carbohidrati consumati.
Exigenta fata de molibden poate fi inlocuita cu vanadiu. Nu
prezinta activitate proteolitica. Este catalaz pozitiv. Ca sursa
de azot valorifica nitrati, amoniac si aminoacizi. Azotobacter chroococcum se intalneste
aproape in toate solurile, frecventa cea mai mare fiind in solurile
cultivate, fertile (Dunca si al., 2004).
Din genul Azomonas dintre speciile libere fixatoare de azot se remarca Azomonas agilis, A. insignis, A. macrocytogenes.
Multe specii din familia
Pseudomonaceae traiesc saprofit
in sol. Ele cuprind mai ales bacterii in forma de bastonas,
nesporogene. Formele mobile prezinta cili polari. Speciile de Pseudomonas au fost considerate
incapabile sa fixeze N![]() ,
dar studiile recente au dovedit capacitatea fixarii de N
,
dar studiile recente au dovedit capacitatea fixarii de N![]() si la unele specii de Pseudomonas. Pseudomonas stutzeri (in prealabil denumit Alcaligenes faecalis) este larg utilizat ca inoculant a orezului in
China. In urma inocularii orezului, Pseudomonas
stutzeri colonizeaza agresiv radacinile si gena nifH se exprima in aceste bacterii
asociate cu radacina.
si la unele specii de Pseudomonas. Pseudomonas stutzeri (in prealabil denumit Alcaligenes faecalis) este larg utilizat ca inoculant a orezului in
China. In urma inocularii orezului, Pseudomonas
stutzeri colonizeaza agresiv radacinile si gena nifH se exprima in aceste bacterii
asociate cu radacina.
Klebsiella pneumoniae se gaseste in soluri si ape, in tubul digestiv la om si animale. Este specia patogena a inbolnavirilor sistemului respirator si urogenital. Este imobila, Gram negativa, chemoorganotrofa, catalaz pozitiva si oxidaz negativa, are capsula. Din glucoza formeaza gaz, din lactoza acid. Utilizeaza glucoza si citratul ca sursa de carbon, iar amoniacul ca sursa de azot.
Din grupa bacteriilor libere anaerobe, fixatoare de azot molecular o importanta deosebita prezinta genul Clostridium.
Clostridium pasteurianum se prezinta in stadiu
vegetativ sub forma de bastonase drepte sau usor curbate de
0,5-0,8/3,4-13,2 m si sub forma de fus in stadiu sporulat. Sporii
sunt deformanti si dispusi central, terminal si
subterminal. Sporul matur este inconjurat de o capsula gelatinoasa,
de forma triunghiulara, cu capetele putin rotunjite. In unele
conditii de cultivare, aceasta bacterie da forme cu aspecte din
cele mai bizare: filamente lungi cu spori la unul din capete sau uneori se
observa desfacerea celulelor intr-o serie de formatiuni
asemanatoare cocilor. In celulele tinere citoplasma este
omogena, inainte de a incepe sporularea apare in ea glicogen si
incluziuni de amidon. Clostridium
pasteurianum este mobil, cu flageli dispusi peritrich, uneori imobil.
Este strict anaerob sau anaerob-aerotolerant. Temperatura optima de
dezvoltare este de 28-30![]() C.
Se dezvolta bine in medii ce contin hidrati de carbon
fermentescibili. Este o bacterie butirica adevarata producand in
cursul fermentarii hidratiilor de carbon acid butiric, acid acetic
si gaze.
C.
Se dezvolta bine in medii ce contin hidrati de carbon
fermentescibili. Este o bacterie butirica adevarata producand in
cursul fermentarii hidratiilor de carbon acid butiric, acid acetic
si gaze.
Numeroase alte specii de Clostridium pot fixa azotul dar in cantitati reduse. In general, s-a constatat ca speciile de Clostridium fixatoare de azot atmosferic au o raspandire mai mare decat speciile aerobe de Azotobacter. Datorita modului lor de viata si a pH-lui optim relativ scazut, ele se pot gasi in solurile mai putin aerisite, precum si in solurile de padure si pasune, compacte si acide. Ele sunt prezente insa si in solurile cultivate, bine aerisite si bogate in humus. In acest caz, sunt prezente aproape exclusiv in interiorul particulelor structurale ale solului. In majoritatea cazurilor ele accepta, ca si fixatorii de azot aerobi, o simbioza cu bacteriile celulozolitice.
A) Microorganisme simbiotice fixatoare de azot:
Bacterii simbiotice care produc nodozitati: Rhizobium, Bradyrhizobium, Sinorhizobium, Azorhizobium - leguminoase; Rhizobium - neleguminoase (Trema cannabina); actinomicete (Frankia) - angiosperme neleguminoase (Alnus, Myrica, Ceanothus, Comptonia, Casuarina); cianobacterii (Nostoc, Anabaena) - Cycas, Bowenia etc (Zarnea, 1984).
Cianobacterii simbiotice cu alte organisme eucariote: Nostoc - fungi (licheni) Peltigera, Collenia; Nostoc - hepatice (Anthocera, Blasia etc.); Anabaena - Pteridophyta (Azolla); Nostoc - angiosperme (Gunnera).
Simbioze asociative: Paspalum - Azotobacter paspali; Digitaria decumbens - Spirillum lipoferum (bacteriile cresc pe / si in radacini fara sa produca nodozitati).
Simbioze cu bacterii celulozolitice (la termite, rumegatoare etc.).
Speciile de bacterii fixatoare de azot care produc nodozitati pe radacini si pe tulpini, valide, descrise in literatura de specialitate sunt: Rhizobium leguminosarum bvs. trifolii, viciae si phaseoli, R. galegae, R. tropici, R. etli, R. gallicum, R. giardini, R. huautlense, R. mongolense, Synorhizobium meliloti, S. fredii, S. saheli, S. teranga, S. medicae, Mesorhizobium loti, M. huakuii, M. ciceri, M. tianshanense, M. mediterraneum, M. plurifarium, M. amorphae, Allorhizobium undicola, Bradyrhizobium elkanii, B. japonicum, B. liaoningense, Azorhizobium caulinodans (Berner si al., 2005).
Bacteriile implicate in fixarea simbiotica la leguminoase apartin familiei Rhizobiaceae (genurile Rhizobium, Bradyrhizobium).
In general, toate solurile normale contin bacterii de nodozitati apartinand genului Rhizobium. Aceasta nu inseamna insa ca oricare specie de planta leguminoasa gaseste in mod sigur specia de Rhizobium cu care sa intre in relatii simbiotice (Eliade si col.,1975).
II.2.1. INCADRAREA SISTEMATICA A RIZOBIILOR.
Speciile de rhizobii care produc nodozitati pe radacini si pe tulpini, valide, descrise in literatura de specialitate sunt: Rhizobium leguminosarum bvs. trifolii, viciae si phaseoli, R. galegae, R. tropici, R. etli, R. gallicum, R. giardini, R. huautlense, R. mongolense, Synorhizobium meliloti, S. fredii, S. saheli, S. teranga, S. medicae, Mesorhizobium loti, M. huakuii, M. ciceri, M. tianshanense, M. mediterraneum, M. plurifarium, M. amorphae, Allorhizobium undicola, Bradyrhizobium elkanii, B. japonicum, B. liaoningense, Azorhizobium caulinodans (Berner si col., 2005).
Bacteriile implicate in fixarea simbiotica la leguminoase apartin familiei Rhizobiaceae (genurile Rhizobium, Bradyrhizobium).
In stare libera, rizobiile se prezinta ca bacterii de forma bacilara, fara endospori, gram-negative, aerobe si mobile datorita prezentei a 1-6 flageli.
Unicitatea acestor bacterii consta in faptul ca pot induce structuri specifice la leguminoase, asa numitele nodozitati situate pe radacini sau tulpini.
Conform ultimelor studii taxonomice, in grupul rizobiilor sunt incluse genurile Rhizobium, Sinorhizobium, Mezorhizobium, Bradyrhizobium si Azorhizobium (de Lajaudie si col., 1994; Jarvis si col., 1997; Haukka, 1997).
Bacteriile difera intre ele, unele formand nodozitati cu un numar limitat de leguminoase, altele avand un cerc larg de plante gazda.
Distinctia facuta intre Rhizobium si Bradyrhizobium se bazeaza, in principal, pe rata de crestere pe extractul de drojdii si capacitatea de a utiliza diferite surse de carbon.
Genul Rhizobium consta din specii care prezinta rate mari de crestere pe extractul de drojdii si care utilizeaza numeroase zaharuri, polioli si acizi organici.
Genul Bradyrhizobium are crestere lenta, are necesitatile nutritionale mai specializate si prefera pentozele.
Genul Azorhizobium se caracterizeaza prin capacitatea de a forma nodozitati pe tulpini (Dreyfus si col., 1988; Martnez-Romero, 1994).
Speciile genului Rhizobium sunt raspandite, de regula, in rizosfera.
Celulele bacilare au diametrul de 1-1,4 x 0,6-0,8 m. Sunt Gram negative, ciliate si foarte mobile. In nodozitatile radacinilor leguminoaselor celulele iau forma literelor T, X si Y.
Sunt bacterii aerobe, heterotrofe si nu
formeaza spori. Temperatura optima de dezvoltare este de 25![]() C.
C.
Rhizobium leguminosarum pe medii de cultura formeaza colonii albicioase si mucilaginoase. Celulele au diametrul de 0,5-0,9 m si o lungime de 1,2-3 m. Celulele sunt flagelate peritrich. In sol se gasesc pe radacinile plantelor de Pisum, Lathyrus, Vicia, Lens (Dunca et al., 2004).
Celulele de Rhizobium leguminosarum var. phaseoli sunt mai mici decat la Rhizobium leguminosarum. Celulele sunt flagelate peritrich. Coloniile sunt gri albicioase si mai putin mucilaginoase ca la Rhizobium leguminosarum. In sol se gasesc pe suprafata radacinilor de Phaseolus vulgaris si Phaseolus coccineus.
Rhizobium leguminosarum var. trifolii prezinta de asemenea celule flagelate peritrich. Celulele sunt in majoritatea cazurilor piriforme si prezinta vacuole. Se gasesc in nodozitatile radacinilor de Trifolium sp. si in sol.
La Rhizobium meliloti bacteroizii din noduli au aspect de bastonas. Celulele au 2-3 flageli peritrichi. Se gasesc in nodozitatile radacinilor de Melilotus, Medicago si Trigonella.
Rhizobium lupini este formata din celule care se deplaseaza cu ajutorul a 2-3 flageli. Se gaseste in sol si in nodozitatile radacinilor de Serradella, Lupinus si Ornithopus.
La Rhizobium loti bacteroizii din noduli au tot aspect de bastonas, celulele au un flagel polar. Formeaza nodozitati la urmatoarele plante: Lotus corniculatus, Lotus tenuis, Lupinus densiflorus, Anthyllis vulneralia, Mimosa etc.
II.2.2. SPECIFICITATEA DE GAZDA A RIZOBIILOR.
Primul cercetator care a vorbit despre rizobiile cu crestere rapida din nodulii prezenti la leguminoasele erbacee si lemnoase a fost Trinick. De atunci, chiar daca simbiontii cu crestere lenta par sa fie mai numerosi in majoritatea ecosistemelor tropicale, tipurile cu crestere rapida au fost semnalate frecvent, si uneori chiar le domina pe celelalte.
Persistenta rizobiilor cu crestere rapida pare sa depinda de speciile legumicole prezente si de conditiile ecologice ale arealului. Arealele unde tipul cu crestere rapida domina pare sa fie cu precadere mediul arid, cu soluri sarace, din Africa, Australia si America de Nord. Speciile de Acacia sunt deseori dominante in areale ca Africa si Australia, si se pare ca acest gen este nodulat de rhizobii cu crestere rapida. Este dificil de spus daca dominanta tipului cu crestere rapida se datoreaza mai mult conditiilor de mediu decat preferintei pentru gazde.
Unii autori sustin fapul ca, anumite conditii adverse de mediu ar favoriza bacteriile cu crestere rapida intrucat acestea pot raspunde rapid fluctuatiilor conditiilor de mediu. Rizobiile cu crestere rapida par a fi, in general, capabile sa utilizeze un grad mare de surse de carbon, comparativ cu bradyrhizobiile. Tipul cu crestere rapida pare sa fie de asemenea mai tolerant la temperaturi mari si la seceta decat tipul cu crestere lenta.
S-a observat rezistenta crescuta a izolatelor rizobiene la antibiotice, posibil rezultat al adaptarii la mediul desertic, unde actinomicetele producatoare de antibiotice sunt prezente in numar mare. In concordanta cu acest studiu, intr-un mediu desertic, rata cresterii rapide, producerea de acizi, toleranta la temperaturi ridicate si rezistenta crescuta la antibiotice par sa fie caracterele ideale.
Studiind legaturile dintre conditiile de mediu si abundenta rizobiilor, s-a ajuns la ipoteza ca, prezenta unei anumite populatii de rizobii este corelata cu:
prezenta unei leguminoase - gazda potrivita;
precipitatiile anuale scazute;
surse de legume si biomasa de lastari;
temperatura solului.
Influenta plantei gazda in evolutia rizobiei este evidenta, dar nu se stie cat de importanta.
Coevolutia planta-rizobie a fost studiata de unii cercetatori care au izolat rizobii nodulante la fasole, din diferite regiuni ale globului. Pe parcursul acestor studii ei au intalnit cazuri de coevolutie datorita izolarii geografice. Spre exemplu, au gasit in sudul Turciei un soi de fasole care forma o simbioza eficienta cu tulpini locale de rizobii, dar nu si cu alte tulpini din alte parti ale Turciei. De asemenea, este relativ usor sa gasesti un centru de diversitate genetica presupus a fi centru de origine pentru plantele leguminoase. Posibil ca aceste areale sa fie aceleasi cu zonele in care rizobiile au coevoluat cu leguminoasele.
Studiile filogenetice bazate pe secventele 16S au aratat ca rizobiile cu crestere rapida sunt mult mai diverse din punct de vedere genetic decat cele cu crestere lenta, fapt reflectat de altfel si de divizarea lor in patru genuri diferite.
Noile specii din America, Rhizobium tropicii si Rhizobium etli au fost considerati simbionti primari pentru fasole, desi pentru Rhizobium tropicii - Leucaena leucocephala era cunoscuta drept gazda inca de la inceput. Mai tarziu s-a observat ca ambele specii noduleaza eficient un numar mare de leguminoase erbacee si lemnoase.
Rizobiile din Africa au fost testate pana acum pe cateva plante erbacee, desi unele dintre tulpini au fost capabile sa noduleze si alte specii precum Lotus cornicatus si Medicago sativa.
Specia Sinorhizobium este o tulpina cu un numar mare de gazde, care a fost timp indelungat studiata. S-a izolat din Lablab purpurens din Noua Guinee, si de atunci a fost testata pe 375 specii de leguminoase, noduland doar 205 dintre ele.
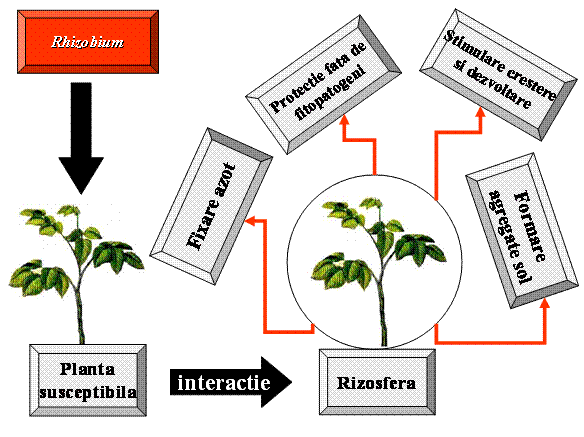 Fig.1. Actiunea
complexa a inoculantilor pe baza de rizobii fixatoare de azot
Fig.1. Actiunea
complexa a inoculantilor pe baza de rizobii fixatoare de azot
II.2.3. EFECTELE INOCULARII CU RHIZOBIUM - FORMAREA NODOZITATILOR.
Formarea nodozitatilor radiculare si manifestarea capacitatii de fixare a azotului atmosferic este rezultatul unei interactiuni reusite intre bacterie si planta gazda, ca urmare a parcurgerii unor etape succesive si obligatorii.
Nodozitatile fixatoare de azot atmosferic la plantele leguminoase se
formeaza pe radacini, fiind un rezultat al unei cresteri
tisulare anormale dar limitate, cu organizare armonioasa si
diferentiata. In interiorul nodozitatii, bacteroizii
fixeaza azotul molecular care este apoi utilizat de catre
planta-gazda (Mihaescu & Gavrila
Acest proces este rezultatul a patru stadii fiziologice distincte: preinfectia, infectia si formarea nodozitatii, functionarea nodozitatii, imbatranirea.
Etapele initiale ale procesului de infectie constau in: colonizarea radiculara, atasarea la radacini, rasucirea perisorilor sugatori si formarea filamentului de infectie (Robertson si col.,1981).
In stadiul de preinfectie, celulele rizobiene libere migreaza spre rizosfera plantei gazda datorita excretarii, de catre radacina, a unei game variate de compusi. Dupa ajungerea in rizosfera, are loc recunoasterea intre gazda si bacterie, proces ce implica transmiterea unor semnale mutuale specifice pentru activarea si transcrierea genelor implicate in simbioza (Schell si col., 1988).
Atasarea bacteriilor la radacini are loc in zone speciale de infectie, care corespund regiunii perisorilor absorbanti tineri. Formarea filamentului de infectie in celulele perisorilor absorbanti e o etapa specifica si se produce ca urmare a unui proces de degradare a peretilor celulari radiculari.
Celulele gazdei reactioneaza prin formarea unor structuri tubulare, asa numitele filamente de infectie, care inchid bacteriile aflate in diviziune.
Filamentele de infectie se dezvolta in directia centrilor meristematici de inductie din cortexul radicular si formeaza ramuri. In contact cu un centru meristematic, rizobiile sunt eliberate in citoplasma celulelor gazdei. In timpul eliberarii, bacteriile se inconjoara cu o membrana peribacterioida, care provine din plasmalema plantei. Bacteriile se diferentiaza in bacteroizi, in care este indusa exprimarea complexului enzimatic reducator de azot.
Bacteriile eliberate intracelular sufera modificari morfologice importante care variaza cu specia plantei, conditiile de infectie si crestere, varsta si dimensiunea nodozitatilor, si ele reprezinta sediul fixarii azotului atmosferic. In mod caracteristic, bacteroizii contin leghemoglobina care protejeaza sistemul enzimatic. Bacteroizii contin tot sistemul enzimatic necesar pentru reducerea azotului atmosferic la azot asimilabil plantelor. Pentru ca toate reactiile de transformare si asimilare sa aiba loc, planta trebuie sa transfere catre nodozitati substraturi bogate in carbon. De asemenea, e necesara difuzia oxigenului in cantitati suficiente.
Tabel 2. Stadiile de dezvoltare a nodozitatilor, etapele parcurse si interactiile dintre rizobii si planta gazda.
|
Stadiul de dezvoltare a nodozitatii |
Etape |
Interactiuni |
|
Preinfectia |
-migratie - atasare -curbarea perilor radiculari - inducerea inhibarii sistemului de aparare a meristemului plantei. |
- chemotaxie specifica - producere de factori de crestere difuzabili. |
|
Infectia si formarea nodozitatii |
-formarea filamentului de infectie - initierea si dezvoltarea nodozitatii. -eliberarea bacteriilor - diferentierea bacteroizilor. |
flavonoid - nodD - limitare de oxigen - exprimarea genelor nod, hns - exprimarea nodulinelor timpurii. |
|
Functionarea nodozitatii |
-fixarea de azot -functionarea complementara -persistenta functionarii nodozitatii. |
-oxigen - limitare - nifA -exprimarea genelor nif si fix -exprimarea nodulinelor timpurii si tarzii. |
|
Imbatranirea nodozitatii |
-dezintegrarea membranei - dezintegrarea celulei - distrugerea nodozitatii. |
II.2.3.1. STRUCTURA NODOZITATILOR.
Intr-o sectiune longitudinala prin nodozitate, zona cea mai indepartata este meristemul apical. Imediat langa acesta este asa numita zona de invazie, alcatuita din celule mari, ce vor fi infectate de rizobii.
In zona simbiozei incipiente celulele sunt diferentiate in infectate si neinfectate. Dupa zona incipienta urmeaza o zona de simbioza activa, unde are loc reducerea si asimilarea azotului.
Regiunea proximala a nodozitatii este zona senescenta, unde apare degenerarea celulei vegetale si a bacteroizilor. Cortexul nodozitatilor inconjoara zonele centrale ale acestora. Cortexul nodozitatii e divizat de un endoderm periferic, intr-un cortex interior si unul exterior. In corexul interior sunt localizate ramificatiile vasculare, acesta fiind separat de regiunile centrale ale nodozitatilor printr-un perete de celule distincte, mici, neinfectate.
II.2.3.2. FUNCTIONAREA NODOZITATILOR.
Sistemul enzimatic nitrogenazic este format din doua componente proteice diferite, care functioneaza asociat intr-un ciclu catalitic. Sinteza sistemului nitrogenazic este guvernata de genele nif.
Morfologia nodozitatii radacinii este atat sub controlul plantei gazda, cat si a bacteriei. Adaptarile morfologice si biochimice observate la simbioza lui Rhizobium cu leguminoasele sunt atribuite proprietatilor genetice si biochimice ale ambelor organisme.
Dintre plasmidele pe care le poseda bacteriile din genul Rhizobium, cele denumite Sym poarta majoritatea genelor implicate in specificitatea de gazda (hsn), in nodulare (nod), si in fixarea azotului (fix/nif). Mai exista si alte gene cromozomale sau plasmidiale, care joaca un rol in simbioza, spre exemplu genele ce codifica sinteza exopolizaharidelor.
Proprietatile genetice ale plantelor gazda determina dezvoltarea nodozitatilor. Exprimarea genelor plantelor difera in nodozitati fata de radacinile neinfectate.
In stadiul de preinfectie, rizobiile interactioneaza cu genomul plantei, prin eliminare de factori de crestere difuzibili - fitohormoni, inducand centri meristematici in cortexul radicular (Schell si col. 1988).
Filamentele de infectie sunt formate de planta gazda, ceea ce indica exprimarea anumitor gene. Produsii acestora sunt numiti noduline timpurii si pe baza compozitiei lor in aminoacizi se pare ca sunt proteine ale peretelui celular (Gloudemans si col., 1998).
Desi plantele au numeroase mecanisme de aparare fata de patogeni, la simbioza leguminoaselor cu rizobiile, absenta reactiilor defensive pe parcursul dezvoltarii nodozitatilor indica o represie a genelor implicate in aceste procese.
Dintre factorii care regleaza functionarea nodozitatilor fac parte oxigenul, haem-ul si amoniul. Pentru exprimarea genelor nif si fix este necesara o concentratie scazuta de oxigen, obligatorie si pentru activitatea nitrogenazica. Haem-ul este o componenta esentiala a leghemoglobinelor care manifesta o mare afinitate pentru oxigen si functioneaza ca purtator de oxigen in nodozitati. Amoniul este produsul actiunii nitrogenazice, se pare ca formarea sa determina cresterea ph-ului si are drept consecinta activarea unor enzime.
II.2.4. FENOMENUL DE QUORUM SENSING LA BACTERIILE FIXATOARE DE AZOT DIN GRUPUL RHIZOBIA.
Quorum sensing (QS) este unul dintre mecanismele de comunicare intercelulara bacteriana cel mai intens studiat, termenul de QS fiind introdus de catre Fuqua si col. in 1994, ca fiind un proces de detectare a densitatii celulare.
Quorum sensing (QS) este un mecanism prin care diferite microorganisme controleaza procese specifice ca raspuns la densitatea populatiilor. Acest sistem de comunicare celula la celula se realizeaza pe calea producerii unor molecule specifice cunoscute sub numele generic de molecule semnal sau autoinductori.
Populatia este astfel capabila sa-si modifice comportamentul ca o structura unitara care reactioneaza la un prag critic de densitate cunoscuta sub numele de qvorat (Fray, 2002). La bacteriile Gram negative moleculele semnal sunt N-acil homoserin lactonele (AHL) care difera intre ele prin structura lanturilor laterale.
Mecanismul QS regleaza sporularea, sinteza de antibiotice, induce factorul de virulenta, diferentierea celulara si fluxul de nutrienti, competenta patogenica etc. La densitati mici ale populatiilor se produce un nivel scazut de AHL. Cand densitatea celulelor creste, avand tendinta de a forma un film biologic invaziv, AHL-ul produs in cantitate mare se acumuleaza in mediu si, cand se atinge concentratia critica, difuzeaza in celule legandu-se la receptorul specific fapt care duce la activarea sau represia exprimarii genelor care determina competenta ecologica a bacteriilor.
Dat fiind rolul important pe care acest fenomen il are in interactiile patogene (Loh si col., 2002), inactivarea sau degradarea moleculelor AHL constituie o cale inovativa de modulare a actiunii agroinoculantilor.
Fenomenul de QS include producerea, emiterea si detectarea a unor mici molecule de semnal, numite "molecule de autoinducere". Aceste molecule se acumuleaza in mediu odata cu cresterea densitatii celulare.
Atunci cand aceste molecule semnal depasesc o concentratia critica, bacteriile le detecteaza, si raspund la ele printr-o expresie genetica sincronizata. Prin utilizarea acestor sisteme de semnal-raspuns, bacteriile pot sa-si regleze diverse activitatii, care sunt eficiente doar daca bacteriile actioneaza simultan.
Procese controlate prin QS includ bioluminescenta, virulenta, formare de biofilme, schimb de ADN sau formarea de spori (Bejerano-Sagie si Xavier, 2007).
Avand in vedere rolul important pe care il are fenomenul quorum sensing in comunicarea intra- si extraspecifica la bacterii, capacitatea de a impiedica producerea sau detectarea moleculelor de semnal ar fii in folosul organismelor gazde sau a bacteriilor concurente. In nisele in care populatiile de bacterii concureaza pentru resursele limitate, capacitatea unei specii de a impiedica comunicarea prin QS la specia cealalta, constituie un avantaj. De asemenea, capacitatea gazdei de a interfera cu comunicarea intercelulara bacteriana poate fii cruciala in prevenirea colonizarii de catre bacteria patogena, care utilizeaza quorum sensing-ul pentru coordonarea virulentei. Acele mecanisme care s-au dezvoltat pentru a interfera cu comunicarea intercelulara bacteriana au fost numite "quorum quenching".
Probabil exista si mecanisme analoage, menite sa promoveze o functie (un comportament) controlat prin quorum sensing, atunci cand aceast comportament aduce beneficii acelor organisme care cohabiteaza cu bacteriile care utileaza QS-ul. Insa despre aceste mecanisme se stie foarte putin la ora actuala Waters si Bassler, 2006).
Speciile din grupul Rhizobia sunt cunoscute datorita faptului ca ele pot avea relatii simbiotice cu plantele leguminoase, in urma carora la radacinile plantei se formeaza nodozitatii fixatoare de azot.
La cateva specii, langa bine cunoscutele molecule de semnal (flavonoide, factori Nod si exopolizaharide) implicate in formarea nodozitatilor, s-a evidentiat fenomenul quorum sensing bazat pe molecule de semnale de tip AHL, aceasta fiind legata de fenomene variate, ca eficienta formarii nodozitatilor, dezvoltarea "simbiozomelor", producerea de exopolizaharide si fixarea azotului. Toate aceste fenomene sunt importante in realizarea cu succes a simbiozei.
Relatia simbiotica care se formeaza intre bacteriile fixatoare de azot din grupul Rhizobia si plantele gazde (leguminoase) este rezultatul unor retele complexe de semnalizare intre gazda si organismul simbiont. Pana astazi, multe aspecte ale schimbului de semnale sunt necunoscute, insa se pare ca mecanismul QS joaca un rol cheie in procesul simbiotic, deoarece acest proces depinde de concentratia bacteriilor din jurul si din interiorul radacinii plantelor si a nodozitatilor. Cresterea densitatii celulelor bacteriene Rhizobia, determinata de procesul QS, este un component important in procesul de semnalizare.
Dintre bacteriile fixatoare de azot, fenomenul QS este cel mai bine studiat la specia Rhizobium leguminosarum. Au fost identificate nu mai putin de patru sisteme quorum sensing (rai, rhi, cin, si tra) cu 6 tipuri de proteine receptoare de tip LuxR (raiR, rhiR, cinR, traR, bisR, expR). Impreuna, aceste sisteme regleaza genele implicate in formarea de biofilme, conjugatie, simbioza, fixarea azotului (Waters si Bassler 2006, Pappas si col. 2004, Whitahead si col. 2001).
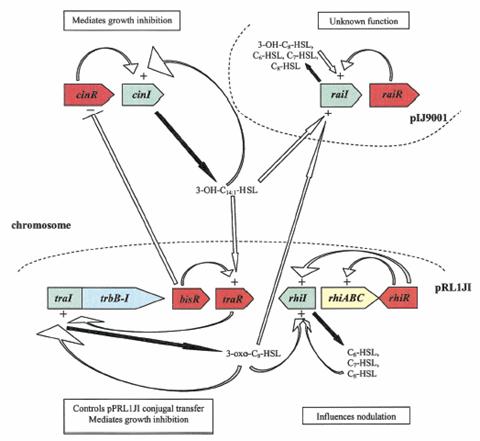
Fig.2. Retea quorum sensing la Rhizobium leguminosarum
(Gonzles si Marketon 2003, pp. 582).
Sistemul quorum sensing la bacteria R. etli este mai putin cunoscuta, dar si mai putin complex, decat cel al speciei R. leguminosarum. Desi la R. etli au fost gasite sapte molecule de AHL diferite, doar doua sisteme de quorum sensing au fost identificate: sitemele raiRI si cinRI, care sunt localizate pe cromozoma. Aceste doua siteme sunt responsabile de sintetizarea tuturor moleculelor de AHL. Locusul cinRI codifica o molecula de AHL cu catena lunga (a carei structura este inca necunoscuta), care are rol in inhibarea cresterii bacteriene.
Locusul cinRI este responsabil de fixarea eficienta a azotului si de formarea simbiozei. Sistemul raiRI este reglat pozitiv de moleculele AHL produse de genele cinI, si la randul lor genele raiI (omoloage cu genele luxI) sunt responsabile de sintetizarea a catorva molecule AHL cu catene scurte (neidentificate inca). Locusul raiRI are rol si in controlarea fixarii azotului si in inhibarea cresterii (Gonzles si Marketon 2003).
II.3. GENUL AZOSPIRILLUM
Prima specie de Azospirillum a fost izolata de Beijerinck (1925) in Olanda, dintr-un sol nisipos sarac in azot si a fost denumita initial Spirillum lipoferum. Ulterior, bacteria a fost izolata din unele alge marine si din filosfera unor plante tropicale. Dbereiner si Day (1976) au relatat larga distributie in rizosfera multor buruieni tropicale. Dupa aceasta, bacteria a fost izolata de la radacinile a numeroase plante salbatice si cultivate, cereale si leguminoase din soluri tropicale, subtropicale si temperate.
Tarrand si col. (1978) a propus divizarea in doua specii distincte: A.brasilense si A.lipoferum, pe baza diferentelor morfologice si fiziologice intre diferite tulpini pe baza experimentelor de omologie a ADN. Aceste 2 specii au fost primele descrise, iar in urma cercetarilor ulterioare s-au identificat ca specii distincte A. amazonese, A. halopraeferans, A. irakenses, A. largimobile. Aceasta din urma a fost mult timp considerata o specie sinonima cu A. lipoferum. Recent, ea a fost denumita A. doebereinerae, in onoarea celui care a deschis studiul acestui gen bacterian.
Ca aspect, speciile de Azospirillum sunt bastonase mai
groase, curbate, cu marimea intre 0,8-1,0 x 25 μm, cu granule de PHB,
care in celulele fixatoare de N![]() pot
sa atinga 50% din masa uscata a celulei. Normal celulele sunt
Gram negative, insa la Azospirillum
brasiliense s-a observat o variabilitate in ceea ce priveste
caracterul Gram. Celulele sunt mobile, cu un singur flagel polar.
pot
sa atinga 50% din masa uscata a celulei. Normal celulele sunt
Gram negative, insa la Azospirillum
brasiliense s-a observat o variabilitate in ceea ce priveste
caracterul Gram. Celulele sunt mobile, cu un singur flagel polar.
Dupa 1993, bacteria s-a bucurat de o atentie deosebita datorita capacitatii sale de stimulare a cresterii plantelor.
 Fig.3. Azospirillum
brasiliense
Fig.3. Azospirillum
brasiliense
II.3.1. SPECIFICITATE SI VARIABILITATE
Una din cele mai controversate probleme referitoare la asocierea cu Azospirillum se refera la specificitatea planta - tulpina bacteriana si modul cum aceasta afecteaza cresterea plantelor. Diferente specifice s-au inregistrat intre raspunsurile plantelor C3 si C4. Azospirillim lipoferum a fost specia predominanta care colonizeaza plantele C4, iar Azospirillum brasiliense a fost specia precominant asociata cu plantele C3 in zonele tropicale.
Plantele C3 si C4 sunt impartite pe baza tipului de fotosinteza pe care o realizeaza. Plantele ce corespund tipului C3 reprezinta circa 300000 de specii raspandite in zonele cu climat termperat; la acestea produsii intermediari ai fotosintezei au 3 atomi de carbon. Tipul fotosintetic C4 se intalneste la aproximatic 1000 de specii originare in zonele tropicale, adaptate la conditii de iluminare intensa si la temperatura ridicata. Produsii intermediari de carbon formati in timpul fotosintezei au 4 atomi de carbon. Exemple: Zea mays, Sorghum vulgare, Beta vulgaris, etc.
O preferinta similara a bacteriilor din genul Azospirillum pentru plantele gazda a fost observata si in zonele temperate.
Principalele dificultati in determinarea specificitatii bacteriei sunt: lipsa testelor de comparatie pentru tulpini obtinute din diferite surse, experimentate pe anumite plante gazda si numarul limitat de tulpini testate pana in prezent.
Exista studii ce indica faptul ca specificitatea nu este la nivel de specie ci la nivel de tulpina. O selectie rapida pentru determinarea asocierii planta - tulpina bacteriana este esentiala pentru prognoza succesului combinatiilor inainte de testarea in camp.
II.3.2. EFECTELE INOCULARII CU AZOSPIRILLUM LA PLANTE
Inocularea cu Azospirillum poate avea ca rezultat o modificare semnificativa a unor parametri de crestere ai plantelor, fapt care poate sau nu sa afecteze recolta. Majoritatea studiilor au fost realizate cu cereale, si doar foarte putine cu plante din alte familii. Cele mai evidente raspunsuri observate au fost: cresteri ale greutatii uscate totale a plantelor, cresteri ale cantitatii de azot din tulpini si seminte, cresterea numarului de frati, inspicare si inflorire timpurie, cresterea numarului de spice si a numarului de seminte/spic, cresterea greutatii semintelor, cresterea inaltimii plantelor si a marimii frunzelor, marirea ratei de germinatie, o crestere marcanta a sistemului radicular, respectiv a lungimii si volumului radacinilor.
Din solurile aride din Iran au fost izolate si identificate 10 tulpini de Azospirillum care, in conditii de sera, cu sol steril, au determinat cresterea inaltimii, a greutatii uscate a tulpinii si radacinii, ramificarea radacinii si, in general au imbunatatit sistemul radicular al hibrizilor de porumb si soiurilor de grau cu care s-a lucrat. Majoritatea tulpinilor native au avut efecte mai bune asupra parametrilor biologici ai plantelor inoculate decat tulpinile straine utilizate ca standard (Rousta si col., 1998).
Efectul inocularii cu Azospirillum asupra cresterii totale a recoltei plantelor cultivate in camp a fost, in general, de 10-30%, dar in cateva comunicari sunt date si valori extrem de mari, de 50-70%, in comparatie cu martorii neinoculati. Chiar cresterile moderate, de pana la 20% sunt considerate ca avand valoare comerciala pentru agricultura moderna, in cazul care se obtin in mod constant.
Efecte negative sau lipsa de efect a rezultatelor inocularii au fost rareori relatate (Albrecht si col.,1 981; Harris si col., 1989; Smith si col., 1984).
In 1985, Okon a evaluat rezultatul inocularii lui Azospirillum in experientele efectuate in toata lumea si a tras concluzia ca, in circa 65% din toate experientele, rezultatele au fost pozitive. Cresteri de recolta datorate inocularii cu Azospirillum au fost relatate in 75% din experimentele cu cereale, bumbac si legume (Bashan si col., 1989).
Semintele de orez germinate, tratate cu Azospirillum au generat plante mai inalte si mai viguroase decat cele provenite din seminte netratate, au avut o rata de supravietuire la 100C in aer superioara si o coacere cu 3-5 zile mai timpurie (Chi si col., 1998).
Efectul tratamentului la Anacardia occidentale, in pepiniere cu Azospirillum, in momentul semanatului, comparativ cu plantele neinoculate si cu tratamentele cu alti biofertilizatori, au pus in evidenta un efect de stimulare a rasaririi si a cresterii dar si de scadere a incidentei bolilor fungice (Kumar si col., 1998).
Rezultate diferentiate au fost obtinute in ceea ce priveste conditiile administrarii biopreparatului in raport cu nivelul fertilizarii cu azot. Astfel, Kapulnik si col.(1981), Lau-Wong (1987), Mertens si Hess (1984), O'Hara si col. (1987) au considerat inocularea cu Azospirillum ca un substitut partial pentru fertilizarea cu azot, rezultatele cele mai bune fiind obtinute in conditiile fertilizarii suboptimale. Datele lui Bashan si col. (1989) sau ale lui Millet si Feldman (1986) arata ca recolta a crescut si la inocularea in conditiile fertilizarii cu niveluri mari de azot care, dupa Kolb si Martin (1988) poate influenta numarul de bacterii din rizosfera.
In conditii tropicale, aplicarea tulpinii A.brasilense la cultura de batat, pe sol nisipos, pentru inlocuirea fertilizarii cu azot a aratat ca plantele au produs o recolta mai mare, au avut o crestere vegetativa mai viguroasa si un continut de azot mai mare in radacini si frunze decat cele neinoculate (Saad si col, 1998). In acelasi experiment s-a pus in evidenta necesitatea stabilirii compatibilitatii soi-tulpina bacteriana, precum si a aplicarii initiale de ingrasamant cu azot la 1/3 sau mai putin din doza recomandata in scopul asigurarii necesarului de azot pana la inceperea activitatii asocierii planta-bacterie.
Una dintre problemele majore ale experimentarilor, o constituie identificarea caii de inoculare care sa aibe rezultate favorabile, constante si repetabile, de aceea impunandu-se clarificarea: modului de actiune al bacteriei ca favorizant de crestere al plantelor; interactiilor si competitiei lui Azospirillum cu alte microorganisme din rizosfera; modului cum sunt influentate supravietuirea si activitatea bacteriei in rizosfera de catre diferiti factori din mediu.
II.3.2.1. MODUL DE ACTIUNE ASUPRA CRESTERII PLANTELOR
Sunt implicate mai multe moduri de actiune printre care:
fixarea azotului contribuind la aprovizionarea cu azot a plantei;
efecte hormonale, care altereaza metabolismul si cresterea plantei;
imbunatatirea cresterii sistemului radicular avand ca rezultat imbunatatirea aportului de nutrienti si apa;
activitatea nitrat reductazica care mareste acumularea de azotat in plantele inoculate.
II.3.2.2. EFECTELE HORMONALE ALE BACTERIEI ASUPRA CRESTERII PLANTELOR
Numeroase tulpini de Azospirillum produc hormoni vegetali in cultura lichida. Principalul hormon produs este acidul indolil-3-acetic (AIA). Alti hormoni, detectati in cantitati mult mai mici dar la niveluri semnificative biologic au fost acidul indollactic, indol-3-butiric, indol-3-etanol, indol-3-metanol, compusi indolici neidentificati, mai multe gibereline, acid abscizic si citokinine.
Hormonii vegetali afecteaza capacitatea de fixare a azotului de catre Azospirillum. La plantule, aplicarea externa de hormoni, sintetici sau purificati de la cultura bacteriana a reprodus in totalitate efectele bacteriei asupra dezvoltarii si morfologiei radacinii. In mod special s-au observat modificari ale lungimii radacini, producerea mai multor peri radiculari si ramificarea perilor radiculari, producerea mai multor radacini laterale, marirea ratei de diviziune celulara si deferentiere in tesuturile meristemice.
Inocularea cu Azospirillum a imbunatatit echilibrul hormonal al unei mutante de gr1u, defectiva hormonal. Cantitati mai mari de IAA si IBA au fost identificate in radacinile de porumb inoculate comparativ cu plantele neinoculate. Aceste rezultate sugereaza implicarea indirecta a lui Azospirillum in reglarea hormonala a plantei.
II.3.2.3. FIXAREA AZOTULUI
Toate tulpinile salbatice de Azospirillum fixeaza cu eficacitate azotul atmosferic fie ca bacterii libere, fie in asociere cu plantele si iau parte la unele reactii din ciclul azotului. Ca urmare a inocularii apare o crestere a azotului total in lastarii i semintele plantelor inoculate. Prin urmare, fixarea de azot este primul mecanism important de actiune care imbunatateste cresterea plantelor.
Incorporarea de azot atmosferic in planta gazda a fost evaluata prin testul de reducere al acetilenei si prin utilizarea izotopului 15N2. Dovada ca fixarea azotului contribuie la echilibrul azotului in plante se bazeaza pe observatia banala a cresterii activitatii nitrogenazice in radacinile inoculate (Rao si Rajaamamohan Rao, 1983; Yahalom si col., 1984).
Aceasta activitate enzimatica este suficient de mare a fi luata in considerare ca responsabila de cresterea azotului total in recolta plantelor inoculate, daca se considera ca tot azotul fixat este incorporat in plante (Sarig si col.,1984; Mertens si Hess, 1984). De asemenea, s-a aratat ca niveluri mici sau chiar neglijabile de activitate nitrogenazica au aparut in plante care au raspuns pozitiv la inoculare (Venktewarlu si Rao, 1983). Cantitatea de azot fixata determinata nu explica cresterea continutului de azot total din plantele inoculate.
II.3.2.4. IMBUNATATIREA DEZVOLTARII RADACINII
Pe langa cresterea sau scaderea multor parametri radiculari, inocularea plantelor cu Azospirillum afecteaza numerosi parametri foliari. Aceste modificari au fost atribuite direct efectelor bacteriene pozitive asupra absorbtiei de substante minerale de catre planta: activitatea nitrat reductazica care mareste acumularea de azotat in plantele inoculate.
Absorbtia marita de substante minerale se crede ca ar fi datorata unei mariri generale de volum a sistemului radicular si nu unui mecanism specific de crestere a absorbtiei de ioni. S-a sugerat ca inocularea cu Azospirillum poate favoriza posibilitatea ionilor de a deveni accesibili in sol, ajutand planta sa evite limitarea nutrientilor. Acest lucru poate explica acumularea de compusi cu azot in planta fara vreo fixare aparenta de azot. Planta poate absorbi azotul mai eficient de la sursele limitate din sol, avand ca rezultat, pentru a atinge o anumita recolta, o micsorare a cerintelor de fertilizare cu azot. Dovada care sustine cresterea absorbtiei minerale de catre radacinile inoculate e furnizata de cresterea activitatii de eflux protonic a radacinilor de grau inoculate cu Azospirillum.
In afara de absorbtia imbunatatita de substante minerale, inocularea cu Azospirillum a imbunatatit starea hidrica in plantele de sorg stresate. Plantele inoculate au fost mai putin stresate hidric, avand mai multa apa in foliaj, un potential hidric foliar mai mare si o temperatura a foliajului mai mica decat plantele neinoculate. Extractia totala de umiditate din sol de catre plantele inoculate cu Azospirillum a fost mai mare, iar apa a fost extrasa din apele mai adanci din sol. Prin urmare, recolta de sorg a crescut la plantele inoculate, fapt atribuit, in principal, utilizarii imbunatatite a umiditatii solului (Sarig si col., 1988). Este probabil ca absorbtia imbunatatita de substante minerale si apa are un rol vital in asocierea Azospirillum-planta.
II.3.2.5. EFECTELE INOCULARII ASUPRA DEZVOLTARII RADACINILOR
Cele mai importante efecte ale inocularii cu Azospirillum la plante sunt modificarile morfologice ale sistemului radicular. S-a constatat ca aceste modificari sunt legate de nivelul concentratiei inoculului, respectiv ca, depasirea nivelurilor optime are efect inhibitor, iar dozele suboptimale nu au efect. Nivelul optim stabilit pentru inocularea semintelor sau plantulelor multor cereale, legume si plante industriale este 105-106 ufc/ml, pentru porumb 107 ufc/ml, pentru tomate in vitro mai mult de 108 ufc/ml.
Efectele pozitive ale inocularii au fost demonstrate cu privire la:
- cresterea in lungime a radacinii, in special a zonei de elongatie;
- cresterea numarului si lungimii radacinilor laterale;
- cresterea numarului si densitatii, precum si aparitia timpurie a perilor radiculari;
- cresterea suprafetei radiculare;
- intensificarea diviziunii celulare in meristemul radicular;
- modificari ale aranjamentului celulelor in cortex;
- stimularea exudarii radiculare.
Ca urmare a studiilor efectuate, s-a emis ipoteza ca modificarile morfologice la nivelul sistemului radicular al plantelor in sensul cresterii volumului acestuia, duc in final, la cresterea recoltei.
II.3.2.6. COLONIZAREA RADACINILOR
Azospirillum este un colonizator atat extern cat si intern al radacinilor. In forma de colonizare externa, bacteria poate fi gasita sub forma de mici agregate, dar si ca celule izolate. Pot fii colonizate atat radacinile vii cat si cele moarte. La colonizarea interna, celulele bacteriene pot penetra in spatiile intercelulare radiculare. Studiile de microscopie electronica au aratat ca bacteriile de la suprafata radacinilor sunt conectate cu radacina si unele de altele, printr-o retea de material fibrilar, de natura proteica.
Atasarea prin fibrile de natura bacteriana depinde de metabolismul bacteriei si explica de ce colonizarea este realizata numai de celulele vii. Atasarea lui Azospirillum la suprafata radacinii apare rapid, in cateva secunde sau minute de expunere a materialului vegetal. Majoritatea zonelor colonizate devin saturate in doua ore dupa inoculare, in functie de tulpina si faza de crestere a bacteriei (Eyers si col, 1988). Unul dintre mecanismele sugerate ca fiind implicate in legarea bacteriilor la radacini il constituie legarea prin lectine, presupunandu-se ca aglutininele pot fi localizate in materialul fibrilar si ajuta la ancorarea celulelor.
Modul de colonizare al radacinii poate sa varieze in functie de tulpina de Azospirillum, specia de planta, conditiile de mediu si alti factori. Interactia intre aceste variabile creaza diferite grade si modele de colonizare, diferite marimi ale populatiei si diferite locuri de colonizare.
CAPITOLUL III. MATERIAL SI METODE
III.1. IZOLAREA UNOR TULPINI BACTERIENE DIN GRUPUL RHIZOBIUM
Grupul Rhizobium face parte din familia Rhizobiaceae. Acesta cuprinde celule fara endospori, de forma bacilara, in stadiile tinere. Sunt microorganisme aerobe gram negative, utilizeaza numeroase substante hidrocarbonate, si produc, in cursul cresterii, un bogat exudat polizaharidic. Toate speciile incita hipertrofii corticale, tulpinale sau radiculare, fapt care constituie specificitatea de gen. Toate speciile fixeaza azotul liber, nu utilizeaza citratul si nu produc 3-ketolactaza, caracteristici care au fost utilizate pentru a deosebi genul Rhizobium de genul Agrobacterium. Bacteriile cresc bine pe medii lipsite de surse de azot. Grupul Rhizobium a fost initia clasificat in functie de ritmul de crestere pe mediul cu extract de drojdii in doua genuri: genul Rhizobium (ex. Rhizobium leguminosarum), cu crestere rapida si flageli peritrichi si genul Bradyrhizobium (ex. Bradyrhizobium japonicum), cu crestere lenta si un flagel polar sau subpolar.
Studiile de biologie moleculara au determinat reclasificarea grupului Rhizobium si aparitia unor noi genuri: Azorhizobium, Mesorhizobium, Ensifer / Sinorhizobium. La modul general continua sa se foloseasca pentru toate aceste genuri apelativul comun de rhizobii (casi denumirea generica de Rhizzobium), delimitand speciile de procariote care, in regiunile temperate, formeaza in mod obisnuit nodozitati la plantele leguminoase. Din aceste nodozitati bacteriile pot fi izolate.
Metodologia de izolare a speciilor de Rhizobium se orienteaza in primul rand dupa obiectul de la care se porneste - solul sau nodozitatile radiculare. In cazul izolarii din sol, se prepara un extract de sol dintr-o suspensie de 1:1 000. Un mediu nutritiv eficient pentru Rhizobium se suplimenteaza cu o solutie de cristal violet foarte diluata (1: 80 000), in vederea inhibarii dezvoltarii bacteriilor Gram pozitive si a actinomicetelor. Mediul nutritiv se repartizeaza in tuburi cu capilar subtire care, ulterior, se introduc in extractul de sol. Speciile de Rhizobium se retrag in tuburile capilare. Din masa de Rhizobium astfel obtinuta, se inoculeaza o anumita cantitate pe mediul nutritiv suplimentat cu cristal violet. Prin inoculari repetate si analizate la microscop, se obtin, in final, culturi pure de Rhizobium.
Prelevarea probelor de nodozitati trebuie sa se efectueze de la plante pe cat posibil sanatoase, de pe terenuri netulburate. Este esentiala descrierea si marcarea locului din care se ridica materialul biologic ce urmeaza sa fie analizat. Numarul nodozitatilor necesare variaza in functie de scopul prelevarii. In general, sunt suficiente 15-20 nodozitati/planta culese din regiunea de varf a sistemului radicular principal. Prelevarea nodulilor poate varia in functie de specie astfel ca, speciile stolonifere pot sa aiba nodozitati dispuse pe radacinile adventive situate la 1-2 cm de la suprafata, iar la speciile de plante lemnoase, nodozitatile pot fi asezate in sol la adancimi mai mari, la o anumita distanta de la trunchiul copacului.
La experimentele de izolare pe care le-am realizat, s-au folosit nodozitati provenite de pe radacinile unor specii de plante leguminoase, au fost dezinfectate prin imersie in acool etilic 60%, timp de 2 minute. Apoi au fost imersate in cloramina 1%, dupa care s-au spalat de trei ori in apa distilata sterila.
Dupa dezinfectare, nodozitatile au fost mojarate in cateva picaturi de ser fiziologic steril. Din suspensia obtinuta in urma mojararii se ia o incarcatura microbiana si se insamanteaza pe placi Petri cu mediul Lazareva agarizat, prin tehnica de epuizare a ansei. Coloniile izolate au fost repicate de 3 ori pentru asigurarea puritatii izolatelor.
Componentele mediului de crestere Lazareva sunt urmatoarele:
- manitol - 10 g/l,
- K2HPO4 - 0,5 g/l,
- MgSO4x7H2O - 0,2 g/l,
- NaCl - 0,2 g/l,
- CaSO4 - 0,1 g/l,
- MnSO4 - 0,005 g/l,
- (NH4)2MoO4x 2H2O - 0,002 g/l
- extract de drojdii - 1 g/l,
- agar-agar - 20 g/l,
- 1000 ml apa distilata;
pH-ul mediului este 6,8; mediul se sterilizeaza prin autoclavare la 0.8 atm timp de 20 de minute.
III.1.1 URMARIREA DEZVOLTARII CULTURII IN MEDIUL LICHID
Izolatele din nodozitati, dupa purificare, au fost inoculate in flacoane cu mediu Lazareva lichid, si incubate la 30C timp de 7 zile, in conditii de aerare. Experienta a fost organizata in 3 repetitii, fiecare varianta cuprinzand trei flacoane/tulpina bacteriana.
La 4 si 7 zile s-au facut observatii asupra nivelului de crestere. Pentru aceasta, s-au centrifugat 30ml suspensie bacteriana timp de 15 minute la 5000 rpm. Supernatantul a fost indepartat, iar sedimentul - biomasa umeda a fost cantarit. Rezultatele au fost extrapolate la 1l de mediu.
III.1.2. ESTIMAREA NUMARULUI DE MICROORGANISME
S-a facut prin tehnica unitatilor formatoare de colonii. Probele prelevate aseptic au fost diluate serial (x 10), iar dilutiile 10-7 , 10-8, si 10-9 au fost inoculate pe placi Petri cu diametrul de 9 cm, continand mediul Lazareva agarizat. S-au numarat numai placile care au intre 30 si 100 colonii pe placa.
III.2. CARACTERIZAREA UNEI TULPINI DE RHIZOBIUM CU MICROSISTEMUL BIOLOG
Pentru identificarea taxonomica si caracterizarea tulpinilor microbiene izolate din diverse medii sunt realizate in prezent studii detaliate, folosindu-se biologia moleculara a acizilor nucleici (procent molar de guanina + citozina in ADN cromozomal, polimorfismul fragmentelor de restrictie pe ADN cromozomal, profilul plasmidial si tiparul polimorfic al fragmentelor rezultate in urma tratarii cu enzime de restrictie, amplificarea specifica a genei care codifica pentru ARN ribozomal de 16S si secventializarea acestei enzime etc.), alte tehnici chemotaxonomice (tiparul electroforetic al proteinelor totale sau al unor forme moleculare multiple ale enzimelor, profilul acizilor grasi membranari, profilul micosterolilor, spectrul de glicolipide membranare etc.) si testele caracterizare fenotipica.
Pana in deceniul sapte al secolului trecut clasificarea bacteriilor se realiza doar prin folosirea informatiilor fenotipice - caracteristicile morfologice, structurale, fiziologice si biochimice ale tulpinilor de microorganisme studiate. Incepand cu anii 80 ai secolului trecut au luat o amploare deosebita cercetarile de caracterizare a bacteriilor la nivel molecular, cristalizandu-se in final taxonomia moleculara, care se bazeaza in mod special pe informatii genotipice ( si in primul rand pe secventierea genelor care codifica molecule de 16s ARN ribozomal).
Informatiile genotipice nu sunt insa foarte relevante din punct de vedere al utilizarii practice, unde esentiale sunt caracteristicile fenotipice ale bacteriilor. Din acest motiv la jumatatea deceniului trecut s-a argumentat in favoarea unei abordari de tip integrator in ceea ce priveste identificarea si incadrarea taxonomica a tulpinilor bacteriene, descriindu-se taxonomia polifazica (Vandamme, 1996). In cadrul acestui sistem integrativ taxonii bacterieni sunt definiti pe baza informatiilor obtinute de diverse discipline, reunind astfel atat date fenotipice (cunoscute ca facand parte din taxonomia conventionala), cat si date de nivel molecular (date de biologia moleculara mentionate mai sus, dar si date derivate din abordarile moleculare integrative de tip -omica: transcriptomica, proteomica, metabolomica.
Pentru reducerea volumului considerabil de munca, materiale si timp necesar realizarii testele utilizate pentru caracterizarea fenotipica a microorganismelor au fost propuse o serie de micrometode integrate unor seturi standardizate, larg accesibile comercial (kit-uri microtest). S-a deschis astfel calea unei standardizarii internationale a testelor de fiziologie microbiana, astfel ca rezultatele obtinute sunt comparabile de la un laborator la altul. Pe plan mondial s-au impus 2 sisteme microtest pentru determinari fiziologice / biochimice necesare caracterizarii fenotipice a bacteriilor:
Sistemul API, produs de compania Biomerieux din Franta,a aparut pe piata in anul 1970, revolutionand lumea bacteriologiei. Sistemul API cuprinde benzi cu diferite teste biochimice si baze de date necesare identificarii si caracterizarii bacteriilor. Api cuprinde 15 sisteme de identificare acoperind practic toate grupele de bacterii si peste 600 de specii diferite.Testele biochimice de identificare si caracterizare se realizeaza pe baza reactiilor de culoare, ce apar ca urmare a metabolismului bacteriilor de testat ce se inoculeaza.
Kiturile Api sunt usor de folosit si interpretarea rezultatelor se face rapid,dupa 18-24 ore de la aplicarea suspensiei bacteriene pe benzi.
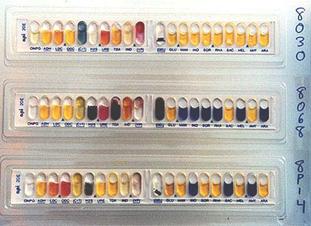
Fig.4. Benzi Api
- Sistemul Biolog, produs de compania americana Biolog Inc., a aparut pe piata in 1989. Sistemul utilizeaza microplaci de titrare cu 95 de godeuri in care exista diferite surse de carbon impreuna cu un colorant tetrazolic, care in urma exprimarii metabolismului bacterian formeaza un precipitat violet, generand astfel reactia de culoare. Exista diferite tipuri de microplaci: GN2, GN2-Ent, GP, YT, s.a.,iar tiparele de amprentare metabolica sunt comparate si identificate folosind softul bazei de date Microlog . Citirea prealabila a placilor se poate face cu un reader automat sau se poate realiza o citire manuala.
In cadrul proiectului, pentru caracterizarea fenotipica a tulpinilor bacteriene s-a folosit microstatia Biolog, existenta in dotarea laboratorului de microorganisme utile din cadrul ICDPP.
Protocolul de lucru a fost urmatorul:
S-a insamantat pe mediul special BUG (Biolog Universal Growth) repartizat in placi Petri, una din culturile izolate anterior si s-a lasat la incubat 16-24 ore la 30C.
S-a calibrat Biologul, apoi, din placa Petri s-a pelevat cu o ansa sterila de unica folosinta putina masa bacteriana si s-a omogenizat in lichidul de inoculare specific (pentru bacterii gram negative). S-a omogenizat masa bacteriana pana cand lichidul inoculat a atins o turbiditate standard, specifica de asemenea tipului gram negativ. Operatiunea s-a realizat cu atentie, si s-a verificat dupa fiecare omogenizare turbiditatea lichidului inoculat la Biolog. Dupa ce s-a atins turbiditatea corespunzatoare lichidul inoculat s-a turnat intr-o cuva sterila, de unde, cu o micropipeta s-a insamantat microplaca GN Biolog cu cate 150 l suspensie in fiecare godeu.
Placa se incubeaza 4-6 ore sau 16-24 ore si apoi se realizeaza citirea manuala sau automata, cu cititorul placi.
Interpretarea rezultatelor s-a facut cu ajutorul softului bazei de date specifice pentru gram negative Microlog.

Fig.5. Sistemul Biolog - aparatul ce masoara turbiditatea inocului.

Fig.6. Sistemul Biolog - Cititorul automat de placi
III.3. CRESTEREA RHIZOBIILOR PE DIFERITE MEDII
Cultivarea microorganismelor pe medii lichide, aerate si agitate are deja o lunga traditie, iar experienta obtinuta in devzoltarea si optimizarea proceselor de biosinteza este deseori folosita pentru accelerarea realizarii tehnologiilor de obtinere a biopreparatelor cu rhizobii.
Provenind din microbiocenoze complexe si solicitante, rizobiile din biopreparate prezinta uneori specializari metabolice care, folosite in scopuri tehnologice pot conduce la solutii inovatoare.
Intre aceste specializari metabolice utile din punct de vedere tehnologic se inscrie si capacitatea de a utiliza produsii de metabolism ai altor microorganisme, capacitate care a fost folosita pentru a creste bradyrizobii pe melase fermentate cu drojdii.
Initial, in aceasta etapa a proiectului s-au cultivat bacteriile izolate pe mediile Fred si Waksman, Bond, Van Schreven, Vincent, Burton, Wright, si Lazareva, in vederea stabilirii necesitatilor nutritionale si a surselor de nutrienti.
pH-ul mediilor a fost ajustat la valoarea 7 cu NaOH respectiv HCl 0,1N. Flacoanele cu mediu au fost sterilizate 20 de minute la autoclav la 121 C. Experienta a fost organizata in trei repetitii, fiecare varianta cuprinzand trei flacoane de mediu cu tulpina bacteriana.
Dupa ce au fost inoculate flacoanele s-au incubat la 30C, iar observatiile privind cresterea bacteriilor s-au efectuat dupa 4 si 7 zile de la inoculare. S-a notat densitatea optica a cresterii bacteriene in mediul de cultura.
Tabel 3. Compozitia mediilor de cultura pentru bacteriile din genul Rhizobium
|
Ingrediente g/l |
Fred si Waksman |
Bond |
Hendrikson |
Van Schreven |
Vincent |
Burton |
Wright |
Lazareva |
|
Zaharoza | ||||||||
|
Manitol | ||||||||
|
K3PO4 |
| |||||||
|
K2HPO4 | ||||||||
|
KH2PO4 | ||||||||
|
MgSO4 x 7H2O | ||||||||
|
NaCl | ||||||||
|
(NH4)HPO4 | ||||||||
|
CaSO4 x H2O | ||||||||
|
MnSO4 | ||||||||
|
(NH4)2MoO4 x 2H2O | ||||||||
|
CaCO3 | ||||||||
|
Gluconat de Ca | ||||||||
|
Apa de drojdii |
100 ml |
50ml |
30ml |
100ml |
100ml | |||
|
Autolizat de Drojdii |
III.3.1. CULTIVAREA BACTERIILOR DIN GRUPUL RHIZOBIUM PE MEDIU HIDROGELIFIAT
A fost elaborat in cadrul acestei etape a proiectului un model experimental, care a presuspus incercarea de cultivare a rizobiilor pe un hidrogel superabsorbant numit Aquasorb.
Acesta este un fixator de apa care, atunci cand este incorporat in sol sau in substrat, absoarbe cantitati mari de apa si nutrienti. Spre deosebire de majoritatea produsilor care se hidrateaza, Aquasorb are proprietatea de a elibera usor apa absorbita si nutrientii, in acest fel permitand plantei sa aiba apa si nutrientii disponibili, functionand in cicluri absobtie-eliberare de apa. Produsul are proprietatea de a absorbi de 500 de ori greutatea lui in apa.
Fig.7. Formarea retelei de hidrogel
Ca materiale s-au folosit pungi urinare cu evacuare sterile, Aquasorb, mediu Lazareva lichid, compresor.
Procedura a fost relativ simpla, in pungile urinare, sterilizate in prealabil s-au introdus cate 4 g de hidrogel. Pungile au fost ambalate apoi individual precum si cate 4. Dupa dubla ambalare a acestora, produsele au fost trimise la sterilizat .
Sterilizarea s-a realizat cu radiatii , la instalatia IRASM, din cadrul IFIN-HH, la o doza de >25 kGy, timp de 37h 36min.
Dupa sterilizare, in pungile urinare s-au introdus cate 300 ml de mediu Lazareva lichid. Dupa ce s-a gonflat hidrogelul, pungile au fost inoculate cu tulpinile rhizobiene. Pentru asigurarea unor conditii de aerare corespunzatoare, cu ajutorul unui compresor probele trebuie gonflate cu aer steril. Astfel prelucrate probele au fost puse la incubat la 27sC, 4 zile.
III.4. VERIFICAREA PASTRARII CARACTERISTICILOR BIOLOGICE ALE RHIZOBIILOR
S-a realizat in aceasta etapa verificarea mentinerii caracteristicilor biologice ale tulpinii Rhizobium leguminosarum FL20, prin reproducerea infectiei la nivel de laborator.
Peste culturile bacteriene de pe mediul agarizat s-a adaugat apa distilata sterila in vederea obtinerii unei suspensii cu titrul de 106 ufc/ml. Cu aceasta suspensie bacteriana s-au tratat seminte de fasole. Semintele au fost imediat semanate in vase de vegetatie si depozitate in camera de crestere. La faza fenologica optima de formare a nodozitatilor au fost recoltate cate 5 plantute din fiecare vas pentru a stabili caracterul infectios al tulpini. S-au realizat izolarile si s-a evidentiat prezenta tulpinii cu care au fost inoculate semintele de fasole.
III.5.ELABORAREA UNUI MODEL EXPERIMENTAL PENTRU DETERMINAREA FENOMENULUI STINGERII DE QUORUM SENSING LA BACTERIILE PGPR
Mecanismul QS regleaza sporularea, sinteza de antibiotic, induce factorul de virulenta, diferentierea celulara si fluxul de nutrienti, competenta patogenica etc.
La densitati mici ale populatiilor se produce un nivel scazut de AHL. Cand densitatea celulelor creste, avand tendinta de a forma un film biologic invaziv, AHL produs in cantitate mare se acumuleaza in mediu si, cand se atinge concentratia critica, difuzeaza in celule legandu-se la receptorul specific fapt care duce la activarea sau represia exprimarii genelor care determina competenta ecologica a bacteriilor.
Dat fiind rolul important pe care acest fenomen il are in interactiile patogene (Loh si col., 2002), inactivarea sau degradarea moleculelor AHL constituie o cale inovativa de modulare a actiunii agroinoculantilor.
Ca metoda pentru modelul experimental propus, s-a folosit un procedeu de evidentiere a moleculelor de AHL, secretate de unele tulpini bacteriene, cu ajutorul unor bacterii biosenzor, de tipul Chromobacterium violaceum CV026.
Chromobacterium violaceum este o bacterie Gram negativa, nesporulanta, facultatuv anaeroba, care traieste in solurile si apele din zona tropicala si subtropicala ale globului. Pe mediile de cultura (ex. nutrient agar, MacConkey agar etc.) C. violaceum formeaza colonii usor convexe, netede, de culoare violet-intunecat, care este datorata producerii unui pigment violet insolubil in apa, numit violaceina.
Unele tulpini ale speciei C. violaceum sunt patogene, cauzand infectii piogenice sau septicemice la mamifere, incluzand si omul. Totodata violaceina are efecte antibiotice, antitumorale si activitate anti-Trypanosoma cruzi
|
|
Fig.8. Colonii de Chromobacterium violaceum ATCC 31532
Pornind de la studiul literaturii, au fost elaborate modele experimentale pentru evidentierea sensibilitatii de grup la bacteriile fixatoare de azot si la populatiile bacteriene cu actiune de favorizare a cresterii plantelor.
Modelele experimentale elaborate au implicat folosirea unor bacterii biosenzor, aplicate intr-un al doilea strat de agar care este depus peste bacteriile de testat inoculate in striuri ale mediului agarizat specific.
Tulpinile bacteriene folosite in cadrul experimentelor preliminare asociate elaborarii modelelor experimentale pentru evidentierea sensibilitatii de grup sunt prezentate in tabelul 4.
Tabel 4. Lista tulpinilor utilizate in cadrul experimentelor preliminare asociate elaborarii modele experimentale
|
Tulpina |
Provenienta |
Comportament |
Scopul |
Sursa |
|
Chromobacterium violaceum CV026 |
Muntanta transpozomi |
Tulpina tip |
Biosenzor; tulpina indicator pentru QS |
Institutul de Biologie, Universitatea Leiden |
|
Agrobacterium tumefaciens WCF47 |
Construct de fuziune Tra LacZ |
Tulpina tip |
Biosenzor; tulpina indicator pentru QS |
Institutul de Biologie, Universitatea Leiden |
|
Bradyrhizobium japonicum SOFR10 |
Nodozitati soia |
Fixator de azot simbiotic |
Determinare producere AHL |
Colectia ICDPP |
|
Rhizobium spp. FL400 |
Nodozitati bob |
Fixator de azot simbiotic |
Determinare producere AHL |
Colectia ICDPP |
|
Rhizobium leguminosarum bv. phaseoli Fl20 |
Nodozitati fasole |
Fixator de azot simbiotic |
Determinare producere AHL |
Colectia ICDPP |
|
Pseudomonas sp. 33 |
Sol de rizosfera din cultura de grau, judet Ilfov, 2007 |
Favorizant de crestere vegetala (PGPR), activitate CB in vitro |
Determinare producere AHL |
Izolat propriu |
|
Bacillus sp. |
Flora epifita de pe frunze de mar, ICDPP, 2007 |
Favorizant de crestere vegetala (PGPR), activitate CB in vitro |
Determinare producere AHL |
Izolat propriu |
Tulpinile biosenzor au fost cultivate pe mediul agarizat Luria Bertani (LB) cu urmatoarea compozitie: 10g bactotriptona; 5g extract de drojdii; 10g NaCl; 20g agar-agar; pH 7.2, sterilizat 20 minute la 121 OC.
Tulpinile din grupul Rhizobium s-au cultivate pe mediu Lazareva cu urmatoarea compozitie: manitol - 10 g/l, K2HPO4 - 0,5 g/l, MgSO4x7H2O - 0,2 g/l, NaCl - 0,2 g/l, CaSO4 - 0,1 g/l, MnSO4 - 0,005 g/l, (NH4)2MoO4x 2H2O - 0,002 g/l extract de drojdii - 1 g/l, agar-agar - 20 g/l, pana la 1000 ml apa distilata; pH 6,8.
Culturile de Pseudomons sp. si Bacillus sp. au fost crescute pe mediu PDA , pH 6.8-7.0, sterilizat 20 minute la 121 OC.
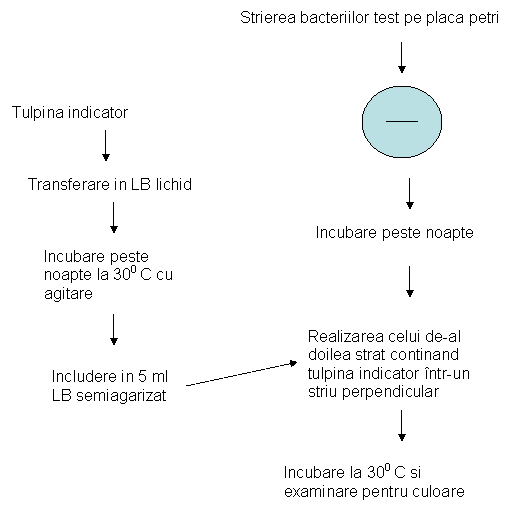

Fig. 9. Procedura de determinare prezentiei mecanismului de quorum sensing
Pentru realizarea stratului superior de acoperire care contine tulpina senzor CV 026, s-a preparat un mediu LB semisolid prin adaugare de 0.5% (g/v) agar. In cazul in care s-a folosit ca tulpina indicator Agrobacterium tumefaciens WCF47 in mediu agarizat s-au adaugat si 80 mg/ml X-gal. Un rezultat QS pozitiv a fost indicat de prezenta pigmentarii microorganismului indicator in vecinatatea organismului testat.
Biotestul pentru evidentierea QS a fost realizat conform procedurii descrise in figura 9. In centrul placii petri cu mediu agarizat s-au insamantat bacteriile test sub forma unor striuri. Placile au fost incubate peste noapte, la 300C pentru a ajunge in faza stationara de crestere. Peste cresterea bacteriana, in placa petri s-a realizat un al doilea strat de mediu LB semisolid, racit la 450 C . In acest strat a fost practicat al doilea striu, perpendicular pe directia primelor bacterii, in care s-au inoculat 50 l de suspensie de bacterii biosenzor cu titrul de 109 ufc/ml. S-a incubat peste noapte la 300 C si s-a examinat pentru aparitia culorii (violete in cazul bacteriei Chromobacterium violaceum 026 ; albastre la Agrobacterium tumefaciens WCF47). S-a apreciat ca rezultat pozitiv, respectiv producere de AHL, prezenta culorii in jurul striului de crestere bacteriana.
Pentru evidentierea tulpinilor cu activitate de stingere a cvorumului a fost elaborata o modificare a metodei descrise mai sus, care a constat in lasarea in prealabil a unui interval de timp, in care posibilele enzime care degradeaza AHL, sintetizate de tulpinile cu capacitate de modulare a QS, sa aiba timp sa actioneze asupra moleculei semnal produsa de tulpinile de agroinoculanti.
In placa petri cu mediu Luria Bertani, au fost realizate godeuri cu diametrul de 6 mm, asezate in perechi la distanta de 10mm. Placile au fost incubate peste noapte la 300 C, dupa care s-a realizat al doilea strat cu tulpina indicator si s-a reluat o incubare de 16 ore, moment in care s-a apreciat aparitia culorii datorata prezentei moleculelor de AHL nedegradate in mediu.
Dupa incubarea peste noapte, bacteriile biosenzor din cel de-al doilea strat au dezvoltat o reactie de culoare specifica. In cazul bacteriilor cu activitate de favorizare a cresterii si dezvoltarii plantelor (din genul Bacillus) s-a folosit ca baza, tulpina indicator / biosenzor Chromobacterium violaceum Cv 026, evidentiindu-se producerea de violaceina, iar in cazul bacteriilor din grupul Rhizobium s-a folosit ca indicator, tulpina de Agrobacterium tumefaciens WCF47. Moleculele de N-acil-homoserin-lactona, AHL, au fost detectate prin reactia cromogenica a b-galactozei raportor cu substratul X-gal, care genereaza o culoare albastra.
III.6. CARACTERIZAREA UNOR TULPINI DE RHIZOBIUM CE PREZINTA ACTIVITATE ACC DEAMINAZICA SI CARE NODULEAZA FASOLEA
Fasolea este una plantele leguminoase care formeaza nodozitati fixatoare de azot cu mai multe specii din genul Rhizobium (R. leguminosarum bv. phaseoli; R. giardinii bv. phaseoli; R. gallicum bv. phaseoli; R. lusitatum; R. etli; R. tropici; R. mongolese).
Unele din tulpinile care noduleaza fasolea prezinta o activitate semnificativa a ACC-deaminazei. Acidul 1-aminociclopropan carboxilic (ACC) este precursorul biosintetic al etilenei, iar etilena (si implicit ACC) exercita (pe langa alte efecte inhibitorii) si un semnficativ efect de reducere a formarii nodozitatilor plantelor. Producerea de ACC-deaminaza reprezinta una din caile utilizate de bacterii pentru cresterea numarului de nodozitati. Datorita producerii de ACC-deaminaza si de fitohormoni aceste tulpini de Rhizobium au si o actiune semificativa de stimulare a cresterii plantelor neleguminoase, cum sunt ar fi de exemplu cele din familia Brassicaceae (Noel si. col, 1996) sau Poaceae (Matiru si Dakora, 2004).
Experimentul si-a propus identificarea si caracterizarea fenotipica prin microsistemul Biolog a unor tulpini de Rhizobium care noduleaza fasolea si care prezinta activitate ACC deaminazica.
S-a procedat la izolarea unor noi tulpini de Rhizobium specifice pentru Phaseolus vulgaris prin tehnica dilutiilor asociata tehnicii capturii pe radacini de mazariche paroasa. 1 g de sol s-au omogenizat cu 99 ml sol fiziologic sterilizat prin autoclavare. Din omogenizat s-au prelevat 0,1 ml care au fost diluati aseptic. Din dilutiile zecimale s-au prelevat 0,1 ml care au fost depusi pe radacinile unei plantule rezultate din seminte de Vicia villosa sterilizate la suprafata prin spalari succesive cu hipoclorit 2%, alcool etilic 30% si apa distilata sterila.
Semintele sterilizate au fost germinate pe apa agarizata. Plantulele inoculate au fost cultivate apoi in vase de vegetatie cu mediu semiagarizat lipsit de azot. Nodulii formati au fost reluati, s-au separat cei de culoare roz, care au fost sfaramati aseptic in ser fiziologic steril. Din suspensia rezultata s-au realizat dilutii zecimale. Din dilutii s-au prelevat 0,1 ml, care au fost raspandite uniform pe suprafata unui mediu manitol - rosu de Congo. Coloniile de Rhizobium au fost identificate prin culoarea roz formata (spre deosebire de contaminanti care preluau tot rosul de Congo) si purificate prin pasaje repetate pe mediu Lazareeva.
Tulpinile izolate au fost testate in privinta eficacitatii fixarii azotului prin determinarea numarului de nodozitati formate pe plantele test. Ca tulpina etalon s-a folosit tulpina de Rhizobium leguminosarum bv. phaseoli FL 220.
Pentru izolarea bacteriilor s-a folosit mediul manitol (YEM) - rosu de Congo, cu urmatoarea compozitie: extract de drojdie: 0,2 g; manitol: 10 g; NaCl 0.1 g; 0.01g CaCO3; rosu de Congo 2,5 ml dintr-o solutie 1%, 15 g agar apa pana la 1000ml.
Tabel 5. Solutia nutritiva Crone modificata de Bryan (solutie nutritiva fara azot), utilizata pentru cultivarea plantelor inoculate cu rhizobii.
|
Sare minerala |
Formula |
grame / l |
mg/l - ppm (solutia finala ) |
|
Clorura de potasiu |
KCl |
10,0 g |
393,3 ppm, K |
|
Sulfat de calciu |
CaSO4 . 2(H2O) |
2,5 g |
116,3 ppm, Ca |
|
Sulfat de magneziu |
MgSO4 . 7(H2O) |
2,5 g |
18,5 ppm, Mg |
|
Fosfat tricalcic |
Ca3(PO4)2 |
2,5 g |
80,2 ppm, P |
|
Fosfat feric cu citrat de fer |
2,5 g |
26,2 ppm, Fe |
Tabel 6. Solutia de oligo si microelemente care se adauga la solutia Crone
|
Sare minerala |
Formula |
grame / l |
mg/l - ppm (in solutia finala ) |
|
Acid boric |
H3BO3 |
50 ppm B |
|
|
Sulfat de mangan |
MnSO4 . H2O |
50 Mn |
|
|
Sulfat de zinc |
ZnSO4 . 7(H2O) |
10 Zn |
|
|
Sulfat de cupru |
CuSO4 . 5(H2O) |
08 Cu |
|
|
Molibdat de sodiu |
Na2MoO4.2H2O |
04 Mo |
|
|
Clorura de cobalt |
CoCl2 . 6(H2O) |
001 Co |
Mediul agarizat fara azot pe care au fost cultivate plantulele de mazariche paroasa a fost mediul nutritiv Crone modificat de Bryan (Hewit, 1965), a carui formula este prezentat in tabelul 5 (macroelemente nutritive), la care se adauga 5 ml/l din solutia de (oligo si microelemente nutritive) prezentata in tabelul 6 si se agarizeaza cu 10 g/l.
Pentru cultivarea in laborator a bacteriilor din genul Rhizobium s-a utilizat mediul Lazareva avand urmatoarea compozitie: 10 g manitol, 0,5 g K2HPO4, 0,2 g MgSO4x7H2O, 0,2 g NaCl, 0,1 g CaSO4 , 0,005 g MnSO4 , 0,002 g (NH4)2MoO4x 2H2O, 1 g extract de drojdii, 20 g agar-agar, 1000 ml apa distilata; pH 6,8.
Mediul de cultura special folosit pentru selectarea rhizobiilor cu activitate ACC deaminazica este mediul DF, cu urmatoarea compozitie: glucoza 2g/l, acid gluconic 2g/l, acid citric 2g/l, KH2PO4 4g/l, MgSO4 0.2g/l, NaCl 0.1g/l, pH 6.8, solutie micronutriente 10 ml , Agar 18g/l.
Solutia micronutriente: CaCl2 200mg/l, FeSO4 200mg/l, H3BO3 15mg/l,
ZnSO4 20mg/l, Na2MoO4 10mg/l, KI 10mg/l, NaBr 10mg/l, MnCl2 10mg/l, CoCl2 5mg/l, CuCl2 2mg/l, AlCl3 2mg/l, NiSO4 2mg/l.
Determinarea incadrarii taxonomice a rhizobiilor izolate. S-a realizat cu ajutorul sistemelor microtest Biolog. Placi Petri cu mediu special BUG (Biolog Universal Growth), au fost inoculate cu tulpinile de testat, izolate anterior din nodulii de pe radacinile de Vicia Villosa. Placile au fost incubate peste noapte la termostat la 28C. Dupa 24 de ore, au fost inoculate microplacile GN astfel: din fiecare placa s-au prelevat cu anse sterile, de unica folosinta, cantitatati mici de masa bacteriana, care au fost omogenizate in flacoane cu lichid de inoculare, specific tipului gram negativ. Turbiditatea lichidului s-a verificat la Biolog dupa fiecare omogenizare, pana la atingerea valorii 52 de unitati. Cu o micropipeta, s-au insamantat cate 150 l suspensie in microplacile GN,apoi au fost lasate la incubat 16-24 ore. Citirea placilor s-a facut la microstatia Biolog iar interpretarea rezultatelor s-a realizat cu ajutorul softului bazei de date pentru bacterii gram negative de la Microlog.
Identificarea tulpinilor de Rhizobium cu activitate ACC-deaminazica . S-au realizat suspensii bacteriene pentru fiecare tulpina de testat. S-a luat 1 ml suspensie si s-a transferat in 50ml mediu DF broth suplimentat cu 2g/l sulfat de amoniu ca sursa de azot. S-a lasat la incubat timp de 24 ore la 30C.
Apoi s-a transferat 1 ml in 50ml mediu DF broth suplimentat cu 0.3033 ACC ca sursa de azot, si s-a incubat din nou 24 ore la aceeasi temperatura.
Dilutii din ultima cultura au fost etalate pe placi petri cu mediu DF solid suplimentat tot cu ACC, placile s-au incubat 48 ore la temperatura camerei.
S-au prelevat coloniile izolate si apoi s-a facut testarea proprietatilor de favorizare a cresterii plantelor.
Evidentierea proprietatilor de stimulare a cresterii plantelor s-a realizat prin biotestarea efectului fitostimulator al tulpinilor bacteriene asupra radacinilor de rapita si prin eficienta formarii de nodozitati la plante de fasole.
Biotestarea in vitro a efectului fitostimulator al biopreparatelor utilizate pentru stimularea cresterii plantelor. Semintele de rapita (cv. Triangle) au fost sterilizate la suprafata prin metoda descrisa de Dobbelaere et al. (1999), prin spalari repetate cu Domestos (Unilever South-East Europe) si clatiri cu apa distilata. Semintele astfel dezinfectate au fost plasate pe discuri de hartie de filtru sterile (depuse in placi Petri f 9 cm) si umectate cu 1,5 ml de apa sterila. Dupa 24 ore de incubare la intuneric la 25C semintele / achenele au fost inoculate cu 0,1 ml suspensie de bacterii in tampon fosfat salin steril. Martorul a fost tratat cu tampon fosfat steril. Dupa 72 de ore de incubare la intuneric a fost masurata lungimea radacinilor.
Teste de formare a nodozitatilor. Testele biochimice de caracterizare fenotipica au fost completate cu teste de determinare a reactiei diferitelor plante test (din familia leguminoaselor) la inoculare. S-a testat eficienta de formare a nodozitatilor de catre izolatele bacteriene la plante de fasole, linte si naut. Semintele sterilizate au fost bacterizate cu suspensii bacteriene ale tulpinilor testate, apoi au fost semanate in vase de vegetatie, in nisip sterilizat la etuva. Dupa 28 de zile, s-a realizat determinarea activitatii nitrogenazice cu ajutorul aparatului Qubbit (care determina hidrogenul produs de nitrogenaza.
III.7.ESTIMAREA EFICACITATII FIXARII SIMBIOTICE A AZOTULUI PRIN DETERMINAREA HIDROGENULUI
Ca si fotosinteza, fixarea N2 este un fundamental proces biologic esential in ciclul global al nutrientilor. Biologic, fixarea azotului in plantele leguminoase necesita dezvoltarea unei simbioze intre bacteria din sol si radacina plantei. Cea mai comuna endosimbioza gasita in radacinile leguminoaselor o formeaza bacteriile din grupul Rhizobium.
Determinarea acumularii de azot fixat a fost initial realizata prin determinarea comparativa a azotului total in plantele provenite din seminte inoculate si din seminte neinoculate.
Diferitele metode de testare folosite urmaresc atat stabilirea modului in care s-a realizat pastrarea caracteristicilor biologice dupa cultivarea pe medii industriale si conditionare, cat si verificarea eficacitatii diferitelor biopreparate in diferite conditii agropedoclimatice.
Una din metodele de laborator recent dezvoltata pentru testarea eficacitatii tulpinilor de rhizobii in conditii de laborator este metoda de dozare a hidrogenului produs de nitrogenaza in conditiile trecerii peste nodozitati a unui sistem gazos din care lipseste azotul. Mai jos sunt prezentate primele rezultate obtinute in cadrul testelor efecuate la ICDPP. Scopul acestui experiment a fost de a demonstra ca activitatea nitrogenazica si rata fixarii azotului in legume poate fi determinata prin masurarea ratei evolutiei H2 din nodozitatile radacinii. Experimentul introduce metoda analizei schimbului de gaze pentru masurarea proceselor fiziologice si arata cum aceasta metoda poate elucida parametrii biochimici prin intermediul nodozitatilor fara a fi necesara extractia din tesuturi.
S-a lucrat cu plante de fasole, inoculate cu tulpinile de Rhizobium care noduleaza fasolea caracterizate anterior - FL220, FL400. Microorganismele au fost cultivate pe mediu Lazareeva. Plantele au fost crescute pe solutie nutritiva Crone lipsita de azot.
Determinarea activitatii nitrogenazice prin producerea de hidrogen. S-a folosit dispozitivul experimental prezentat in figurile de mai jos. Acest dispozitiv experimental permite determinarea ratei de eliberare a hidrogenului de catre tulpinile fara Hup (enzima de preluare a hidrogenului).
Fig.10. Cultivare pe medii hidroponice lipsite de azot a plantelor
provenite din seminte / plantule inoculate cu rhizobii (pentru a se
elimina interferentele azotului fixat)


Fig.11 Ansamblu de laborator pentru determinarea activitatii nitrogenazice prin masurarea hidrogenului produs in timpul fixarii azotului
Rata eliberarii de H2 furnizeaza o masura a activitatii nitrogenazice aparente (ANA), deoarece doar o proportie din fluxul de electroni prin nitrogenaza este folosit pentru reducerea protonilor. Pentru masurarea activitatii nitrogenazice totale (TNA) este necesara expunerea radacinilor cu nodozitati la un amestec de gaz lipsit de N2, cum ar fi amestecul Ar:O2 (80:20). Trecerea unui gaz lipsit de azot molecular permite determinarea nitrogenazei si in cazul tulpinilor care au hidrogenaza de preluare (Hup). Rata de fixare a azotului de calculeaza conform formulei:
Rata de fixare N2 = (TNA-ANA)/3
Plantele de fasole crescute pe medii lipsite de azot au fost recoltate, iar sistemul radicular a fost introdus in cuva si supus diferitelor amestecuri de gaze (aer; argon: oxigen 80:20).
III.8. CULTIVAREA UNEI TULPINI DE AZOSPIRILLUM BRASILIENSE PE DIFERITE MEDII DE CULTURA.
Unul dintre obiectivele prioritare ale agriculturii moderne este reprezentat de reuducerea dependentei culturilor agricole de fertilizarea chimica cu azot, avand in vederea costurilor importante, atat de productie a ingrasamintelor chimice, cat si sociale (costuri ale poluarii, datorate deopotriva productiei si utilizarii). Activitatea biologica a azotului, aflat in cantitati practic inepuizabile (circa 75% din volumul atmosferei) se realizeaza prin procesul de fixare datorat unei reactii enzimatice guvernata de sistemul nitrogenazic.
Inca din secolul XIX a fost semnalata capacitatea de fixare a azotului atmosferic la diferite grupe de microorganisme. In prezent sunt descrise aproximatic 200 de specii de microorganisme diazotrofe, libere sau incluse in sisteme asociate cu alte organisme vegetale.
Speciile de Rhizobium si Azospirillum au primit o atentie deosebita datorita efectelor pe care le produc ca urmare a inocularii.
La speciile de Azospirillum efectele benefice constau in sporuri medii de recolta de 20%, in unele cazuri fiind chiar semnalate si sporuri de 60% datorate fixarii azotului dar si stimularii cresterii plantelor si a modificarilor absorbtiei substantelor nutritive.
In vitro, bacteria isi pastreaza viabilitatea in conditii de uscaciune severa datorita capacitatii sale de a produce forme de rezistenta. Aceasta calitate nu s-a manifestat in experientele de camp, in conditiile in care nu era asociata cu radacinile unei plante gazda. Azospirillum ca inoculant trebuie aplicat cu un titru destul de mare pentru a stabili marimi de populatii fixatoare de azot, eficiente.
S-a demonstrat experimental faptul ca administrarea lui Azospirillum ca inoculant imbunatateste rasarirea plantelor, inaltimea plantelor, momentul infloririi, substanta uscata totala, rezistenta la seceta, continutul in azot din masa vegetala si seminte, componentele recoltei si productia de boabe.
Obiectul cercetarilor din aceasta etapa a fost stabilirea conditilor optime pentru cresterea bacteriei pe diferite medii si a producerii de biomasa.
Ca material biologic s-a folosit tulpina SP 001 de Azospirillum brasiliense, izolata din rizosfera unei plante de grau, recoltata dintr-o sola cu monocultura indelungata. Tulpina face parte din colectia de microorganisme utile din cadrul ICDPP.
Mediile de cultura utilizate pentru aprecierea capacitatii de crestere a bacteriei au fost mediile lichide AFM 10, AFM 20, si Dbereiner semisolid, repartizate in baloane Erlenmeyer.
Retele mediilor sunt prezentate in tabelul de mai jos.
Experienta a cuprins cele variante de mediu descrise mai sus, fiecare in 3 repetitii pentru fiecare din cele momente de timp alese pentru observatii 24, 48 si 72 ore.
Fiecare balon Erlenmeyer a fost inoculat cu 10% suspensie bacteriana cu titrul calibrat la 109 ufc. Dupa inoculare, baloanele au fost incubate la 28C, in conditii de agitare continua, pe un agitator la 140rpm.
La fiecare 24, 48, si 72 ore, baloanele inoculate au fost analizate in ceea ce priveste cresterea bacteriana.
Au fost facute de asemenea observatii asupra variatiei pH-ului mediilor inainte si dupa inocularea bacteriilor, la cele trei intervale de timp de incubare, utilizand un pH-metru de laborator.
Cresterea bacteriana a fost apreciata prin: cantarirea biomasei umede la balanta analitica si masurarea nefelometrica a turbiditatii, estimata in NTU.
Centrifugarea mediilor inoculate s-a facut la 10000 rpm timp de 10 min, apoi s-a decantat supernatantul, iar sedimentul a fost cantarit.
Tabel 7. Compozitia mediilor de cultura testate pentru cresterea bacteriilor utile plantelor de cultura Azospirillum brasiliense, tulpina SP001
AFM 10 |
AFM 20 |
Dbereiner. |
|||
CaCl2 |
20 mg |
CaCl2 |
20 mg |
CaCl2x2H2O |
20 mg |
NaCl |
100 mg |
NaCl |
100 mg |
NaCl |
100 mg |
MgSO4x 7 H2O |
200 mg |
MgSO4x 7 H2O |
200 mg |
MgSO4x 7 H2O |
200 mg |
FeSO4x 7 H2O |
10 mg |
FeSO4x 7 H2O |
10 mg |
|
|
Na2Mo4 x 2 H2O |
2 mg |
Na2Mo4 x 2 H2O |
2 mg |
Na2Mo4 x 2 H2O |
20 mg |
K2HPO4 |
600 mg |
K2HPO4 |
600 mg |
K2HPO4 |
600 mg |
KH2PO4 |
400 mg |
KH2PO4 |
400 mg |
KH2PO4 |
400 mg |
Extract de drojdii |
3 g |
Extract de drojdii |
3 g |
|
|
Glucoza |
10 g |
Glucoza |
15 g |
|
|
|
|
|
|
|
acid malic |
5 g |
|
|
|
|
|
KOH |
4 g |
|
|
|
|
|
FeCl3x3H2O |
16 mg |
|
|
|
|
|
albastru deBromtimol |
2 ml |
|
|
|
|
|
Agar |
2 g |
Apa |
1000 ml |
Apa |
1000 ml |
Apa |
1000ml |
III.9. TESTAREA ACTIVITATII BIOLOGICE A BIOMASEI DE BACTERII A. BRASILIENSE
Bacteriile Azospirillum sunt rizobacterii diazotrofe, favorizante ale cresterii si dezvoltarii vegetale (PGPR - "plant growth promoting rhizobacteria") cu o actiune complexa asupra plantelor de cultura. Unele din efectele produse in tesuturile vegetale, si in special in cele radiculare, sunt datorate producerii de fitohormoni in situ de catre microorganismele din genul Azospirillum. In cadrul acestui test biologic s-a urmarit evidentierea activitatii fitohormonilor giberelici produsi de bacterii utile plantelor de cultura din genul Azospirillum, pentru a se stabili prezervarea calitatii biologice de favorizant diazotrof dupa cultivarea pe mediile de cultura.
Evidentierea si estimarea activitatii fitohormonilor giberelinici s-a efectuat prin biotestul inducerii a-amilazei din endospermul de orz cuplat cu tehnica difuziei radiale in gel.
Etapele procedurii utilizate au fost:
inducerea biosintezei a-amilazei in endospermul boabelor de orz de catre fitohormonii prezenti in extractele de testat;
difuzia radiala in gelul de agar-amidon a enzimelor sintetizate sub actiunea fitohormonilor si hidroliza consecutiva a amidonului;
evidentierea zonei de actiune a enzimei prin colorarea substratului.


a) proba b) martor
Fig.10. Evidentierea activitatii fitohormonilor giberelinici
In cadrul acestui test s-au folosit urmatorii reactivi:
gel de agar - amidon. In 100 ml tampon fosfat pH 6,9 diluat cu 100 ml apa, 2,5 g agar si 1 g de amidon s-au dizolvat la cald si s-au fiert pana cand solutia a devenit limpede;
solutia tampon fosfat pH 6,9 a fost preparata astfel: 590 mg acetat de sodiu + 1,47 g veronal sodic + 1,4 g NaCl + 780 mg CaCl2 s-au dizolvat in apa disitilata, s-au adaugat 62 ml HCl 0,1 N si s-a adus la un volum final de 250 ml cu apa distilata;
solutie de developare: 45 ml iod 0,1 N cu 55 ml etanol absolut;
extract de biomasa bacteriana realizat prin osmoza inversa;
endosperm de orz din seminte curatate de palee si de la care s-a inlaturat, prin taiere, partea care contine embrionul.
Modul de lucru a fost:
depunerea unui gel de agar-amidon in grosime de 2-3 mm pe suport reprezentat de lame microscopice;
practicarea in gel a unor godeuri cu diametrul de 3 mm;
depunerea in godeuri a endospermului de orz (jumatate de bob fara embrion) si a extractului de testat;
incubarea 72 h la temperatura camerei in incinta umeda;
colorarea cu solutie de developare a gelului;
observarea existentei unei zone de difuzie.
Fitohormonii giberelinici au fost extrasi din mediul de cultura prin dializa inversa. Cultura de bacterii in mediul de cultura s-a centrifugat (7500 g, 30 min), iar supernatantul a fost concentrat, trecut in sacul de dializa sii supus extractiei in acetat de etil (dializa inversa).
Interpretarea rezultatelor s-a facut cu ajutorul unei curbe standard, obtinuta prin utilizarea in aceeasi procedura experimentala a diferitelor concentratii de acid giberelic (GA3). Biotestul s-a realizat in trei repetitii, utilizand ca martori extracte ale mediilor de cultura folosite pentru multiplicarea bacteriilor (AFM10, AFM20, Dbereiner).
III.10. TEHNOLOGIA DE REALIZARE A INOCULUI DE LABORATOR
Inoculul de laborator reprezinta cultura pura de microorganisme aduse in forma viabila si care este utilizata la insamantarea ulterioara a mediilor de cultura.
Tehnologia a fost similara pentru bacteriile din genul Rhizobium si pentru cele din genul Azospirillum, si s-a realizat dupa cum urmeaza:
Un numar de cinci eprubete stoc din camera de depozitare a culturii de microorganisme au fost aduse lent la temperatura de 20-24 C (temperatura camerei). S-au pregatit cinci eprubete cu cinci mililitri de mediu inclinat Lazareva insamantat cu tulpina de Rhizobium leguminosarum bv. phaseoli FL20 din eprubetele stoc.
S-au incubat eprubetele inoculate la 28 C timp de 3-5 zile si apoi s-a controlat microscopic puritatea culturii din eprubete.
Pentru realizarea inocului de Azospirillum la nivel de laborator s-au insamantat cinci eprubete cu trei mililitri de mediu D bereiner semisolid, din eprubetele stoc ale culturii SP001. Au fost incubate la 30 C timp de 5 zile, dupa care s-a verificat puritatea culturii din eprubete.
In ambele cazuri, culturile proaspete s-au multiplicat in 15 eprubete cu cate cinci mililitri de mediu inclinat Lazareva pentru Rhizobium, respectiv D bereiner pentru Azospirillum, care constituie inoculul de laborator.
III.11. CONDITIONAREA BIOMASELOR DE RHIZOBIUM SI AZOSPIRILLUM CA BIOPREPARATE AGLOMERATE - FLOCULI
Optimizarea tehnologiei de utilizare a unor produse biologic active pentru stimularea cresterii si dezvoltarii plantelor si a aprovizionarii culturilor cu azot fixat a implicat si modificarea formei de conditionare a bacteriilor utile din genurile Azospirillum si (Brady)rhizobium. Formele uzuale (ca de ex. suspensie microbiana absorbita in turba) nu adera corespunzator la unele seminte tratate si/sau nu asigura o rata corespunzatoare de supravietuire a bacteriilor inoculante si /sau de colonizare a rizosferei. Din acest motiv a fost stabilit, un protocol care a impus si o modificare a mediului de cultura pentru a favoriza formarea de agregate (floculi) de bacterii din genul Azospirillum, care sa permita co-flocularea bacteriilor utile asiguand in final o mai buna rezistenta la conditiile dificile de mediu (uscaciune, lipsa de substrat metabolic etc.) si o mobilitate ridicata a inoculului.
Mediul de cultura folosit pentru cultivarea bacteriilor utile din genul Azospirillum are avantajul de a favoriza pe de o parte acumularea de polibeta-hidroxibutirat, iar pe de alta parte flocularea microorganismelor in agregate avand densitati de peste 1015 ufc/ml. Acumularea de polibetahidroxibutirat asigura o buna rezistenta a bacteriilor la factorii de stres (Kadouri si col. 2003), iar agregarea in floculi favorizeaza recoltarea si supravietuirea bacteriilor (Neyra si col., 1997).
In fig.11. este prezentat aspectul microscopic al unei culturi de Azospirillum brasilense Sp001 pe mediu AFM 20.
Fig. 11. Aspectul in microscopie optica al culturii de 48 ore Azospirillum brasilense Sp001 in mediul lichid AFM 20, aerat si agitat.
Pentru asigurarea unui inocul bogat in bacterii cu mobilitate ridicata, s-a elaborat un procedeu tehnologic de co-floculare a bacteriilor agroinoculante din grupurile Azospirillum si Rhizobium cultivate pe mediu lichid, aerat si agitat.
Procedeul elaborat permite obtinerea unui inocul bogat in bacterii (min 1011 ufc/ g de produs final) capabil sa colonizeze rapid rizosfera plantelor de cultura.
Mod de lucru:
Cultivarea bacteriilor din grupul Rhizobium (R. leguminosarum bv. phaseoli izolatul FL20 ) pe mediu Lazareva hidrogelifiat timp de 7 zile.
Resuspendarea inoculului in mediu AFM 20S, ce contine acid malic in loc de glucoza, in proportie de 10% (s-au inoculat 10 ml de suspensie bacteriana in 100 ml mediu);
Cultivarea bacteriilor din grupul Azospirillum (Azospirillum brasiliense Sp001), in mediu lichid AFM20 aerat si agitat, timp de 2 zile. Agitarea a avut loc la 140 rpm, in vederea formarii de flocoane;
Normalizarea suspensiilor de bacterii Rhizobium si Azospirillum, la o densitate optica de circa 0,4. Aceasta densitate optica corespunde unei suspensii cu titrul de 108 ufc/ml.
Realizarea co-inocularii in raport de 1:1 pe mediu AFM 20S;
Cultivarea peste noapte, recoltarea biomasei floculante prin decantarea si uscarea acestei biomase in curent de aer in conditii blande (max. 35C).
Pentru cultivarea bacteriilor Azospirillum brasilense s-au folosit mediul lichid AFM20 si mediul semisolid Dbereiner, ale caror formule au fost prezentate deja. Mediul Dobereiner a fost folosit pentru mentinerea bacteriilor, iar mediul AFM 20 pentru producerea biomasei de azospirillii necesara pentru conditionare.
CAPITOLUL IV. REZULTATE SI DISCUTII
IV.1. Izolarea si cresterea pe diferite medii
Pe mediul Lazareva cu rosu de congo din plantele de soia au fost izolate si purificate 11 tulpini din trifoi s-au obtinut 12 izolate, de la fasole 17 izolate FAP 2340 si de la mazare 9 izolate.
S-a constatat ca viteza de crestere pe mediul Lazareva a fost de 4 zile pentru izolatele de la fasole, mazare si trifoi si de 7 zile pentru cele de la soia.
Coloniile bacteriene realizate prin epuizare de ansa au fost observate dupa 4 si dupa 7 zile de incubare la termostat. Pe mediul nutritiv Lazareva, tulpinile bacteriene au prezentat caractere morfologice si culturale distincte si caracteristice pentru fiecare biovar. Caracteristicile morfologice inregistrate la fiecare specie de Rhizobium izolata au fost confirmate de Manualul sistematic a lui Bergey.
Fig. 12. Aspectul unei placi insamantate cu Rhizobium leguminosarum bv. phaseoli FL20
Tabel 8. Caracteristicile tulpinilor bacteriene izolate din nodozitatile unor leguminoase
|
Planta Gazda |
Aspecte morfologice ale coloniilor |
Viteza de crestere pe mediul Lazareva |
Specia bacteriana |
|
Fasole |
Circulare, convexe semitranslucide, mucilaginoase, 2-4mm diam. |
4 zile |
Rhizobium leguminosarum bv.phaseoli |
|
Mazare |
Circulare, convexe semitranslucide, mucilaginoase, 2-4mm diam. |
4 zile |
Rhizobium leguminosarum bv. leguminosarum |
|
Trifoi |
Circulare, convexe semitranslucide, mucilaginoase, 2-4mm diam. |
4 zile |
Rhizobium leguminosarum bv. trifoli |
|
Soia |
Circulare, punctiforme, convexe, opace, albe, granulare, 1mm diametru. |
7 zile |
Bradyrhizobium japonicum |
Analizand valorile densitatii optice obtinute prin cultivarea tulpinilor selectionate pe cele 7 medii de cultura, cunoscute din literatura ca fiind specifice pentru bacteriile din genul Rhizobium, s-a constatat ca pe substraturile care au inclus manitolul in compozitie tulpinile izolate de la soia au avut o dezvoltare mai buna decat pe mediile cu zaharoza, si au atins valorile maxime (82 - 99,4) in 7 zile de la inoculare, dupa cum se observa in tabelele 9 si 10. Tulpinile izolate de la mazare, fasole si trifoi au avut o crestere mai buna pe mediile cu zaharoza (88,8 - 96,4) au ajuns la cresterea maxima dupa 4 zile de la inoculare. S-a constatat ca mediul Lazareva a asigurat o crestere foarte buna pentru tulpinile izolate de la soia si satisfacatoare pentru tulpinile izolate de la mazare, fasole si trifoi.
Bacterizarea plantelor gazda cu tulpinile bacteriene pentru reproducerea infectiilor la nivel de laborator, a determinat formarea unui numar mare de nodozitati la colet, dintre care un procent foarte mare (peste 85%) erau active, stare exprimata si prin greutatea proaspata si uscata a nodozitatilor; datele sunt prezentate in tabelul 11 de mai jos.
Tabel 9. Densitatea optica a cresterii bacteriene pe diferite medii de cultura inoculate cu tulpini izolate din nodozitati - dupa 4 zile
|
Tulpina |
Fred si Waksman |
Bond |
Hendrikson |
Van Schreven |
Vincent |
Burton |
Wright |
Lazareva |
|
FL 20 | ||||||||
|
MAP 48 | ||||||||
|
TAP 21 | ||||||||
|
SOAP 10 |
Tabel 10. Densitatea optica a cresterii bacteriene pe diferite medii de cultura inoculate cu tulpini izolate din nodozitati - dupa 7 zile
|
Tulpina |
Fred si Waksman |
Bond |
Hendrikson |
Van Schreven |
Vincent |
Burton |
Wright |
Lazareva |
|
FL 20 | ||||||||
|
MAP 48 | ||||||||
|
TAP 21 | ||||||||
|
SOAP 10 |
Tabel 11. Efectul inocularii plantelor cu tulpini din colectia de lucru
|
Inoculant |
Planta Test |
Nr. total nodozitati/pl |
% nodozitati active/planta |
Greutate nodozitatii(g)/pl |
||
|
La colet |
Pe radacini secundare |
proaspata |
Uscata |
|||
|
SOAP 10 |
Soia | |||||
|
FL 20 |
Fasole | |||||
|
MAP 48 |
Mazare | |||||
|
TAP 21 |
Trifoi | |||||
Cantitatea de biomasa obtinuta prin cultivarea tulpinilor selectionate pe diferite medii confirma datele inregistrate prin determinarea densitatii optice.
IV. 2. CARACTERIZAREA UNEI TULPINI DE RHIZOBIUM LA BIOLOG
In fig. 13-17 sunt prezentate diferite aspecte din timpul determinarilor experimentale cu ajutorul microsistemului Biolog. Acest microsistem utilizeaza microplaci de titrare cu 95 de godeuri in care exista diferite surse de carbon impreuna cu un colorant tetrazolic, care in urma exprimarii metabolismului bacterian formeaza un precipitat violet, generand astfel reactia de culoare. Exista diferite tipuri de microplaci specifice diferitelor tipuri de microorganisme (GN2, GN2-Ent, GP, YT, s.a.), iar tiparele de amprentare metabolica sunt comparate si identificate folosind softul bazei de date Microlog
Fig.13. Tulpina FL20 crescuta pe mediu Lazareva 7 zile

Fig. 14. Coloniile de R. leguminosarum FL20 dupa 24 ore de incubare
Protocolul de lucru a fost urmatorul:
S-a insamantat pe mediul special BUG (Biolog Universal Growth) repartizat in placi Petri, una din culturile izolate anterior si s-a lasat la incubat 16-24 ore la 30C.
S-a calibrat sistemul si din placa Petri s-a pelevat cu o ansa sterila de unica folosinta putina masa bacteriana si s-a omogenizat in lichidul de inoculare specific (pentru bacterii gram negative).
S-a omogenizat masa bacteriana pana cand lichidul inoculat a atins o turbiditate standard, specifica de asemenea tipului gram negativ. Operatiunea s-a realizat cu atentie, si s-a verificat dupa fiecare omogenizare turbiditatea lichidului inoculat la Biolog.
Dupa ce s-a atins turbiditatea corespunzatoare lichidul inoculat s-a turnat intr-o cuva sterila, de unde, cu o micropipeta s-a insamantat microplaca GN Biolog cu cate 150 l suspensie in fiecare godeu.
Fig. 15. Suspensia bacteriana la turbiditatea standard pentru GN

Fig. 16. Inocularea microplacii GN

Fig. 17. Aspectul placii
GN pe care s-a efectuat mircroetalarea fenotipica

Citirea placilor s-a realizat cu un reader automat, iar interpretarea rezultatelor s-a facut cu ajutorul softului bazei de date pentru bacterii gram negative.
Identificarea si caracterizarea fenotipica au la baza reactiile din godeurile microplacii, respectiv sursele de carbon folosite de catre bacteria testata.
Godeurile in care sursele de carbon au fost utilizate de catre bacterie se coloreaza in violet. Reactia chimica are la baza prezenta in godeuri a unui colorant tetrazolic, care actioneaza ca un acceptor final de electroni in lantul respirator al bacteriei, formand un precipitat intens colorat de formazan.
Sursele de carbon din placa GN specifica bacteriilor gram negative sunt prezentate in tab. 12, iar in fig. 18 este prezentata citirea placii GN inoculata cu una din tulpinile de R. leguminosarum (FL20) folosita in cadrul proiectului.
Sistemul Biolog permite o caracterizare rapida a tulpinilor izolate si o verificare simpla a pastrarii caracteristicilor biologice dupa diferitele procedee de cultivare si de conditionare.
Tabel 12. Sursele de carbon din microplacile GN
|
A |
Apa |
Tween 80 |
N-acetil-D-galacto zamina |
N-acetil-D-gluco zamina |
N-acetil-b-D-mano zamina |
Adonitol |
Amigdalina |
D-arabi-noza |
L-arabi-noza |
D-arabitol |
Ar-butina |
D-celobioza |
|
B |
a-ciclo dextrina |
b-ciclo dextrina |
Dextrina |
i-Eritritol |
D-fructoza |
L-fructoza |
D-galactoza |
Acid-D- galactu ronic |
Gentio bioza |
Acid D-gluconic |
D-gluco zamina |
a-D-glucoza |
|
C |
a-D- glucoza-1-fosfat |
Glucurono-amida |
Acid D-glucuronic |
Glicerol |
Glicogen |
m-Inozitol |
Acid 2-cheto-D-gluconic |
a-D-lactoza |
Lac-tuloza |
Maltitol |
Maltoza |
Malto trioza |
|
D |
D-manitol |
D-manoza |
D-melezitoza |
Melibioza |
a-metil-galacto zida |
b-metil-galacto zida |
a-metil-D-glucozida |
b-metil-D- glucozida |
Palati noza |
D-Psicoza |
D-rafinoza |
L-ramnoza |
|
E |
D-riboza |
Salicin |
Sedo heptulozan |
D-sorbitol |
L-sorboza |
Stachioza |
Zaharoza |
D-tagatoza |
D-trehaloza |
Tura Noza |
Xilitol |
D-xiloza |
|
F |
Acid g-amino butiric |
Acid bromo Succinic |
Acid fumaric |
Acid b-hidroxi butiric |
Acid g-hidroxi butiric |
Acid p-hidroxifenil acetic |
Acid a-ketoglutaric |
Acid metil ester D-lactic |
Acid L-lactic |
Acid D-malic |
Acid L-malic |
Acid quinic |
|
G |
Acid D-zaharic |
Acid sebacic |
Acid succinamic |
Acid succinic |
Acid mono metil ester succinic |
Acid N-acetil L-glutamic |
L-alanin-amida |
L-alanina |
L-alanin-glicina |
L-aspara Gina |
Acid L-aspartic |
Acid L-glutamic |
|
H |
Acid glicil-L-glutamic |
L-ornitina |
L-fenil-alanina |
L-prolina |
Acid L-piro glutamic |
L-serina |
L-treonina |
2 amino etanol |
Putresce Ina |
Adeno Zina |
Uridina |
5-Mono fosfat adenozina |
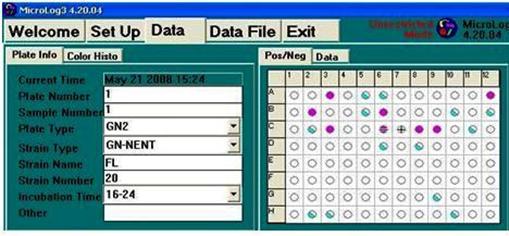
Fig. 18. Citirea placii GN inoculata cu tulpina R. leguminosarum FL20
IV.3. CULTIVAREA RHIZOBIILOR PE DIFERITE MEDII
S-a verificat faptul ca mediul Lazareva lichid poate fi folosit in scopuri industriale, deoarece asigura o crestere buna a tuturor rizobiilor si permite realizarea de sarje reproductibile, atat din punct de vedere al timpului de fermentare, cat si al cantitatii de masa bacteriana realizata.
Tabel 13. Biomasa umeda rezultata dupa 4 zile, in urma cultivarii pe diferite medii de cultura inoculate cu tulpini izolate din nodozitati (g/l mediu) .
|
Tulpina |
Fred si Waksman |
Bond |
Hendrikson |
Van Schreven |
Vincent |
Burton |
Wright |
Lazareva |
|
FL 20 | ||||||||
|
MAP 48 | ||||||||
|
TAP 21 | ||||||||
|
SOAP 10 |
1,6 |
Tabel 14. Biomasa umeda rezultata dupa 7 zile, in urma cultivarii pe diferite medii de cultura inoculate cu tulpini izolate din nodozitati (g/l mediu).
|
Tulpina |
Fred si Waksman |
Bond |
Hendrikson |
Van Schreven |
Vincent |
Burton |
Wright |
Lazareva |
|
FL 20 | ||||||||
|
MAP 48 | ||||||||
|
TAP 21 | ||||||||
|
SOAP 10 |
5,0 |
Tulpinile de Rhizobium care au fost cultivate pe mediu lichid Lazareva conditionate cu Aquasorb in pungile urinare sterile su crescut normal si si-au pastrat viabilitatea.

Fig. 19. Aspectul unui pungi cu mediu de cultura hidrogelifiat
Testele preliminare realizate au demonstrat ca hidrogelul utilizat nu sufera modificari in urma sterilizarii prin radiatii si ca bacteriile din grupul Rhizobium cresc pe mediile hidrogelifiate. In fig. 19 este prezentat aspectul pungii cu hidrogel, iar in fig. 20 este ilustrata dinamica de crestere a rizobiilor pe mediu hidrogelifiat.
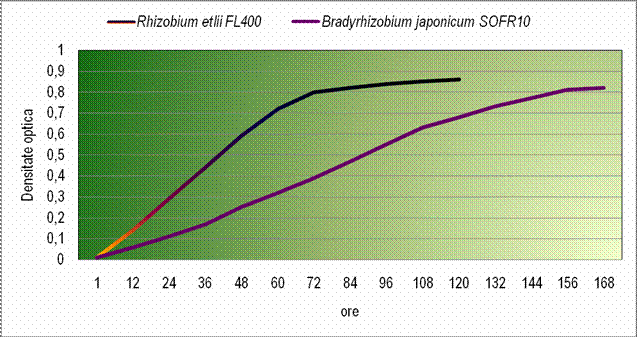
Fig. 20. Dinamica de crestere a rhizobiilor pe mediu hidrogelifiat.
IV.4. VERIFICAREA PASTRARII CALITATIILOR BIOLOGICE ALE RHIZOBIILOR
In cazul rhizobiilor nu au fost evidentiate modificari semnificative ca urmare a cultivarii pe medii hidrogelifiate. In fig. 21 sunt prezentate comparativ activitatea nitrogenazica (prin estimarea hidrogenului nitrogenazic) a nodozitatilor rezultate in urma inocularii plantelor de fasole cu bacterii Rhizobium leguminosarum bv. phaseoli, tulpina FL20, cultivata pe mediu Lazareeza, hidrogelifiat si agarizat.
A B C
Fig. 21. Activitatea nitrogenazica a radacinilor de fasole inoculate cu rhizobii cultivate pe medii Lazareeva. A. hidrogelifiat B. agarizat. C. martor neinoculat.
Datele obtinute demonstreaza faptul ca nu se constata diferente semnificative in cazul activitatii biologice a rhizobiilor cultivate pe medii hidrogelifiate sau agarizate. Preturile de cost ale hidrogelului fiind mult mai reduse decat cele ale agarului sunt astfel create conditiile pentru realizarea unor biopreparate pe medii semisolide, (hidro)gelifiate.
Aceste medii gelifiate au fost cele mai utilizate pentru producerea biopreparatelor pe baza de rhizobii. Aceste biopreparate sunt cele mai larg folosite si cele mai vechi. La cativa ani dupa descoperirea simbiozei dintre rhizobii si leguminoase de catre Hellriegel (1889) doi alti cercetatori germani (Nobbe si Hiltner) au obtinut patentul SUA 570 813/1895 pentru primul biopreparat pe baza de fixatori de azot in simbioza cu leguminoasele. Acest biopreparat continea o tulpina de bacterii specifica pentru cultura unei anumite leguminoase, inmultita pe un mediu artificial solidificat cu gelatina pe peretii unui flacon de sticla cu capacitatea de 250 ml. In urma criticilor unor utilizatori (care pretindeau ca gelatina, fiind o sursa semnificativa de azot fixat, inhiba capacitatea de fixare biologica a azotului) componenta de gelifiere a mediului de cultura a fost schimbata cu agar, un polizaharid algal care prezinta aceeasi caracteristica de gelifiere functie de temperatura ca si gelatina. S-a obtinut astfel un produs a carui denumire comerciala (Nitragin, detinuta in prezent de Merck) a fost inregistrata la data de 6 decembrie 1898. Desi este un nume comercial inregistrat ca marca (adica un substantiv propriu) Nitragin a devenit in limba romana, ca si in alte cazuri (ness, xerox, adidasi sau, mai nou, pampersi) un nume (substantiv) comun, fiind utilizat ca denumire generica pentru biopreparatele cu rhizobii.
Inoculantii agricoli pe mediu agarizat prezentau insa o serie de dezavantaje (pretul de cost ridicat datorita pretului mare al agarului, termenul de valabilitate realtiv scazut care impunea o activitate sezoniera de productie, manopera multa la producere) fapt care a determinat cautarea unor solutii alternative. In 1907 s-a comercializat biopreparatul Azotogen, in care rhizobiile erau inmultite direct in borcane de pamant neutralizat cu praf de calcar. Un an mai tarziu (1908) Coates a obtinut un brevet pentru un biopreparat cu rhizobii care continea bacteriile adsorbite pe faina sterilizata din plante de trifoi, mazare, fasole sau seminte de bumbac. Doi ani mai tarziu a obtinut altul in care propunea faina de oase ca suport de crestere si conditionare pentru rhizobii. Ale suporturi au fost propuse de Diller, in 1920 (bioxidul de siliciu, carbonatul de calciu, fosfatul de calciu) si de Dugger in 1924 (compost, negru de fum, bentonita). Nici unul dintre suporturile de conditionare nu a cunoscut insa succesul turbei, propusa pentru prima oara in 1932 ca suport de conditionare a rhizobiilor. Tehnologia de producere a biomasei de rhizobii si de utilizare a turbei ca suport de conditionare a fost perfectionata in anii '60 de australieni, si apoi de britanici si de francezi.
Desi foarte larg utilizate aceste biopreparate inoculante pe turba prezinta si o serie de dezavantaje. Un prim dezavantaj este legat de conditiile de productie si se refera la competititivitatea din ce in ce mai redusa a tulpinilor utilizate. Prima etapa in obtinerea biopreparatelor pe turba este producerea biomasei pe mediu lichid, aerat si agitat. Pentru a corespunde cerintelor acestei etape s-au eliminat din productia de inoculanti agricoli tulpinile cu crestere mai lenta si producere masiva de exopolizaharid, adica tocmai tulpinile cu competitivitate ridicata si activitati biologic mare.
Un alt dezavantaj practic al biopreparatelor pe turba este legat de conditiile de aplicare, care solicita manopera suplimentara pentru descarcarea sacilor cu samanta, umectarea semintelor cu solutii de adezivi (care necesita conditii speciale de preparare, fiind de obicei hidrocoloizi dificil de dispersat in apa), tratarea semintelor cu biopreparat pe turba, reinsacuire, transport la camp. In plus, in cazul anumitor tipuri de semanatori (si in special a celei romanesti SPC), exhaustorul aspira turba de pe samanta de leguminoase.
Pentru a contracara aceste dezavantaje au fost dezvoltate in ultimii 15 ani noi tipuri de biopreparate inoculante. Doua directii principale au fost urmate: (i) producerea de biopreparate pe medii gelificate (biopreparate care sunt solide in etapele de producere, transport, receptie si lichide in etapa de utilizare) si (ii) producerea de biopreparate cu eliberare controlata, care permit o mai buna asigurare a colonizarii.
Gelificarea mediilor de cultura prin adsorbtia unui mediu de cultura specific intr-un hydrogel cu capacitate de gonflare mare. Capacitatea de gonflare a hidrogelului superadsorbant variaza intre 50 si 200 ori masa initiala, ceea ce inseamna max. 20 g pentru 1 kg (aprox . 1 litru) de mediu. Aceasta capacitate de gonflare este controlabila prin concentratia initiala de saruri ale acidului acrilic folosita pentru polimerizare, prin doza/concentratia de initiator de polimerizare si prin tipul si marimea cationului destinat neutralizarii polianionului acrilat. Este de mentionat aici ca hidrogelurile superadsorbante acrilat-amidon sunt larg utilizate in adsorbenti de igiena personala, dar nu sunt inca utilizate ca gelifianti pentru biopreparate. Hidrogelurile (super)adsorbante acrilat - poliacrilamida sunt: (i) compatibile cu mijloacele uzuale de sterilizare (autoclavare 30 min la 121 C, sterilizare cu radiatii gamma si/sau oxid de etilena); (ii) neutre fata de rhizobii, permitand dezvoltarea microorganismelor la suprafata lor ca si mediile agarizate; (iii) lipsite de toxicitate pentru om, plante sau alte organisme netinta din mediu; (iv) lent biodegradabile (ca si agarul). Desi mai greoaie decat solutia deja clasica a gelifierii cu agar folosirea hidrogelurilor (super)adsorbante are avantajul de fi mult mai ieftina. Daca 1 kg de agar, necesar pentru gelifierea a 50 litri de mediu, costa peste 250 EUR depozit furnizor (pret pentru agar industrial, de calitate redusa valabil numai pentru comenzi mari), pretul unui kg de hidrogel superadsorbant acrilat/ amidon nu depaseste 5 dolari SUA.
IV.5. EVIDENTIEREA FENOMENULUI DE QS LA BACTERIILE INOCULANTE
In fig. 22 este reprezentat apsectul tulpinii indicator de Chromobacterium violaceum Cv 026, folosita in cadrul experimentelor de evidentiere a QS la rhizobiile studiate.

In fig. 23 este prezentat aspectul bacteriiilor biosenzor in prezenta tulpinilor de rhizobii producatoare de AHL.
|
|
|
|
A |
b |
Fig. 23. Aspectul bacteriilor biosenzor (a- Chromobacterium violaceum Cv026; b - Agrobacterium tumefaciens WCF47) in prezenta tulpinilor producatoare de AHL.
Dupa incubarea peste noapte bacteriile biosenzor din cel de-al doilea strat au dezvoltat o reactie de culoare specifica. In cazul rizobiilor care au si o activitate de favorizare a cresterii si dezvoltarii plantelor s-au folosit ca tulpini indicator / biosenzor Chromobacterium violacearum Cv026 (evidentiindu-se producerea de violaceina), si tulpina de Agrobacterium tumefaciens WCF47 (la care moleculele de N-acil-homoserin-lactona, AHL, sunt detectate prin reactia cromogenica a b-galactozei raportor cu substratul X-gal, care genereaza o culare albastra).
IV.6. CARACTERIZAREA UNOR TULPINI DE RHIZOBIUM CARE AU ACTIVITATE ACC DEAMINAZICA SI CARE NODULEAZA FASOLEA
Un aspect esential in cadrul studiilor de izolare a tulpinilor de rhizobii care au activitate ACC deaminazica si care noduleaza fasolea este cel metodologic, respectiv cel al modului in care sunt obtinute mediile selective.
Procedeul descris in literatura prevede cultivarea pe mediu minimal DF care are urmatoarea compozitie: glucoza 2g/l, acid gluconic 2g/l, acid citric 2g/l, KH2PO4 4g/l, MgSO4 0.2g/l, NaCl 0.1g/l, pH 6.8, solutie micronutrienta 10 ml , Agar 18g/l.
Solutia micronutriente: CaCl2 200mg/l, FeSO4 200mg/l, H3BO3 15mg/l, ZnSO4 20mg/l, Na2MoO4 10mg/l, KI 10mg/l, NaBr 10mg/l, MnCl2 10mg/l, CoCl2 5mg/l, CuCl2 2mg/l, AlCl3 2mg/l, NiSO4 2mg/l.
Pentru a se evita formarea unor compusi greu solubili, degradarea ACC sau impiedicarea gelifierii s-a definitivat prin incercari repetate, urmatorul procedeu de lucru:
solutia de microelemente (cu exceptia FeSO4.7H20) s-a dizolvat in apa distilata sterila, s-a adus la semn la cotat de 100 ml, s-a transvazat in flacon de sticla bruna si s-a depozitat la rece;
solutia de sulfat de fer (1 mg / 10 ml) s-a preparat separat, s-a transvazat in flacon de sticla bruna si s-a depozitat la rece;
mediul s-a preparat la 1 l, prin dizolvarea pe rand a fiecarui ingredient ( nu s-a trecut la adaugarea unui nou ingredient pana cand cel anterior nu era complet dizolvat). Ordinea de adaugare a fost: KH2PO4 4g/l, MgSO4 0,2g/l, NaCl 0,1 g/l glucoza 2g/l, acid gluconic 2g/l, acid citric 2g/l, 0,1 ml solutie microelemente, 0,1 ml solutie sulfat de fer, agar 18 g/l/
Sterilizarea se face la 121 C pentru max. 20 min.
ACC se prepara ca solutie 0,5 M, se sterilizeaza prin filtrare pe membrana de 0,2 mm si se pastreaza la - 20 C. Solutia sterila de ACC se adauga chiar inainte de distribuirea mediului in placi, in proportie 300 ml la 50 ml mediu.
In cadrul acestui experiment privind activitatea ACC-deaminazica, au fost testate o serie de tulpini de Rhizobium care noduleaza fasolea, dintre are doar tulpinile R. leguminosarum bv. phaseoli FL220 si Rhizobium spp. FL400 au manifestat aceasta activitate.
In urma izolarilor facute din nodulii de culoare roz formati pe radacinile plantulelor de mazariche, prezenta tulpinilor de Rhizobium s-a evidentiat pe mediul de cultura manitol-rosu de Congo, prin culoare roz a coloniilor bacteriene (fig. 24). Doar la tulpinile FL400 si FL220, in urma cresterii pe mediile DF lichid si DF solid suplimentate cu ACC ca unica sursa de azot s-a evidentiat manifestarea activitatii ACC deaminazice, in timp ce tulpina FL20 nu a crescut pe aceste medii specifice.
Cresterea tulpinilor selectate pe mediu minimal DF suplimentat cu ACC a fost slaba, ceea ce dovedeste ca aceste tulpini au o activitate ACC-deaminazica redusa.
Fig.24. Coloniile de FL20, FL400 si FL220
Bacteriile selectionate prezinta insa o activitate semnificativa de stimulare a cresterii plantulele test. Aceasta stimulare implica si activitatea ACC-deaminazica, care inhiba producerea de etilen (fitohormon care inhiba cresterea) in tesuturile vegetale.
Fig. 25.
Semintele de Vicia villosa germinate pe mediu agarizat

Fig. 26. Plantulele de Vicia villosa dupa inoculare cu tulpinile de testat, pe mediu
semilichid Crone

Aceasta activitate de fitostimulare a tulpinilor izolate fost evidentiata si prin stimularea cresterii plantulelor in vitro. Rezultatele acestei biotestarii in vitro a efectului fitostimulator al tulpinilor cu activitate ACC-deaminazica (suspendate in mediu de cultura Lazareeva) pentru stimularea cresterii plantelor sunt prezentate in tabelul 15.
Tabel 15. Efectul fitostimulator in vitro al tulpinilor de Rhizobium cu activitate ACC-deaminazica.
|
Varianta experimentala |
Lungime radacini* (cm) |
|
|
Rapita cv. Triangle |
Vicia villosa |
|
|
Rhizobium leguminosarum bv. phaseoli FL 220 |
1,42 a |
1,17 ab |
|
Rhizobium spp FL400 |
1,12 ab |
0,84 b |
|
Martor mediu de cultura |
1,22 b |
0,93 a |
|
Martor netratat |
0,42 c |
0,64 c |
*Valorile urmate de aceeasi litera nu difera semnificativ statistic pentru p<0.05
Lungimea radacinilor prezentata in tabelul 15 este cea a plantulelor formate din seminte de rapita (cv. Triangle) si de mazariche paroasa, sterilizate la suprafata (prin spalari repetate cu Domestos si clatiri cu apa distilata) si inoculate cu 0,1 ml suspensie de bacterii cu activitate ACC-deaminazica in mediu Lazareeva. Martorul mediu de cultura a fost mediul Lazareeva. Datele obtinute demonstreaza un efect stimulator in vitro semnificativ, asigurat statistic, al tulpinilor cu activitate ACC-deaminazica.
Tulpinile au fost caracterizate prin microetalare fenotipica in placile GN Biolog. In cazul tulpinii FL220, s-a confirmat prin microetalare fenotipica, apartenenta acesteia la specia R. leguminosarum biovar. phaseoli. In cazul tulpinii FL400 nu s-a confirmat incadrarea taxonomica prin microetalarea fenotipica, fiind necesare studii suplimentare.
IV.7. ESTIMAREA EFICACITATII FIXARII SIMBIOTICE A AZOTULUI PRIN DETERMINAREA HIDROGENULUI
In fig. 27 este prezentat dispozitivul experimental utilizat pentru estimarea eficacitatii fixarii simbiotice a azotului prin determinarea hidrogenului produs de nitrogenaza.

Fig. 27. Dispozitivul de laborator pentru masurarea hidrogenului nitrogenazic
Rezultatele preliminare obtinute demonstreaza functionalitatea metodei de estimare a eficacitatii de fixare a azotului prin determinarea hidrogenului nitrogenazic.
Traditional, fixarea N2 a fost masurata prin metoda reducerii acetilenei, care necesita utilizarea unui gaz-cromatograf pentru a masura reducerea acetilenei la etilena de catre organismele fixatoare de azot. Gaz-cromatografele sunt scumpe, iar o concetratie de 10% de acetilena in mediu de reactie necesara metodei este exploziva. Pe langa acest motiv excelent pentru evitarea metodei, s-a dovedit recent ca reducera acetilenei inhiba reactia nitrogenazei.
In fig. 28 sunt prezentate activitatile nitrogenazice ale celor doua tulpini de Rhizobium care noduleaza fasolea si prezinta activitate ACC-deaminazica.

Fig. 35. Activitatea nitrogenazica determinata prin producerea de hidrogen de catre sistemul nitrogenazic.
Datele obtinute reliefeaza functionalitate metodei, ca si faptul ca, in conditiile experimentale date, tulpinile cu ACC-deaminazica au si o activitate nitrogenazica semnificativa.
IV.8. CULTIVAREA UNEI TULPINI DE AZOSPIRILLUM BRASILIENSE PE DIFERITE MEDII
Datele obtinute in urma cantaririi biomasei bacteriene de Azospirillum brasiliense tulpina Sp001 rezultata in urma cultivarii pe cele 3 medii de crestere, precum si a masurarii densitatii optice / turbiditatii sunt prezentate in tabelul de mai jos:
Tablel 15. Acumularea biomasei de A. brasilene in mediile lichide testate, (masa medie a sedimentului bacterian) si a turbiditatii (NTU) (datele reprezinta media a trei repetitii)
|
Varianta (mediu) |
Greutate sediment (g/l) |
pH-ul |
densitate turbidimetrica (NTU) |
||||||
|
24h |
48h |
72h |
0h |
24h |
48h |
24h |
48h |
72h |
|
|
AFM10 | |||||||||
|
AFM 20 | |||||||||
|
Dobereiner | |||||||||
Se remarca faptul ca, pe mediile testate, cresterea bacteriilor din genul Azospirillum se plafoneaza dupa 72 ore. Deoarece aceasta plafonare nu este datorata de epuizarea surselor de azou si/sau carbon, este de presupus ca inhibarea dezvoltarii bacteriilor dupa 72 de ore se datoreaza acumularii metabolitilor de excretie bacterieni.
Randamentele de conversie ale componentelor mediului de cultura in biomasa (tab. 16), sunt in general ridicate. Mediul care asigura conversia cea mai ridicata este mediul AFM 10. Totusi din punct de vedere practic sunt mai convenabile mediul AFM 20 si mediul Dobereiner.
Tabel 16. Randamentele de conversie a componentelor mediului de cultura
|
Mediul |
Randament de conversie la: |
||
|
24h |
48h |
72h |
|
|
Mediul AFM 10 | |||
|
Mediul AFM 20 | |||
|
Mediul Dobereiner | |||
Mediul AFM 20 asigura un nivel rezidual de nutrienti care permite o supravietuire optima in cadrul etapelor de conditionare. Mediul Dobereiner este un mediu semisolid, care are ca principal avantaj practic selectivitatea - fapt care permite in procesul de tehnologie evitarea contaminarilor.
In concluzie, se propune ca mediul AFM 20 sa fie utilizat in procesele tehnologice de biosinteza a biomasei de bacterii Azospirillum brasiliense SP001, iar mediul Dobereiner sa se foloseasca pentru mentinerea culturilor si pentru prepararea culturilor starter.
IV.9. TESTAREA ACTIVITATI BIOLOGICE A BIOMASEI DE AZOSPIRILLUM
Efectuarea biotestului producerii de gibereline in laborator, a permis evaluarea influentei exercitate de mediile de cultura asupra calitatii de favorizant de crestere a tulpinii testate.
Tabel 17. Influenta mediilor de cultura asupra calitatilor biologice a tulpinii SP001
|
Varianta experimentala |
Media diametrelor zonelor de difuzie (mm) |
Conc. Echivalenta GA3 (ng/ml) |
|
M1 - extract de AFM 10 | ||
|
M2 - extract de AFM 20 | ||
|
M3 - extract de mediu Dobereiner | ||
|
V1 - extract de cultura pe mediu AFM 10 | ||
|
V2 - extract de cultura pe mediu AFM 20 | ||
|
V3 - extract de cultura pe mediu Dobereiner | ||
|
DL 5% |
Datele din tabel aduc dovezi ale existentei fitohormonilor giberelinici in extractele analizate. Diametrele zonelor de difuzie ale enzimelor giberelic induse sunt semnificativ mai mari la variantele incubate in prezenta mediilor de cultura. Toate mediile testate asigura o capacitate ridicata de biosinteza a fitohormonilor giberelinici, comncentratia acestora in mediul de cultura depasind 100 ng echivalenti GA3/ ml.
Mediul de cultura care a asigurat bacteriilor cea mai ridicata capacitate de a sintetiza fitohormoni giberelinici a fost mediul AFM 20.
Rezultatele obtinute demonstreaza faptul ca se pastreaza calitatea biologica de agent favorizant al cresterii vegetale a bacteriilor A. brasiliense Sp001, dupa cultivarea pe mediile AFM 10, AFM 20 si Dobereiner.
IV.10. CONDITIONAREA TULPINILOR BACTERIENE FL220 SI Sp001 SUB FORMA DE AGREGATE - FLOCULI
Pentru asigurarea unui inocul bogat in bacterii cu mobilitate ridicata, s-a realizat un procedeu tehnologic de co-floculare a bacteriilor agroinoculante din grupurile Azospirillum si Rhizobium cultivate pe mediu lichid, aerat si agitat.
Procedeul elaborat permite obtinerea unui inocul bogat in bacterii (min 1011 ufc/ g de produs final) capabil sa colonizeze rapid rizosfera plantelor de cultura.
In fig. 36 sunt prezentate aspecte din timpul conditionarii tulpinilor FL220 sub forma de agregate / floculi.
Procedura de lucru utilizata pentru realizarea acestui biopreparat co-floculat este urmatoarea:
Cultivarea bacteriilor din grupul Rhizobium (R. leguminosarum bv. phaseoli izolatul FL220 ) pe mediu Lazareva hidrogelifiat timp de 7 zile.
Resuspendarea inoculului in mediu AFM 20S, ce contine acid malic in loc de glucoza, in proportie de 10% (s-au inoculat 10 ml de suspensie bacteriana in 100 ml mediu);
Cultivarea bacteriilor din grupul Azospirillum (Azospirillum brasiliense Sp001), in mediu lichid AFM20 aerat si agitat, timp de 2 zile. Agitarea a avut loc la 140 rpm, in vederea formarii de flocoane;
Normalizarea suspensiilor de bacterii Rhizobium si Azospirillum, la o densitate optica de circa 0,4. Aceasta densitate optica corespunde unei suspensii cu titrul de 108 ufc/ml.
Realizarea co-inocularii in raport de 1:1 pe mediu AFM 20S;
Cultivarea peste noapte, recoltarea biomasei floculante prin decantarea si uscarea acestei biomase in curent de aer in conditii blande (max. 35C).
a

b

Fig. 37. Aspecte din timpul conditionarii sub forma de agregate / floculi. a) tulpina de Rhizobium leguminosarum bv. phaseoli FL220 pe mediul solid Lazareva si mediul lichid AFM S; b) tulpina de Azospirillum brasiliense Sp001 pe mediul semisolid Dobereiner si mediul lichid AFM 20.
Prin aplicarea procedeului rezulta un biopreparat mixt sub forma de floculi in care rata de supravietuire este buna (fig. 38). Pentru cultivarea bacteriilor Azospirillum brasilense s-au folosit mediul lichid AFM20 si mediul semisolid Dbereiner, ale caror formule au fost prezentate deja. Mediul Dobereiner a fost folosit pentru mentinerea bacteriilor, iar mediul AFM 20 pentru producerea biomasei de azospirillii necesara pentru conditionare.
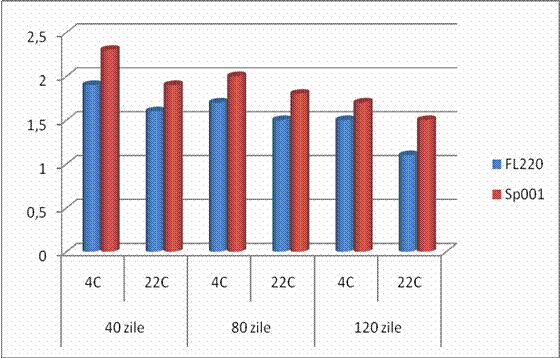
Fig. 38. Supravietuirea bacteriilor in bioprepatul mixt pe baza de Azospirillum brasilense Sp001 (x109 ufc/g) si Rhizobium leguminosarum bv. phaseoli FL 220 (x107 ufc/g). Estimari ale numarului de microorganisme prin dilutii succesive si cultivare pe medii specifice, Lazareeva cu rosu de Congo si Dobereiner.
Fig.39
Biopreparat mixt Azospirillum SP001 si Rhizobium FL220 conditionat sub forma de
floculi


Tab. 18. Influenta biopreparatului mixt FL220-Sp001 asupra fenomenelor de fitostimulare a plantelor de rapita si mazariche
|
Varianta experimentala |
Lungime* radacini (cm) |
|
|
Rapita cv. Triangle |
Vicia villosa |
|
|
Martor netratat, neinoculat |
0,75 c |
0,32 c |
|
Tratat suspensie 109 ufc/ml Azospirillum brasilense Sp001 |
0,62 b |
0,97 b |
|
Tratat suspensie 107 ufc/ml Rhizobium leguminosarum bv. phaseoli FL220 |
0,54 b |
1,31 b |
|
Biopreparat Azospirillum brasilense Sp001 + Rhizobium leguminosarum bv. phaseoli FL220 |
2,07 ab |
1,83 ab |
Datele din literatura arata un efect sinergic al co-inocularii plantelor de leguminoase cu cele doua tipuri de fixatori de azot. In cadrul acestei lucrari s-a urmarit influenta exercitata de acest biopreparat mixt asupra proceselor de stimualre a cresterii (tab.18), intrucat amandoua tulpinile prezentau si activitate de stimulare a cresterii
Datele arata un efect aditiv, la limita sinergismului. Sunt necesare studii suplimentare pentru stabilirea pastrarii acestei interactii sinergice pe parcursul pastrarii biopreparatului, ca si al efectelor asupra fixarii biologice de azot.
CAPITOLUL V. CONCLUZII
Au fost izolate mai multe tulpini de Rhizobium folosindu-se tehnica capturii pe radacini de leguminoase si a recuperarii tulpinilor din nodulii eficienti (de culoare roz) pe mediul Lazareva cu rosu de Congo. Din plantele de soia au fost izolate si purificate 11 tulpini, din trifoi s-au obtinut 12 izolate, de la fasole 17 izolate, iar de la mazare 9 izolate. S-a constatat ca viteza de crestere pe mediul Lazareva a fost de 4 zile pentru izolatele de la fasole, mazare si trifoi si de 7 zile pentru cele de la soia.
Tulpinile izolate au fost selectionate dupa capacitatea lor de crestere pe diferite medii de cultura, in final optandu-se pentru mediu Lazareeva si o serie de tulpini de rhizobii care noduleaza fasolea.
A fost pusa la punct o procedura de caracterizare fenotipica a rhizobiilor prin utilizarea (micro)sistemului Biolog. Procedura porneste de la modul de lucru recomandat de producatori si este adaptata conditiilor laboratorului in care s-a lucrat si specificului bacteriilor studiate.
Mediu Lazareeva a fost gelifiat prin folosirea a doi compusi macromoleculari, agarul (compus utilizat traditional) si hidrogelul pe baza de poliacrilamida - poliacrilat (Aquasorb).
Tulpinile de rhizobii cultivate pe medii hidrogelifiate si-au pastrat caracteristicile biologice de fixare a azotului atmosferic.
S-a evidentiat producerea de AHL si prezenta sensibilitatii de grup (quorum sensing) la tulpinile de Rhizobium. S-au folosit ca tulpini indicator / biosenzor Chromobacterium violacearum Cv026 (evidentiindu-se producerea de violaceina), si tulpina de Agrobacterium tumefaciens WCF47 (la care moleculele de N-acil-homoserin-lactona, AHL, sunt detectate prin reactia cromogenica a b-galactozidazei raportor cu substratul X-gal, care genereaza o culoare albastra).
S-a elaborat un procedeu de selectare a tulpinilor de Rhizobium cu activitate ACC-deaminazica prin crestere pe mediu minimal DF cu ACC ca unica sursa de azot. Pentru a se evita formarea unor compusi greu solubili, degradarea ACC sau impiedicarea gelifierii s-a definitivat prin incercari repetate, urmatorul procedeu de lucru:
solutia de microelemente (cu exceptia FeSO4.7H20) s-a dizolvat in apa distilata sterila, s-a adus la semn la cotat de 100 ml, s-a transvazat in flacon de sticla bruna si s-a depozitat la rece;
solutia de sulfat de fer (1 mg / 10 ml) s-a preparat separat, s-a transvazat in flacon de sticla bruna si s-a depozitat la rece;
mediul s-a preparat la 1 l, prin dizolvarea pe rand a fiecarui ingredient ( nu s-a trecut la adaugarea unui nou ingredient pana cand cel anterior nu era complet dizolvat). Ordinea de adaugare a fost: KH2PO4 4g/l, MgSO4 0,2g/l, NaCl 0,1 g/l glucoza 2g/l, acid gluconic 2g/l, acid citric 2g/l, 0,1 ml solutie microelemente, 0,1 ml solutie sulfat de fer, agar 18 g/l/
Sterilizarea se face la 121C pentru max. 20 min.
ACC se prepara ca solutie 0,5 M, se sterilizeaza prin filtrare pe membrana de 0,2 mm si se pastreaza la - 20 C. Solutia sterila de ACC se adauga chiar inainte de distribuirea mediului in placi, in proportie 300 ml la 50 ml mediu.
Doar la tulpinile FL400 si FL220, in urma cresterii pe mediile DF lichid si DF solid suplimentate cu ACC ca unica sursa de azot s-a evidentiat manifestarea activitatii ACC deaminazice, in timp ce alte tulpini nu au crescut pe aceste medii specifice.
Bacteriile selectionate ca avand activitate ACC-deaminazica prezinta o activitate semnificativa de stimulare a cresterii plantulele test (rapita si mazariche). Aceasta stimulare implica pe langa producerea de fitohormoni in situ si activitatea ACC-deaminazica, care inhiba producerea de etilen (fitohormon care inhiba cresterea) in tesuturile vegetale.
A fost pusa la punct o procedura de lucru pentru estimarea activitatii nitrogenazice prin determinarea producerii de hidrogen in conditiile utilizarii de amestecuri gazoase lipsite de azot (ca de exemplu argon : oxigen 80 : 20). Datele obtinute reliefeaza functionalitate procedurii puse la punct, ca si faptul ca, in conditiile experimentale date, tulpinile cu ACC-deaminazica au si o activitate nitrogenazica semnificativa.
In urma studiile de cultivare s-a decis ca mediul AFM 20 sa fie utilizat in procesele tehnologice de biosinteza a biomasei de bacterii Azospirillum brasiliense SP001, iar mediul Dobereiner sa se foloseasca pentru mentinerea culturilor si pentru prepararea culturilor starter.
S-a realizat un biopreparat mixt pe baza unui consortiu de bacterii fixatoare de azot, Azospirillum brasiliense Sp001 si Rhizobium leguminosarum bv. phaseoli FL 220. Biopreparatul s-a realizat prin co-floculare. Datele experimentale obtinute arata un efect aditiv, la limita sinergismului. Sunt necesare studii suplimentare pentru stabilirea pastrarii acestei interactii sinergice pe parcursul pastrarii biopreparatului, ca si al efectelor asupra fixarii biologice de azot.
CAPITOLUL VI. BIBLIOGRAFIE
1. ABDEL-SALAM, M. S. , KLINGMOLLER, W. -1988. Isolation of transposon induced auxin-negative mutations in Azospirillum lipoferum. In Azospirillum. IV. Genetics, physiology , ecology. Edited by W. Klingmuller. Springer- Verlag , Berlin, Heidelberg, pp. 40-48.
2. ABDEL-SALAM, M. S. , KLINGMULLER, W. 1987. Transposon Tn5 mutagenesis in Azospirillum lipoferum: isolation of indoleacetic acid mutants. Mol. Gen. Genet. 210: 165-170.
3. ALBRECHT, S. L., GASKINS, M. H., MILAM, J. R., SCHANK, S.C., SMITH, R. L. 1983. Ecological factors affecting survival and activity of Azospirillum in the rhizosphere. In Azospirillum. II. Experientia Suppl.48: 138-148.
4. ALBRECHT, S. L., OKON, Y., LONNQUIST, L. , BURRIS, R. H. 1981. Nitrogen fixation by corn-Azospirillum associations in a temperate climate. Crop Sci. 21: 301-306.
5. Antoun, H., Beauchamp, C.J., Goussard, N., Chabot, R., Lalande, R., 1998, Plant and Soil, 204, 57-67
6. Appels, M.A., 1989, The symbiosis between Rhizobium leguminosarum and Pisum sativum: regulation of nitrogenase activity, 138 pp.
7. Appleby, C.A., 1984, Ann. Rev. Plant Physiol., 35, 433-478.
8. AVlVI, Y., FELDMAN, M. 1982. The response of wheat to bacteria of the genus Azospirillum. Isr. J. Bot. 32: 237-245.
9. Andrighetti-Frhner, C.R., Antonio, R.V., Creczynski-Pasa, T.B., Barardi, C.R.M., Simoes, C.M.O. 2003. Cytotoxicity and potential antiviral evaluation of violacein produced by Chromobacterium violaceum. Mem. Inst. Oswaldo Cruz, 98(6): 843-848.
10. Bassler, B.L. 2002. Small talk. Cell-to-cell communication in bacteria. Cell, 109: 421-424.
11. Bassler B.L., Losick, R. 2006. Bacterially speaking. Cell, 125: 237-246.
12. Bains W. (2004): Biotechnology, University Press, Oxford.
13. Berner D.J., Krieg N.R., Stanley J.T (editors)
(2005): Bergey![]() s
Manual of Systematic Bacteriology (second edition), Springer Science and
Business Media Inc., New York.
s
Manual of Systematic Bacteriology (second edition), Springer Science and
Business Media Inc., New York.
14. Bitton G. (2002): Encyclopedia of environmental microbiology, University of Florida.
15. Bull A.T. (2004): Microbial diversity and bioprospecting, ASM press, Washington.
16. Bejerano-Sagie, M., Xavier, K.B. 2007. The role of small RNAs in quorum sensing. Curent Opinion in Microbiology, 10:189-198.
17. CHI, N.T.P., THANH, H.H., DZUNG, N.N., MIRZA, M.S., LADHA, J.K., 1998, Responses of rice plants to inoculation with Azospirillum sp. under field conditions. In: Proc. 7th Int. Symp. with Nitrogen Fixation with Non-Legumes, 16-21 Oct.1986, Faisalabad, Pakistan, 1998, 237-240.
18. Ceccatto, V.M., Gomes, J.E., Sarris, G.A., Moon, D.H., Tsai, S.M., 1998, Plant and Soil., 79-87.
19. Daniels, R., De Vos, D.E., Desair, J., Raedschelders, G., Luyten, E., Rosemeyer, V., Verreth, C., Schoeters, E., Vanderleyden, J., Michiels, J. 2002. The cin quorum sensing locus of Rhizobium etli CNPAF512 affects growth and symbiotic nitrogen fixation. J. Biol. Chem. 277:462-468.
20. DE-POLLI, H., BOHLOOL, B. B., and DOBEREINER., J. 1980. Serological differentiation of Azospirillum species belonging to different host plant specificity groups. Arch. Microbiol.126:217-222.
21. DOBEREINER., J., 1988. Isolation and identification of root associated diazotrophs. Plant Soil, 110: 207-2l2.
22. DOBEREINER., J., BALDANI,V. L. D. 1979. Selective infection of maize roots by streptomycin-resistant Azospirillum lipoferum and other bacteria. Can. J. Microbiol. 25: 1264-1269.
23. DOBEREINER., J., MARRIEL, I. E., NERY, M. 1976. Ecological distribution of Spirillum lipoferum Beijerinck. Can. J. Microbiol. 22: 1464-1473.
24. Dreyfus, B., Garcia, J.L., Gillis, M., 1988, Int. J.,Syst. Bacteriol., 38, 89-98.
25. DROZDOWICZ, A. , FERREIRA SANTOS, G. M. 1987. Nitrogenase activity in mixed cultures of Azospirillum with other bacteria. Zentralbl. Mikrobiol.142: 487-493.
26. Dunca S., Octavita A., Stefan M. (2004): Microbiologie aplicata, Editura Tehnopress, Iasi.
27. Duta,F.P., de Frana,F.P., de Almeida Lopes,L.M., 2006, Optimization of culture conditions for exoplysaccharides production in Rhizobium sp. Using the response surface method, Electronic J. Biotechnology.
28. Eliade Gh., Ghinea L., Stefanic Gh. (1975): Microbiologia solului, Editura Ceres, Bucuresti.
29. FALLIK, E., OKON, Y., EPSTEIN, E., GOLDMAN, A., FISCHER, M. 1989. Identification and quantification of IAA and IBA in Azospirillum brasilense - inoculated maize roots. Soil Biol. Biochem. 21: 147-153.
30. EYERS, M. , VANDERLEYDEN, J., VAN GOOL, A. 1988a. Attachment of Azospirillum to isolated plant cells. FEMS Microbiol. Lett. , 49: 435-439.
31.Fuqua, W.C.,Winans S.C.
32. Fuqua, C., Burbea, M., Winans, S.C. 1995. Activity of the Agrobacterium Ti plasmid conjugal transfer regulator TraR is inhibited by the product of the traM gene. J. Bacteriol. 177:1367-1373.
33. Gavrila, L., Mihaescu, G. 1989. Biologia microorganismelor fixatoare de azot. Editura Ceres.
34. Ghetea, L., Dumitrescu, M., Toma, N. Aspecte moleculare si biochimice ale interactiei gazda - parazit in cadrul procesului tumorigen la plante. In: Dumitru I.F. (ed.) 2002: Romanian Biotechnological Letters, pp. 84-87.
35. Gonzlez, J.E., Marketon, M.M. 2003. Quorum Sensing in Nitrogen-Fixing Rhizobia. Microbiology and Molecular Biology Reviews, 67:574-592.
36. Hardy, R.W.F., 1987, "Genetic Engineering for Nitrogen Fixation", National Academy of Sience Washinghton D.C., 77-106.
37. Hellingwerf, K.J. 2005. Bacterial observations: a rudimentary form of intelligence? Trends in Microbiology, 134:152-158.
38. Higa, T. 1991. Effective microorganisms: A biotechnology for mankind. p.8-14. In J.F. Parr, S.B. Hornick, and C.E. Whitman (ed.) Proceedings of the First International Conference on Kyusei Nature Farming. U.S. Department of Agriculture, Washington, D.C., USA.
39. Hurst C.J., Knudsen G.R., McInerney M.J., Stetzenbach L.D., Walter M.V. (1997): Manual of environmental microbiology, ASM. Press, Washington.
40. Haukka K., 1997, Teza de doctorat
41. Kavadiaa A., Vayenasb D.V., Pavlouc S., Aggelisa G.:Dynamics of free-living nitrogen-fixing bacterial populations in antagonistic conditions, Ecological modeling, (2007) 243-253.
42. KUMAR, D.P., HEGDE, M., BAGYARAJ, D.J., MADHAVA-RAO, A.R., 1998, Influence of biofertilizers on the growth of cashew (Anacardium occidentale L.) rootstocks.
43. Lyon, G.J., Wright, J.S., Christopoulos, A., Novick, R.P., Muir, T.W. 2002. Reversible and specific extracellular antagonism of receptor-histidine kinase signaling. J. Biol. Chem. 277:6247-6253.
44. MILLET, E., FELDMAN, M. 1986. Yield response of a common spring wheat cultivar to inoculation with Azospirillum brasilense at various levels of nitrogen fertilization. Plant Soil, 80: 255-259.
45. MERTENS, T., HESS, D. 1984. Yield increases in spring wheat (Triticum aestivum L.) inoculated with Azospirillum lipoferum under greenhouse and field conditions of a temperate region. Plant Soil, 82: 87-99.
46. Mclean, K.H., Winson, M.K., Fish, A., Taylor, A., Chhabra, S.R., Camera, M., Daykin, M., Swift, S., Lamb, J., Bycroft, B.W., Stewart, G.S.A.B. & Williams, P. 1997. Quorum sensing in Chromobacterium violaceum: exploitation of violacein production and inhibition for the detection of N-acylhomoserine lactones. Microbiology 143: 3703−3711.
47. Mor, M.I., Finger, L.D., Stryker, J.L., Fuqua, C., Eberhard, A., Winans, S.C. 1996. Enzymatic synthesis of a quorum-sensing autoinducer through use of defined substrates. Science, 272:1655-1658.
48. O'HARA, G. W., DAVEY, M. R., LUCAS, J. A. 1981. Effect of inoculation of Zea mays with Azospirillum brasilense strains under temperate conditions. Can. J. Microbiol. 27: 871-877.
49. O'HARA, G. W., DAVEY, M. R., LUCAS, J. A. 1987. Effect of nitrogen on the yield response of Pennisetum americanum, Triticum aestivum and Zea mays to inoculation with Azospirillum brasilense under temperate conditions. Biol. Fertil. Soils. 4: 67- 72.
50. OKON, Y. 1982. Azospirillum: physiological properties, mode of association with roots and its application for the benefit of cereal and forage grass crops. Isr. J. Bot. 31: 214-220.
51. OKON, Y., HADAR, Y. 1987. Microbial inoculants as crop-yield enhancers. CRC Crit. Rev. Biotechnol. 6: 61-85.
52. OKON, Y., HEYTLER, P.G., HARDY, R.W.F. 1983. N2 fixation by Azospirillum brasilense and its incorporation into host Seratia italica. Appl. Environ. Microbiol. 46: 694-697.
53. OKON, Y., KAPULNIK, Y. 1986. Development and function of Azospirillum -inoculated roots. Plant Soil, 90: 3-16.
54. Pappas, K.M., Weingart, C.L., Winans, S.C. 2004. Chemical communication in proteobacteria: biochemical and structural studies of signal synthases and receptors required for intercellular signalling. Molecular Microbiology, 53:755-769.
55. Parsek, M.R., Greenberg, E.P. 2005. Sociomicrobiology: the connections between quorum sensing and biofilms. Trends in Microbiology, 13:27-33.
56. Parr, J.F., S.B. Hornick, and D.D. Kaufman. 1994. Use of microbial Inoculants and organic fertilizers in agricultural production.
57. Pearson, J.P., Gray, K.M., Passador, L., Tucker, K.D., Eberhard, A., Iglewski, B.H., Greenberg, E.P. 1994 Structure of the autoinducer required for expression of Pseudomonas aeruginosa virulence genes. Proc. Natl. Acad. Sci. USA 91:197-201.
58. Pearson, J.P., Passador, L, Iglewski, B.H.,
Greenberg, E.P.
59. Redfield, R.J. 2002. Is quorum sensing a side effect of diffusion sensing? Trends in Microbiology, 10(8):365-370.
60. SARIG, S., KAPULNIK, Y., OKON, Y. 1986. Effect of Azospirillum inoculation on nitrogen fixation and growth of several winter legumes. Plant Soil, 90: 335-342.
61. SARIG, S., BLUM, A., OKON, Y. 1988. Improvement of the water status and yield of field-grown grain sorghum (Sorghum bicolor) by inoculation with Azospirillum brasilense. J. Agric. Sci. l18: 271-277.
62. Scragg. A, 2005: Environmental biotechnology, University Press, Oxford.
63. Schultze M., Kondorosi A.: Regulation of symbiotic root nodule development, Annu. Rev. Genet., 32 (1998) : 33 - 57.
64. Szab I.M. (1989): A bioszfra mikrobiolgija, Akadmiai Kiad, Budapesta.
65. Thorne, S.H., Williams, H.D. 1999. Cell density-dependent starvation survival of Rhizobium leguminosarum bv. phaseoli: identification of the role of an N-acyl homoserine lactone in adaptation to stationary-phase survival. J. Bacteriol., 181:981-990.
66. Van Heerden P.D.R., De Beer M., Mellet D.J., Maphike H.S., Foit W.: Growth media effects on shoot physiology, nodule numbers and symbiotic nitrogen fixation in soybean, South African Journal of Botany (2007) in press.
67. Waters, C.M., Bassler, B.L. 2005. Quorum sensing: cell-to-cell communication in bacteria. Annu. Rev. Cell. Dev. Biol. 21:319-346.
68. Whitehead, N.A., Barnard, A.M.L., Slater, H., Simpson, N.J.L., Salmond, G.P.C. 2001. Quorum-sensing in Gram-negative bacteria. FEMS Microbiology Reviews, 25:365-404.
69. Wilkinson, A., Danino, V., Wisniewski-Dye, F., Lithgow, J.K., Downie, J.A.. 2002. N-Acyl-homoserine lactone inhibition of rhizobial growth is mediated by two quorum-sensing genes that regulate plasmid transfer. J. Bacteriol., 184:4510-4519.
70. Winzer, K., Hardie, K.R., Wiliams, P. 2002. Bacterial cell-to-cell communication: sory, can`t talk now - gone to lunch! Curent Opinion in Microbiology, 5:216-222.
71. Wisniewski-Dye, F., and J. A. Downie. 2002. Quorum-sensing in Rhizobium. Antonie Leeuwenhoek, 81:397-407.
72. Xavier, K.B., Bassler, B.L. 2005. Regulation of
uptake and processing of the quorum-sensing autoinducer AI-
73. Zarnea G. (1984): Tratat de microbiologie generala, Editura Academiei Romane, Bucuresti.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 8569
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved