| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Cromozomul bacterian
1 Dimensiune
Imensa majoritate a bacteriilor detin ca material genetic esential o molecula ADN dublu catenar circular covalent inchis, suprarasucit negativ si impachetat. Desi complexarea cu proteine si impachetarea nu este identica cu cea din cromozomii de tip eucariot, totusi aceasta molecula este denumita tot "cromozom" sau nucleoid. Cromozomul bacterian este atasat la membrana plasmatica a celulei bacteriene in aproximativ 20 de puncte, dar o functie deosebita o are punctul de atasare de langa regiunea de origine a replicarii acestei molecule de ADN (ori C).
Cei mai mici cromozomi de tip procariot masoara mai putin de 1 Mpb si se intalnesc mai ales la bacteriile fara perete celular (bacteriile din genurile Mycoplasma, Ureaplasma au cromosomi cu dimensiuni cuprinse intre 600 si 800 kpb). La cealalta extrema se afla bacteriile din genurile Myxococcus si Calothrix, care au cromozomi foarte mari (aproximativ 12-13000 kpb). Escherichia coli are un cromozom de dimensiune intermediara: 4700 kpb.
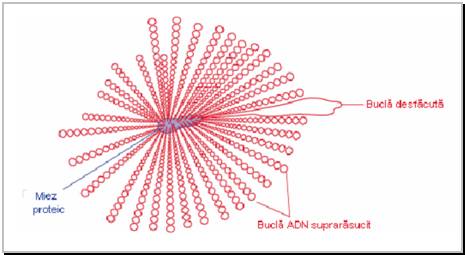
Figura 3.1 Modelul Pettijohn de structura a nucleoidului din bacteria Escherichia coli
Intre 40 si 50 de bucle suprarasucite radiaza dintr-un miez proteic (dupa Brown, 2002).
2 Modelul Pettijohn
In general, molecula ADN ce formeaza cromozomul bacterian este de aproximativ 1000 de ori mai lunga decat celula bacteriana. Aceasta molecula este complexata cu proteine, suprarasucita si impachetata formand o structura conforma cu modelul elaborat de Pettijohn in 1974 (Figura 3.1). Conform acestui model, prin asocierea ADN cu proteine si cu ARN (de regula, ARN nascent) se formeaza aproximativ 50 de domenii topologice, semi-independente (denumite si bucle), per genom de E.coli. Fiecare domeniu (bucla) este suprarasucit separat de celelalte domenii si poate fi relaxat independent de celelalte prin introducerea unei rupturi monocatenare. Gradul de suprarasucire negativa este dat de balanta dintre activitatea a doua enzime - ADN giraza si ADN topoizomeraza I - amandoua regland densitatea helicala a moleculelor de ADN. ADN giraza creste numarul de spire per kilopereche de baze azotate (suprarasuceste molecula de ADN), iar ADN topoizomeraza I scade numarul de spire.
Principalul tip de structura secundara a ADN ce formeaza cromozomul bacterian este forma B de dreapta, dar aceasta configuratie nu se gaseste uniform de-a lungul intregului cromosom bacterian. Alte configuratii, prezente pe distante scurte, sunt ADN-Z (in dublu helix de stanga), structuri cruciforme (in regiunile cu secvente invers repetate) si regiuni triplu-catenare (in zone extrem de bogate in purine sau pirimidine).
Este de reamintit faptul ca, in general, expresia genica este afectata nu numai de secventa primara a ADN (succesiunea de nucleotide), ci si de structura secundara si tertiara a acestuia.
In majoritatea cazurilor, compozitia globala in nucleotide a unei molecule de ADN se exprima prin procentul molar de guanina + citozina (%mol GC), restul pana la 100% fiind, evident, reprezentat de adenina + timina (datorita criteriului de complementaritate intre cele doua catene ADN, respectiv intre bazele purinice ii cele pirimidinice). Compozitia in nucleotide a cromozomului bacterian variaza in limite foarte largi: 25 %mol GC la Mycoplasma capricolum si 75 %mol GC la Micrococcus luteus.

Procentul molar de guanina - citozina in cromozomul bacterian
Procentul molar de guanina + citozina este corelat si cu compozitia in codoni a genomului. Aceasta variaza in sensul preferintei pentru 1-2 codoni dintr-un grup de codoni sinonimi. Se constata astfel ca la Mycoplasma capricolum este favorizata prezenta A/T in pozitia 3-a a codonilor sinonimi.
S-a mai constatat ca in cromosomul bacterian exista si o serie de gene a caror activitate poate afecta compozitia totala in nucleotide a unei molecule de ADN. Astfel, la E.coli au fost descrise doua gene (mut T si mut Y) care afecteaza frecventa transversiilor A-T / C-G si, respectiv, C-G / A-T. Echilibrul intre exprimarea acestor doua gene afecteaza compozitia globala in nucleotide a moleculei de ADN ce reprezinta cromosomul de E.coli.
O alta problema o reprezinta contextul de citire a unui anumit codon, in speta, configuratia codonilor adiacenti. Numarul teoretic posibil de codoni (sens) adiacenti este foarte mare (612 = 3721), dar, examinand 237 de gene de la E.coli s-a constatat ca perechile de codoni adiacenti nu sunt distribuite randomizat, ci anumite perechi de codoni sunt mai abundente decat altele. S-a dedus astfel ca, compozitia in nucleotide a unui codon este corelata si cu compozitia codonilor adiacenti, corelat probabil cu procesele de atasare la situsurile A (Aminoacil) si P (Peptidil) ale ribozomilor. Se pare deci, ca aparatul de traducere a informatiei genetice ar fi putut sa determine evolutia unor anumite trasaturi ale matritei genetice.
|
Figura 3.2 Organizarea genomului la cateva specii de procariote.
|
5 Proteine asociate cu cromozomul bacterian
In afara de ARN polimeraza, care datorita ratei inalte de transcriere la procariote, ramane asociata cvasi-permanent cu moleculele de ADN, cromosomul bacterian este asociat cu o serie de proteine bazice, similare cu histonele de la organismele eucariote, denumite proteine histone-like.
La E.coli au fost descrise 9 specii moleculare de proteine bazice, cu greutate moleculare intre 9 si 28 kd, care se ataseaza la ADN intr-o maniera situs-nespecifica si care au functii similare cu histonele de la organismele eucariote. Dintre cele 9 specii moleculare proteice, cea mai importanta este o proteina denumita HU ( Helix-Unwinding = desfacerea dubului helix). Din complexarea moleculelor HU cu ADN cromozomal bacterian se formeaza structuri similare cu nucleosomii de la eucariote, denumite structuri nucleosom-like (chiar si in conditii in vitro), in care sunt cuprinse circa 200 pb/nucleosom. Structurile nucleosom-like nu sunt statice, ci se afla intr-un echilibru dinamic.
Studii mai ample au demonstrat faptul ca proteina HU nu este o simpla proteina structurala a arhitecturii cromozmului bacterian, ci are rol foarte important in diverse procese din celula bacteriana - in mod special in procese ce implica interactiuni ADN-proteine, situatie in care moleculele HU favorizeaza atasarea altor proteine la ADN.
Desi proteinele histone-like au fost studiate extensiv la E.coli, totusi au fost identificate proteine cu functii omoloage si la alte genuri si specii bacteriene. Pe de alta parte, s-a constatat ca o serie de microorganisme din grupul Archaea prezinta proteine histone-like cu structura intermediara intre cele de la Bacteria si histonele de la Eukarya.
 Proteina HU
Proteina HU
este cea mai importanta proteina histone-like de la E.coli
au fost descrise proteine tip HU la toate bacteriile studiate
ca si histonele de la organismele eucariote, proteina HU se ataseaza la molecule de ADN d.c. indiferent de secventa de nucleotide, deci situs - nespecific
are un caracter bazic si o greutate moleculara de 9,7 kd
functioneaza ca dimer
un monomer este format din 2 polipeptide, denumite HU- si HU- si codificate de gene diferite: hup A si, respectiv, hup B.
are proprietati fizice si compozitie totala in aminoacizi similara cu histonele de la eucariote, dar secventa de aminoacizi este diferita de acestea.
ca si histonele de la eucariote, proteina HU are un grad inalt de conservare in lumea bacteriana
exista aproximativ 25000 de molecule HU per genom de E.coli, dar aceste molecule nu sunt dispuse uniform de-a lungul cromozomului bacterian, ci sunt mai dense la periferia nucleoidului (in zonele extrem de active transcriptional)
Proteina IHF
 Alte proteine histone-like de la E.coli sunt proteina H (similara cu histona H2A de la eucariote),
proteina H1, proteina H-NS, proteina IHF.
Alte proteine histone-like de la E.coli sunt proteina H (similara cu histona H2A de la eucariote),
proteina H1, proteina H-NS, proteina IHF.
Proteina IHF (Integration Host Factor, g.m.=20 kd)
a fost descoperita in studiile privind integrarea genomului fagului in cromozomul de E. coli ; ulterior, s-a constatat ca aceasta proteina intervine si intr-o serie de alte procese din celula bacteriana
este formata din 2 subunitati codificate de genele him A si him B.
spre deosebire de alte proteine histone-like, moleculele de IHF se ataseaza la ADN intr-o maniera situs-specifica (adica doar la anumite secvente de nucleotide)
moleculele de IHF nu sunt necesare pentru recombinarea bacteriana omoloaga, dar par sa intervina in fenomene de recombinare neomoloaga, specializata, cum sunt de altfel, procesele de transpozitie si de conjugare bacteriana
6 Secvente ADN repetate
In cromosomul bacterian au fost identificate secvente ADN repetate, dintre care unele nu codifica proteine/ARN (secvente repetate necodificatoare), iar altele codifica o serie de proteine sau specii moleculare de ARN (ARNr, ARNt) - secvente repetate codificatoare.
6.1 Secvente repetate necodificatoare = ADN repetitiv
Acestea sunt reprezentate de secvente scurte ce apar de regula in afara ORF-urilor (Open Reading Frames). Marea majoritate a acestor secvente servesc ca situsuri de interactiune ADN-proteine (inclusiv in procese de recombinare, inversie, excizie, transpozitie) si ca situsuri modificate ce identifica catena matrita in timpul replicarii semiconservative a cromozomului bacterian. Dintre cele mai cunoscute asemenea secvente sunt secventele REP, Chi, Dam.
Secvente REP (Repeated Extragenic Palindromes)
Sunt formate din palindroame repetate, majoritatea aflate in regiuni extragenice. O secventa REP contine o secventa consensus de 38 bp. Grupuri (denumite "clusteri") de 2-4 secvente REP separate intre ele prin 20 pb formeaza un element REP (REPE = REP Element). La E.coli si Salmonella typhimurium exista 100-200 structuri REPE (reprezentand deci, aproximativ 0.5% din cromozomul bacterian), localizate insa diferit. La secventele REP se leaga molecule de proteina HU si de ADN giraza, aceste secvente avand deci rol in impachetarea nucleoidului. Tot la aceste secvente se leaga si ADN polimeraza intervenind in procese de replicare si reparare ADN.
Situsuri Chi Un situs Chi are o lungime de 8 pb:
5' GCTGGTGG 3'
Datorita faptului ca reprezinta situsul de recunoastere si taiere a moleculelor ADN de catre endonucleaza RecBCD, secventele Chi stimuleaza recombinarea omoloaga mediata de RecA si RecBCD. S-a mai constatat ca efectul recombinativ al situsului Chi este polar: stimuleaza recombinarea la capatul 5' al lui Chi, si nu la 3'. Situsuri Chi au fost identificate atat pe cromosom, cat si pe unele plasmide de la E.coli. Mai mult chiar, s-a constatat ca secventele Chi reprezinta situsuri de stimulare a recombinarii genetice la toti membrii familiei Enterobacteriaceae. In contrast, la Pseudomonadaceae, complexul enzimatic RecBCD nu actioneaza la situsuri Chi.
In cromosomul de E.coli au fost identificate aproximativ 950 de situsuri Chi, ce par sa fie distribuite relativ uniform (1 situs Chi la 5 kbp), cu exceptia zonei adiacente regiunii oriC, unde exista un cluster de 22 de situsuri Chi.
Situsuri Dam sunt formate din 4 bp (5' GATC 3') si reprezinta situsurile de metilare a adeninei (adenina este metilata la pozitia N6). Cromosomul de E.coli contine foarte multe situsuri Dam (peste 18.000) care, daca ar fi dispuse randomizat ar exista 1 situs Dam/250 bp. Si in acest caz s-a constatat un numar foarte mare de situsuri Dam in, si in jurul, regiunii oriC: in cele 245 bp ce definesc oriC la E.coli exista 8 situsuri Dam, iar in cele 350 bp ce flancheaza oriC exista inca 12 situsuri Dam.
6.2 Secvente repetate codificatoare
Asemena secvente au dimensiuni mult mai mari decat cele necodificatoare.
operoni rrn exista la toate bacteriile si cuprind secvente ce codifica pentru molecule de ARN ribozomal (ARNr). La cele mai multe dintre speciile bacteriene, linkage-ul (ordinea) in cadrul unui operon rrn este 16S - 23S - 5S. E.coli si S.typhimurium au cate 7 loci (operoni) rrn, localizati in pozitii echivalente. Cei 7 operoni sunt notati rrnA..rrnG, fiecare cuprinzand secvente 16S-23S-5S cotranscrise intr-o molecula ARN de 30S. Numarul operonilor rrn variaza de la un gen/specie bacteriana la alta: Bacillus subtilis are 10 operoni rrn, Mycobacterium sp. are 1-2 operoni rrn. Chiar in cadrul aceleiasi tulpini bacteriene, locii rrn pot diferi intre ei in ceea ce priveste prezenta/absenta genelor pentru ARNt in regiunile "spacer" dintre genele ARNr. Astfel, toti cei 7 operoni rrn de la E.coli au 1-2 gene pentru ARNt intre secventele pentru 16 si 23S, si 0-2 gene ARNt dupa 5S.
Distributia locilor rrn pe cromosom variaza la diverse specii bacteriene: la E.coli, majoritateta locilor rrn sunt localizati in jumatatea cromosomala ce are in centru regiunea oriC; la B.subtilis, locii rrn sunt grupati intr-o zona ce reprezinta 30% din cromosom.
genele pentru arnt
Cromosomul de E.coli contine 41 de gene/operoni pentru ARNt distribuiti in tot cromosomul. Unii din acesti operoni codifica pentru o molecula de ARNt, altii pentru mai multe molecule de ARNt, iar altii au si secvente ce codifica diverse proteine.
secvente rhs ("rearrangement hot-spots") sunt capabile sa genereze duplicatii genice prin procese de crossing-over inegal intre secvente repetate. La E.coli K-12 au fost identificate 4 secvente rhs, notate rhs A, B, C si D. Fiecare din aceste 4 secvente au dimensiuni intre 8 si 9 kbp si sunt compuse dintr-o regiune "core" (miez) (cu secventa conservata de aproximativ 3700 bp) si din segmente flancatoare. Prin exprimarea regiunilor "core" ale secventelor rhs A, B si C sunt sintetizate 2 proteine cu functie deocamdata necunoscuta. In contrast cu E.coli K-12, la E.coli B si E.coli C, precum si la S.typhimurium, nu au fost descrise secvente rhs.
7 Numarul de copii genice
La bacterii, in afara de genele sau operonii aflati in copii multiple, numarul de copii ale unei gene cromosomale poate varia in functie de urmatorii factori:
1. pozitia genei pe cromosom, de-a lungul unui gradient pornind de la oriC spre regiunea ter. In timpul replicarii cromosomului bacterian, in functie de pozitia lor pe cromosom, unele gene sunt deja replicate, in timp ce altele inca nu. Astfel, unele gene se gasesc intr-un numar mai mare de copii decat altele.
2. copii ale unei gene cromosomale pot exista si pe plasmide.
In aceasta situatie se foloseste termenul de meroploidie partiala, care se refera la o ploidie partiala ce afecteaza anumite gene, fara sa existe multiplii de cromosomi intregi (este cazul genelor prezente si pe cromosom si pe un plasmid).
3. intr-o celula bacteriana pot exista mai multe copii ale intregului cromosom.
La anumite specii bacteriene exista mai multe copii cromosomale in aceeasi celula: Desulfovibrio vulgaris (4 copii), D.gigas (17 copii), Azotobacter chroococcum (20-25 de copii), A.vinelandii (40-80 de copii cromosomale).
Este de subliniat faptul ca la majoritatea speciilor bacteriene cu copii cromosomale multiple exista procese de inactivare cromosomala ce asigura functionarea unui singur cromosom si inactivarea celorlalti.
Densitatea informatiei biochimice, adica numarul de reactii biochimice catalizate per kb de material genetic, este un parametru extrem de variabil in genomul bacterian, in primul rand datorita faptului ca nu intotdeauna genele au o relatie 1-la-1 cu reactiile biochimice. In mod uzual, este valabila relatia 1 gena => 1 polipeptida => 1 reactie biochimica. Exista insa si foarte multe exceptii, de exemplu:
- enzime formate din mai multe lanturi polipeptidice: de exemplu, enzima succinat dehidrogenaza este formata din 4 polipeptide codificate de 4 gene diferite - sdh A, B, C si D. In acest caz, relatia este 4 gene =>1 reactie biochimica;
- enzime polifunctionale, ce catalizeaza mai multe reattii biochimice: de exemplu, complexul enzimatic FAD ce oxideaza acizii grasi este format din 2 polipeptide codificate de 2 gene diferite - fadA si fadB. Acest complex enzimatic catalizeaza 5 reactii biochimice (din care 4 sunt catalizate de FadA). In acest caz, relatia este 1 gena => 4 reactii biochimice.
 Gene suprapuse
Gene suprapuse
In cadrul genomului bacterian variaza si densitatea de citire a informatiei genetice prin transcriere. Astfel, desi marea majoritate a genelor bacteriene sunt contigue, totusi cromosomul bacterian cuprinde si gene cu diverse grade de suprapunere:
- gene cu promotor plasat in regiunea terminala a genei "upstream": trpA-trpB, ilvA-ilvD
- gene cu promotor plasat in interiorul genei "upstream": mioA-mioD
- gene cu grad ridicat de suprapunere:
1. gene suprapuse total, codificate pe cele 2 catene ale ADN; de exemplu, locusul CysE (codifica serin-acetil-transferaza), ce este implicat in biosinteza cisteinei, are 2 ORF (cys X si cys E) codificate pe cele 2 catene ale locusului CysE.
2. secvente codificatoare ce sunt folosite de mai multe ori, pornind transcrierea din pozitii diferite; de exemplu, locusul McrB (codifica proteine implicate in restrictia ADN la 5-metil-citozina) contine 3 ORF-uri pe aceeasi catena ADN, pornind din 3 situsuri diferite de initiere a transcrierii. Traducerea celor 3 transcripte duce la formarea a 3 proteine (de 51, 53 si, respectiv, 54 kdal) cu secventa carboxi-terminala identica, dar diferita in regiunea amino-terminala.
3. gene suprapuse total, cu molecule transcript identice, dar traduse diferit prin procese de "frameshift" translational: de exemplu, gena trpR care codifica 2 proteine si gena dnaX, care codifica 2 subunitati ale ADN polimerazei III.
Asemenea gene - cu grad ridicat de suprapunere - sunt foarte bogate in informatie genetica (unele au chiar informatie dubla). Multe din ele (de exemplu, genele mcrB si dnaX) nu sunt tipice pentru cromosomul de E.coli, in ceea ce priveste procentul molar de guanina + citozina. Astfel, gena mcrB are 48% mol GC, iar gena dnaX are 58 % mol GC, in timp ce cromosomul de E.coli are, in toto, 51-51.5 % mol GC. S-a emis ipoteza ca asemenea gene care prezinta un procent molar de guanina + citozina extrem de diferit fata de media cromosomala, ar fi fost achizitionate de E.coli din alte genomuri, prin procese de transfer de material genetic pe orizontala (transformare, conjugare, transductie).
9 Redundanta genelor si a produselor genetice
Pentru foarte multe functii celulare, exista cate 2 gene, ca si cum programul genetic al acestor bacterii ar avea sisteme de "backup" (copii de siguranta). Aceasta redundanta ar putea fi considerata ca fiind o densitate redusa a informatiei genetice. Cu toate acestea, cel putin in unele cazuri, aceasta informatie este reglata diferit si folosita in scopuri metabolice diferite.
In cromosomul de E.coli exista 58 de perechi (de cate 2 gene) sau clusteri (mai mult de 2 gene), ce codifica 125 de produse genice. Din acestea, in 56 de perechi/clusteri reactiile catalizate sunt identice. In cazul anumitor grupuri genice, reglajul exprimarii lor este diferit, de exemplu:
- genele aroF, aroG si aroH codifica, toate trei, o aceeasi enzima: 3-deoxi-D-arabino-heptulosonat-7 - fosfat-sintetaza (DAHPaza). Enzima codificata de aroF este sensibila la concentratia de tirozina, cea codificata de aroG este sensibila la concentratia de fenil-alanina, iar cea codificata de aroH este sensibila la concentratia de triptofan ;
- genele glpA si glpD codifica amandoua glicerol-3-fosfat-dehidrogenaza. Enzima codificata de glpA este sintetizata si folosita in conditii de anaerobioza, iar cea codificata de glpD - in conditii de aerobioza.
Unele din aceste gene pereche au secventa similara una cu cealalta (si proteinele corespunzatoare au secventa similara in aminoacizi) si este probabil ca au aparut prin procese de duplicatie genica, urmata de o oarecare divergenta functionala.
Altele insa, desi enzimele desfasoara aceeasi functie, totusi au secventa diferita (atat ca proteine, cat si genele corespunzatoare). Este posibil ca asemenea gene sa fi aparut fie prin evolutie convergenta, fie prin transfer lateral de la alti repliconi bacterieni (aceasta ultima situatie poate fi identificata prin determinarea procentului molar de guanina + citozina).
Multi bacteriofagi isi insera genomul in cromosomul bacterian, fie prin transpozitie in pozitii randomizate in cromosom (bacteriofagul Mu), fie prin recombinare situs-specifica numai in anumite pozitii (fagii lambdoizi ce constituie fagii inruditi cu fagul ). Situsurile cromosomale in care fagul lambda (si fagii lambdoizi) isi insera genomul, sunt denumite situsuri attB ("attachment on bacteria") si corespund unor situsuri attP ("attachment on phage") pe genomul fagic. Fagul lambda necesita un situs attB de minimum 21 bp, din care 15 sunt identice cu 15 bp din attP (234 bp). Lambda are un situs preferat attB in cromosomul de E.coli K-12 (dar si situsuri secundare) ce e situat intergenic. Alti fagi lambdoizi (21, e14, P22) se insereaza in situsuri attB cu localizare intragenica. Pe cromosomul de E.coli K-12 au fost cartate situsurile pentru 15 fagi lambdoizi si s-a constatat ca ele sunt grupate intre pozitiile 6 min si 44 min.
Aceasta organizare a situsurilor in care se inserera genomul fagilor lambdoizi reprezina inca un argument in favoarea ipotezei conform careia cromosomul de E.coli (si nu numai) ar avea o origine himerica, unul din segmente derivand dintr-o gazda ancestrala a fagilor lambdoizi.
Pe de alta parte, s-a constatat ca foarte multe elemente genetice cu caracter "mobil" se insera in genele pentru ARNt, fapt pentru care s-a sugerat ca acestea ar fi reprezentat situsurile folosite de fagii ancestrali.
In cromosomul de E.coli, majoritatea genelor (operonilor) par sa fie distribuiti randomizat (indiferent de inrudirea lor fiziologica), cu exceptia celor implicati in catabolismul glucozei ce par sa fie dispusi in 4 zone echidistante pe harta circulara a cromosomului. Aceasta randomizare a dispunerii operonilor a fost verificata si prin rearanjamente cromosomale induse experimental - caz in care operonii au continuat sa functioneze normal.
Totusi, in treimea cromosomala ce are in centru regiunea ter (ca, de altfel, si in regiunea oriC), pozitia operonilor pare sa fie destul de fixa, orice rearanjament deranjand total functionarea acestora. Orice aranjament nerandomizat al genelor duce la ipoteza ca pozitia genelor ar putea afecta functionarea lor, deducandu-se astfel o organizare mai inalta decat operonul sau reglonul.
La cele mai bine studiate specii de Pseudomonas (P.aeruginosa, P.putida) s-a constatat ca genele cu functii "housekeeping" (gene implicate in metabolismul central al oricarei celule, precum si in metabolismul anabolic) sunt grupate in jumatate din harta circulara a cromosomului. In acelasi timp, genele implicate in metabolismul catabolic par sa fie dispuse randomizat in cealalta jumatate a cromosomului bacterian.
Acest mod de dispunere a genelor ar putea reflecta istoria constructiei cromosomului din parti separate: genele pentru metabolismul esential ar fi existat probabil in acel genom ancestral mai mic, care a fost ulterior marit la actuala dimensiune prin fuziune cu un alt element genetic sau prin transfer de material genetic (transformare, conjugare, transductie).
In general, s-a constatat ca la foarte multe bacterii clusterii genici din jurul regiunilor oriC si ter sunt foarte similari intre taxoni bacterienei foarte indepartati filogenetic, fapt ce sustine ipoteza ca initierea si terminarea replicarii cromosomului bacterian sunt procese extrem de vechi si sunt coordonate prin mecanisme similare la marea majoritate a bacteriilor.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 18527
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved