| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Metabolismul energetic bacterian
Metabolismul celulei bacteriene, ca si al celulei eucariote, are doua compartimente fundamentale:
caile de producere a energiei, denumite catabolice sau degradative;
caile consumatoare de energie, denumite anabolice sau de biosinteza.
Deoarece caile catabolice si anabolice sunt strans interconectate prin intermediul unor compusi cheie ai metabolismului intermediar, adevarate "placi turnante" ale metabolismului, denumirea comuna de "cai amfibolice" este justificata.
La bacterii, datorita ratei foarte inalte a metabolismului, functioneaza o categorie speciala de cai, denumite anaplerotice (sau de alimentare), menite sa aprovizioneze o cale metabolica ce functioneaza intens, cu produsul intermediar necesar.
Caile catabolice reprezinta ansamblul proceselor biochimice prin care are loc degradarea nutrientilor preluati din mediu si eliberarea energiei. Complexitatea reactiilor catabolice este variabila, in functie de natura si complexitatea substantelor supuse proceselor de degradare.
In general, reactiile catabolice degradative ale unei macromolecule se desfasoara in trei faze succesive:
in prima faza se desfasoara reactiile biochimice catalizate de exoenzime, in cursul carora macromoleculele nutritive sunt scindate in spatiul extracelular, la subunitatile specifice: glucide, aminoacizi, oligopeptide, nucleozide, fosfati organici cu molecula mica (glicerol-fosfat). Bacteriile, fungii si protozoarele produc enzime extracelulare (exoenzime), pe care le elimina in mediu sau raman legate de structurile de suprafata ale celulei. Exoenzimele sunt hidrolaze: polizaharidaze (amilaza, celulaza, pectinaza), mucopolizaharidaze (hialuronidaza, chitinaza, neuraminidaza, lizozim), nucleaze, lipaze, fosfolipaze, proteaze. Ele hidrolizeaza moleculele mari, rezultand oligomeri sau monomeri, pentru care structurile celulare de invelis sunt permeabile. Pentru microorganismele patogene, exoenzimele reprezinta un factor major de virulenta, deoarece ataca tesuturile constitutive ale gazdei. La bacteriile Gram negative, sediul exoenzimelor este spatiul periplasmic, deoarece traiesc in medii oligotrofe. In aceasta etapa se elibereaza numai 1% din energia macromoleculei, care se pierde sub forma de caldura
in cea de a II-a etapa, monomerii sunt degradati pe cai specifice si rezulta un numar limitat (circa l2) de molecule mici, denumite intermediari metabolici ai cailor centrale, aceiasi la diferite tipuri de bacterii. In aceasta etapa se elibereaza circa 1/3 din energia moleculara;
cea de a III-a faza este diferita in functie de natura reactiilor metabolice: in aerobioza, reactiile evolueaza in sensul metabolizarii integrale a compusilor, pana la CO2 si H2O, cu eliberarea intregii cantitati de energie. In anaerobioza, reactiile evolueaza dupa modelul respiratiei anaerobe sau al proceselor fermentative si cantitatea de energie care se elibereaza este mica.
Reactiile catabolice sunt exergonice: ele realizeaza tranzitia unei molecule de la o stare mai putin stabila, cu un continut mai mare de energie, la molecule mai mici si mai stabile, cu un continut mai mic de energie.
Tipuri de metabolism energetic in functie de acceptorul final de electroni
In functie de natura acceptorului final de e-, microorganismele pot prezenta trei tipuri de metabolism energetic:
metabolism energetic de tip aerob (oxibiotic) sau metabolism respirator, dependent de citocromii catenei de respiratie celulara, propriu microorganismelor al caror acceptor final de e-, la capatul catenei de respiratie este O2;
metabolism energetic anaerob (anoxibiotic), caracteristic microorganismelor al caror acceptor final de e- este un compus anorganic oxigenat (nitratul, sulfatul, carbonatul) sau un compus organic (fumaratul). In prezenta O2, respiratia este de tip aerob, deoarece catena de respiratie celulara este functionala;
reactii metabolice de tip fermentativ sunt acelea care au loc numai in conditii de anaerobioza, dar acceptorul final de e- este un compus organic (de exemplu, piruvatul).
La bacteriile fermentative lipseste lantul citocromilor din catena de respiratie, pentru transportul e-. Substratul acceptor este redus la produse de fermentatie (de exemplu, piruvatul, la lactat). Reducerea acceptorului final este cuplata cu reoxidarea coenzimelor.
Pentru nevoile curente ale cultivarii microorganismelor in conditii de laborator sunt recunoscute urmatoarele tipuri respiratorii:
bacterii strict aerobe sunt acelea care necesita O2 molecular ca acceptor final de e-. Ele realizeaza metabolism de tip oxidativ si nu cresc in absenta O2. Pentru multe bacterii aerobe este necesara aerarea extensiva, pentru ca O2 este putin solubil in apa si nu difuzeaza cu o rata care sa egaleze rata utilizarii. Aerarea se face prin agitare viguroasa a recipientului sau prin barbotarea aerului sterilizat in mediul lichid, printr-un tub de sticla. Bacteriile aerobe cresc mult mai bine in conditii de aerare fortata decat daca O2 difuzeaza liber;
bacteriile microaerofile folosesc O2 ca acceptor final de e-, dar concentratia naturala a O2, de 20-21% este prea mare si din aceasta cauza nu cresc pe suprafata mediului expusa aerului atmosferic, dar nu cresc nici in anaerobioza;
bacteriile facultativ anaerobe isi obtin energia pe cale oxidativa, folosind O2 ca acceptor final de e-, dar cresc si in conditii de anaerobioza si isi obtin energia pe cale fermentativa;
bacteriile anaerobe nu folosesc O2 molecular ca acceptor de e-. In interiorul acestui grup se disting cateva diviziuni fiziologice:
a) bacteriile anaerobe aerotolerante cresc putin (dar cresc) in conditii de aerobioza, intr-o ambianta care contine intre 2-8% O2. Cresterea optima are loc in anaerobioza (de exemplu, bacteriile lactice);
b) bacteriile anaerobe obligate variaza in privinta sensibilitatii lor la O2: unele tolereaza cantitati mici de O2, iar altele strict anaerobe nu cresc in prezenta unei concentratii mai mari de 0,5% O2.
Efectul toxic asupra bacteriilor
anaerobe il exercita O2 si nu potentialul redox
ridicat al mediului. In prezenta O2, bacteriile anaerobe
isi pierd viabilitatea deoarece nu pot sa detoxifice intermediarii
toxici ai reducerii O2: H2O2, O2-
(radicalul superoxid),
O2 se formeaza dupa ce molecula de O2 absoarbe o cantitate de energie. In molecule, in general, e- apar in perechi stabilizate, cu spini de directie opusa. O2 este o molecula neobisnuita, prin faptul ca spinii electronilor au aceiasi directie. Absorbtia energiei schimba unul dintre e- pe un orbital cu energie mai inalta si se produce inversia spinului.
Multe bacterii anaerobe obligate sunt bogate in flavine (enzime), care pot reactiona spontan cu O2 si formeaza produse toxice. Din aceasta cauza, cultivarea lor se face in recipiente din care O2 se elimina complet, pentru a crea un potential redox scazut.
Exista numeroase procedee pentru diminuarea continutului de O2:
unele simple (umplerea recipientelor cu mediu si acoperirea cu dop de cauciuc), pentru bacteriile care tolereaza o concentratie mica de O2 in mediu;
altele mai complexe pentru cultivarea bacteriilor strict anaerobe: in mediul de crestere se adauga agenti reducatori (acidul tioglicolic, cisteina) sau alaturi de mediu, in atmosfera inchisa a recipientului de cultivare (pirogalolul).
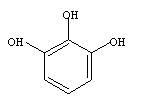
Pirogalolul
Agentii reducatori reactioneaza cu O2 dizolvat in mediu sau existent in atmosfera recipientului si-l reduc la H2O.
Reactii de oxidare a substratului energetic
Metabolismul energetic este o inlantuire de reactii de oxido-reducere, in cursul carora substratul energetic este oxidat. Reactiile de oxidare sunt de trei tipuri:
l. Reactia de oxidare consecutiva pierderii de e-. De exemplu, Fe2+ (feros), forma redusa este oxidata la Fe3+ (feric), dupa reactia:
![]()
2. Reactii de oxidare catalizate de dehidrogenaze, in care molecula oxidata pierde simultan protoni si e-, de exemplu, oxidarea alcoolilor la aldehide:
![]()
3. Reactia de oxidare care comporta castig de atomi de oxigen de catre substratul oxidat; de exemplu, oxidarea aldehidelor la acizi:
![]()
sau reactia de decarboxilare oxidativa a piruvatului, la acetat si CO2:
![]()
Aceste reactii de oxidare constau intr-o dehidrogenare, in care apa are, in acelasi timp, rolul de donor de O si de sursa de protoni si e-.
Toate reactiile de oxidare biologica au comun faptul ca, in esenta, ele semnifica o pierdere de e- de catre substrat si eliberarea energiei.
Electronul cedat de un substrat molecular trebuie obligatoriu sa fie acceptat de un alt substrat, astfel incat reactiile de oxidare biologica sunt cuplate totdeauna cu reactii de reducere, formand un cuplu de oxido-reducere (un sistem redox), in care o molecula se comporta ca donoare de e-, iar cealalta, ca acceptoare.
Molecula care cedeaza e- sau protonii (H+) se oxideaza, iar cea care ii accepta se reduce.
Potentialul redox si forta proton-motrice
Din punct de vedre chimic, oxidarea semnifica pierderea unuia sau mai mulor e- dintr-o molecula, iar reducerea semnifica castigul e-. In biochimie, oxidarea si reducerea nu implica totdeauna transferul e-, ci si al atomilor de H. {n reactiile de oxido-reducere biologica, dehidrogenarile sunt foarte importante si de aceea, termenii de donor de H+ si de acceptor de H+ sunt folositi ca sinonimi cu cei de donor si respectiv de acceptor de e-.
Tendinta unui compus chimic de a ceda sau de a accepta e- in reactiile de oxido-reducere biologica se exprima cantitativ prin potentialul oxido-reducator (redox) al cuplului. Electronii sunt transportati totdeauna de la un cuplu cu un potential redox mai negativ, la un cuplu cu un potential redox mai putin negativ (de exemplu, de la cuplul NADPH+ +H+, la cuplul Fe2+/Fe3+).
In natura, mediile bogate in O2 au un potential redox de + 0,82 V, iar cele bogate in H, au un potential de - 0,42 V.
Ca regula generala, substraturile cu potential redox pozitiv (O2, Fe3+) sunt oxidante (acceptoare de e-), iar cele cu potential redox negativ (H2, NADH) sunt agenti reducatori (donori de e-).
O substanta chimica, cu cat este mai redusa, cu atat contine o cantitate mai mare de energie si cu atat este mai mare tendinta ei de a ceda e-.
Transferul protonilor (H+) si al e- in sistemele biologice, se face de la substratul energetic la O2. Daca acest transfer s-ar face direct de la substratul redus la O2, diferenta maxima de potential ar fi de 0,82 - (- 0,42) = l,23 V, ceea ce ar corespunde unei diferente energetice de 57 kcal/mol. Un astfel de transfer ar fi insotit de o pierdere foarte mare de energie.
Microorganismele (ca si celelalte organisme) dispun de mai multe sisteme redox, care
transporta e- in cascada, permitand eliberarea treptata a energiei.
Transportorii de e- pot fi impartiti in doua clase:
liber difuzibili (NAD+, NADP+). Totdeauna transfera 2 atomi de H spre urmatorul purtator al catenei. Transferul atomului de H se numeste dehidrogenare;
ferm atasati de enzimele din membrana citoplasmatica. In dublul strat fosfolipidic sunt inserate sistemele redox - proteine cu rol de transport in catena de respiratie celulara: flavoproteine, quinone, proteine cu Fe-S, citocromii.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 1536
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved