| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Criteriile de evaluare a diversitatii bacteriilor
Estimarea diversitatii vietii este o provocare permanenta a biologiei. Pentru microorganisme, scopul este complicat de faptul ca subiectul inventarierii nu este vizibil cu ochiul liber si nici nu se diferentiaza pe criterii morfologice. Estimarile anterioare ale numarului de specii bacteriene erau de 107- 109.
Clasificarea microorganismelor, anterior utilizarii analizei moleculare, s-a facut pe baza urmatoarelor criterii:
morfologice (coci, bacili, spirili)
tinctoriale (reactia Gram)
structurale
metabolice (prin evidentierea capacitatii de a metaboliza anumite glucide sau aminoacizi)
biochimice prin evidentierea markerilor moleculari caracteristici (mureina, acizii teichoici, lipidele cu legaturi eterice din membrana Archaea etc).
Criteriile clasice de clasificare a bacteriilor s-au dovedit utile pentru determinarile curente de laborator, dar nu au permis elaborarea unui sistem de clasificare bazat pe criterii naturale si nici explicarea legaturilor filogenetice dintre procariote si eucariote.
Pornind de la observatia lui Chatton (l925), in acord cu care exista doua tipuri distincte de organizare a lumii vii, procariot si eucariot, R. Stanier (in anii '60) a elaborat conceptul clasic, unificator, privind structura si functiile celulei bacteriene, pe baza unor criterii discriminatorii de tipul 'totul sau nimic'. Procariotele au fost definite pe baza diferentelor de compozitie chimica si a functiilor pe care le indeplinesc structurile omologe ale organismelor procariote si eucariote.
Conceptul elaborat de Stanier, desi a dominat gandirea microbiologica circa 30 de ani, nu corespunde realitatii stiintifice, deoarece nu cuprinde toate procariotele.
Clasificarea lumii vii in 3 domenii pe baza criteriilor moleculare
In ultimii 15-20 de ani, locul criteriilor clasice a fost luat de metodele de analiza moleculara.
Descoperirea organismelor procariote, grupate in domeniul Archaea a invalidat conceptul clasic de "bacterie", bazat pe particularitati biochimice si functionale discriminatorii, in raport cu celula eucariota. Microorganismele Archaea creeaza o punte de legatura intre tipul de organizare procariota si eucariota (Woese, l994), permitand elaborarea unui arbore filogenetic comun al organismelor procariote si eucariote.
Metodele de biologie moleculara, de secventiere a proteinelor si acizilor nucleici (denumite semantide - Zuckerkandl si Pauling, 1965) au permis conturarea filogeniei bacteriene intr-un sistem de clasificare care cuprinde toate procariotele. Cu cat secventele semantidelor a doua organisme sunt mai asemanatoare, cu atat ele sunt mai apropiate filogenetic. Acest criteriu presupune ca genele codificatoare sa nu se transfere pe orizontala, de la o celula la alta.
La bacterii, secventierea proteinelor pentru scopuri filogenetice este neadecvata, pentru ca uneori, o proteina are o distributie limitata sau este greu de secventiat (Zinder, l998).
Metodele de biologie moleculara au fost orientate in primul rand asupra studiului acizilor nucleici. S-a determinat procentul G + C din ADN, dar testul nu este edificator, deoarece doua organisme cu secvente diferite de ADN pot avea aceeasi proportie de G + C. Testul ramane valabil pentru a caracteriza un organism nou.
Tehnica secventierii ADN a revolutionat sistemele de clasificare a bacteriilor, deoarece a permis accesul la informatia continuta in ADN.
Un alt test furnizat de biologia moleculara, util pentru studiul filogeniei bacteriene este hibridarea ADN-ADN. ADN se denatureaza prin tratament termic sau cu baze. Metoda evidentiaza capacitatea unei secvente de ADN monocatenar al unui organism de a forma un heteroduplex cu ADN monocatenar de la alt organism, pe baza omologiei secventei de baze. Daca deosebirile secventei de baze sunt mai mari de l5%, heteroduplexul nu se formeaza, astfel ca acest test este negativ pentru organismele indepartate filogenetic, dar este decisiv pentru delimitarea speciei bacteriene.
Woese si Fox au propus, ca instrument filogenetic pentru evaluarea raportului evolutiv dintre microorganisme, analiza secventei ADN din care este transcris ARNr 16S (ribotipia).
Ribotipia presupune folosirea probelor capabile sa detecteze genele ce codifica ARNr. Utilizarea genelor ARNr pentru identificarea bacteriilor isi are originea in gradul inalt de conservare a genelor codificatoare a ARNr. Genele pentru sinteza ARNr sunt organizate in operoni, in care genele individuale sunt adeseori separate prin ADN necodificator. O proba de ARNr marcat sau ADN de la o specie va hibrida cu variate regiuni ale ADN de la specii bacteriene neinrudite.
Tehnica de ribotipie implica izolarea ADN total al celulei bacteriene si fragmentarea sub actiunea unei enzime de restrictie. Rezulta astfel o colectie de fragmente de ADN cu o distributie uniforma a dimensiunilor. Fragmentele se separa prin electroforeza in gel de agaroza si se transfera pe o membrana de nitroceluloza prin capilaritate (blotting). Apoi fragmentele sunt hibridate cu o proba marcata care contine genele ARNr (ARNr de E. coli). Hibridarea apare numai in acele fragmente cromosomale care contin secventele genelor ARNr.
Ribotipia a fost una dintre primele tehnici moleculare folosita cu succes pentru taxonomia vibrionilor. Astfel s-au identificat (si s-au depozitat in GenBank) peste 78 000 secvente ale genei pentru sinteza ARNr 16S, izolate de la bacterii cultivate sau obtinute prin amplificare, direct de la probele din sol, fara cultivare.
In anii '70, C. Woese a initiat studiul secventei ARNr (16S), ca un posibil marker filogenetic, folosind ARNr l6S cu lungimea l500-l600 nucleotide, component al subunitatii mici (30 S) a ribosomului. Tehnica presupune izolarea ARNr si digestia sa cu RN-aza T1, care cliveaza dupa fiecare rest de G. Rezulta o varietate de fragmente de oligoribonucleotide, cu lungimea de l-25 baze, fiecare terminandu-se cu G. Amestecul se separa prin metoda electroforezei bidimensionale. Se inregistreaza tabloul oligonucleotidelor si se compara cu cel obtinut de la alte organisme, stabilindu-se un coeficient de asemanare, a carui valoare variaza intre 0 (pentru neasemanare) si 1 pentru identitatea completa. Astfel s-a evidentiat ca Cyanobacteria sunt asemanatoare cu cloroplastele. Secventele obtinute prin amplificarea directa a probei din mediu, ofera singura informatie disponibila pentru 99% dintre procariotele din mediile naturale.
Studiile recente sugereaza ca numarul total al speciilor bacteriene, evaluat prin secventa de baze a genelor pentru ARNr 16S, este de peste 1030, grupate in cel putin 50 de filumuri bacteriene. Jumatate dintre ele sunt alcatuite in intregime din bacterii necultivabile. Alte 3 filumuri sunt formate din specii cultivate in proportie mai mica de 10%. Membrii a numai 6 filumuri au fost cultivati in proportie de peste 90%.
Rezultatele cercetarilor de nivel molecular au aratat ca sistemul de clasificare a lumii vii in cele 5 regnuri nu este corect din punct de vedere filogenetic, din mai multe motive:
cele doua regnuri de microorganisme eucariote - Protista si Fungi sunt artificiale;
plantele si animalele (Metaphyta si Metazoa) au evoluat din organisme eucariote unicelulare;
in sistemul celor 5 regnuri, diferentele dintre Monera (Procaryotae) si celelalte 4 regnuri sunt mult mai mari decat diferentele intre reprezentantii celor 4.
Noul sistem de clasificare accepta ca forma de organizare eucariota insumeaza o multitudine de caractere comune si defineste o unitate filogenetica.
Regnul Procaryotae (Monera) reuneste diviziunea EUBACTERIA si ARCHAEA. La nivel citologic, ambele sunt procariote, dar la nivel molecular ARCHAEA se disting de EUBACTERIA. Ele nu constituie o unitate filogenetica, deoarece nu intrunesc caractere unitare. La nivel molecular, organismele ARCHAEA se aseamana mai mult cu EUCARIOTELE decat cu EUBACTERIILE. Din aceasta cauza, regnul Procaryotae (Monera) este un taxon nevalidat de datele de ordin molecular.
Woese (l990) considera ca noul sistem filogenetic, pe baza criteriilor moleculare, permite impartirea lumii vii in trei sisteme primare distincte. Clasificarea trebuie sa recunoasca, in primul rand, cele trei tipuri fundamentale de organisme: Eubacteria, Archaea si Eucaryotae. Gruparile trebuie sa aiba statutul de domenii, o categorie taxonomica superioara regnului. Autorul propune abandonarea denumirii de Archaebacteria, deoarece sugereaza inrudirea stransa cu Eubacteria, care in realitate nu exista. Cele trei domenii se numesc Bacteria (sinonim Eubacteria), Archaea si Eucarya (sinonim Eucaryotae), care include organismele eucariote (protozoare, alge, fungi, plante, animale).
Din punct de vedere fiziologic, microorganismele domeniului Archaea apartin la trei tipuri: metanogene, halofile si sulf-dependente.
Metanogenele sunt strict anaerobe si produc gaz metan ca produs final al fermentatiei.
Cele sulf-dependente traiesc in medii termofile si reduc sulful elementar sau oxideaza compusii sulfului pentru a obtine energie sub forma potentialului reducator. Sunt specii aerobe si anaerobe.
Halofilele traiesc in medii hipersaline si unele se protejeaza de lumina cu un pigment care contine carotenoizi. Sunt aerobe obligate.
Din punct de vedere filogenetic, domeniul Archaea are doua diviziuni distincte (Kandler, l993):
Crenarchaeota (crenos = izvor) cuprinde exclusiv microorganisme hipertermofile dependente de sulf, care isi dobandesc energia prin reactii de reducere a sulfului (Thermoproteus, Pyrodictium etc.), sau de oxidare a compusilor cu sulf (Sulfolobus). Sunt raspandite in mediul marin si in mediile vulcanice terestre;
Euryarchaeota cuprinde metanogenele chimiolitotrofe hipertermofile strict anaerobe ce cresc la ll0o (Methanopyrus) si metanogenele mezofile (din apa, sol, intestinul animalelor). Alti reprezentanti sunt halofilele extreme ce traiesc in solutii saturate de NaCl (Halobacterium halobium), sulfat-reducatorii termofili (Thermococcus, Pyrococcus).
Clasificarea lumii vii in trei domenii recunoaste independenta filogenetica a bacteriilor (Eubacteria) si a reprezentantilor Archaea.
Nici unul dintre sistemele de clasificare a lumii vii nu include virusurile. Ele formeaza un grup de entitati infectioase fara echivalent in lumea vie, dar au relatii foarte stranse cu aceasta, deoarece se multiplica numai in substratul celular viu.
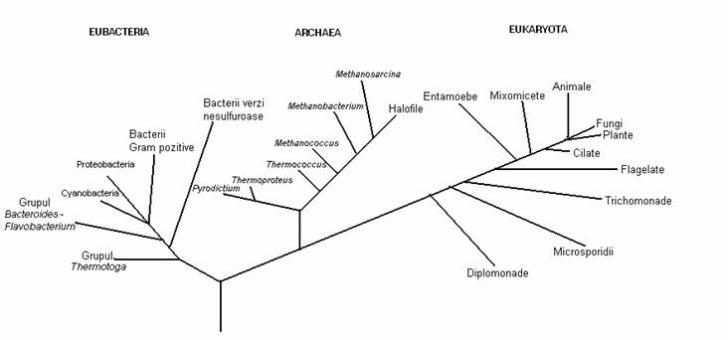
Fig. 1: Arborele filogenetic universal, bazat pe secventa ARNr a subunitatii ribosomale mici, care cuprinde cele trei domenii (dupa Woese, 1991)
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 1763
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved