| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Nucleul
Nucleul a fost descoperit in 1831 de catre R. BROWN in celulele vegetale. A putut fi usor observat in microscopia optica datorita refringentei sale, superioare citoplasmei.
Totodata, nucleul este rezistent fata de fixatori, are afinitati tinctoriale ridicate, iar comportamentul sau particular in timpul diviziunii a atras atentia citologilor, astfel ca acest organit celular a fost unul din cele mai bine studiate.
Nucleul indeplineste un rol in nutritie, morfogeneza (diferentiere), depoziteaza informatia genetica si asigura transmiterea acesteia in generatiile celulare succesive.
Majoritatea celulelor poseda un singur nucleu, adica sunt uninucleate. Alte celule ale eucariotelor pot fi lipsite de nucleu, anucleate, caz foarte rar, ca de ex. celulele tuburilor ciuruite din tesutul liberian de la plantele vasculare. La inceput insa si acestea sunt nucleate dar isi pierd nucleul in timpul diferentierii. La ciupercile superioare, ascomicete si bazidiomicete exista stadii in dezvoltarea lor in care celulele contin doi nuclei, fiind binucleate, in cazul miceliului dicariotic de scurta durata (Ascomicetae) sau de lunga durata (Bazidiomicete). Algele verzi sifonale, ciupercile inferioare (Phycomycetae) ca si mixomicetele prezinta o structura cenocitica (sincitiala sau plasmodiala) adica o masa citoplasmatica cu mai multi nuclei, neseparati de perete celular.
Pozitia nucleului in celula este una centrala in celulele tinere, meristematice, cu citoplasma abundenta dar lipsite de vacuole. In timpul diferentierii celulare se formeaza vacuole mari care preseaza citoplasma si nucleul la periferie, dar nucleul ramane intotdeauna cuprins in citoplasma (Fig. 66).
Dimensiunile nucleului sunt reduse la ciuperci (1-2 μm in diametru), la vegetalele superioare prezinta o dimensiune medie de 10-50 μm si pot deveni foarte mari la unele celule reproducatoare ca de ex. oosferele (300-500 μm la Zamia, Dioon edule).
A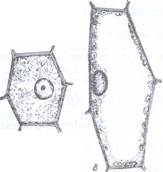
Fig. 66: Pozitia nucleului in celula in timpul diferentierii celulare.
Raportul dintre volumul nucleului si al citoplasmeieste unul constant pentru celulele apartinand aceluias tesut si specie si este cunoscut sub numele de raport nucleoplasmatic al lui
V nucleului
![]() HERTWIG: RNP =
HERTWIG: RNP =
Vcitoplasmei
In celulele meristematice acest raport este de 0,5 apoi se micsoreaza progresiv, atingand valori minime la celulele mature (0,1).
Forma nucleului este una sferica in celulele meristematice dar devine lenticular in celulele parenchimatice adulte sau chiar fuziform in cele tubuloase. Mai rar poate capata forme lobate, ramificate sau filamentoase. In timpul diviziunii, nucleul sufera modificari importante, cromatina sa da nastere la cromozomi iar membrana nucleara si nucleolii se dezorganizeaza.
Nucleul care nu este in diviziune se numeste nucleul intefazic, in repaus sau metabolic. Desi nici una din aceste denumiri nu este corecta trebuie precizat ca aceste denumiri nu semnifica altceva decat ca nu este in diviziune. Notiunea de nucleu interfazic este potrivita numai pentru celulele meristematice, cea de metabolic nu este potrivita deoarece si nucleul in diviziune are metabolism, iar prima notiune de nucleu in repaus nu este potrivita sistemelor biologice care exclud starea de repaus.
1. Nucleul in repaus este format din 4 componente: membrana nucleara, cromatina, nucleoli si nucleoplasma (Fig. 67).
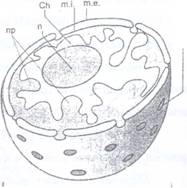
Fig. 67: Schema structurii si componentei unui nucleu interfazic. me- membrana externa; mi- membrana interna; Ch-cromatina; Pn- pori nucleari; n- nucleol; np- nucleoplasma.
Structura nucleului se poate studia in microscopia optica, dupa fixare si colorare cu coloranti azici care se fixeaza usor pe ADN (fuxina bazica) cromatina nucleara fiind Feulgen pozitiva. Nucleolul poate fi colorat si cu colorantii acizi dar este Feulgen negativ datorita ARN.
1. Membrana nucleara este o membrana dubla, una externa si una interna, separate de un spatiu perinuclear. Fiecare membrana are grosimea de 75 A si o structura tripartita. Cele doua membrane se unesc la nivelul porilor membranei. Porii au un diametru de 300-1000 A si sunt inconjurati de un material inelar de natura proteica. Prezenta porilor s-a evidentiat in ME prin tehnica criodecapajului. In unele puncte, membrana externa nucleara este in contact cu reticulul endoplasmatic iar spatiul perinuclear comunica cu reticulul endoplasmatic. La fel ca unele regiuni ale RE, si membrana externa nucleara poate retine ribozomi (Fig. 68, 69).
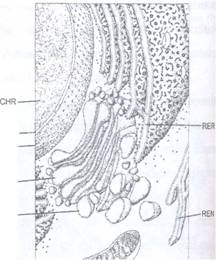
Fig. 68: Diagrama reprezentand nucleul (stg. sus) cu membrana dubla (MN), pori (P) si cromatina (CHR), reticul endoplasmatic rugos cu ribozomi atasati (RER) si neted (REN), un dictiozom cu saculi si vezicule (D, VG) si o mitocondrie (M).
Desi existenta porilor membranei nucleare a fost confirmata, transportul prin membrana nucleara prezinta o mare selectivitate iar in transportul macromolecular intervine si materialul inelar din jurul porilor, ex. transportul ARN din subunitatile ribozomale.
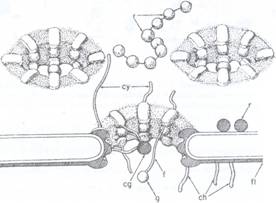
Fig. 69: Diagrama reprezentand structura porilor membranei nucleare. r- ribozomi pe fata citoplasmatica a membranei nucleare; ch- legaturi cu cromatina nucleara; g-granule; f- fibrile; cg- granula centrala. Fiecare por are 8 subunitati dispuse in 3 zone: externa, centrala si interna.
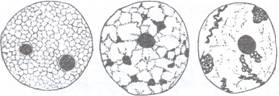
2. Cromatina nucleara. In nucleul interfazic cromatina se gaseste in retea, datorita impletirii filamentelor de cromatina care poate fi mai condensata si se numeste heterocromatina sau mai laxa, dispersata si se numeste eucromatina considerata partea activa a cromatinei. In cele doua zone ale cromatinei raportul histone/ ADN este diferit, fiind mai mare pentru zonele heterocromatice. Dupa densitatea retelei de cromatina, nucleul poate avea o structura:
eureticulata, cu o retea foarte densa de cromatina (ex. Allium cepa)
reticulata cu o retea mai putin densa (ex. Vicia faba, Pisum sativum, Zea mays)
semireticulata cu o retea foarte laxa de cromatina (ex. Solanum, Bryonia)
areticulata fara o retea de cromatina (Fig. 70).
Fiecare din cele patru tipuri descrise pot prezenta sau nu regiuni heterocromatice, cu cromatine foarte condensate care se numesc cromocentre. Daca aceste cromocentre sunt in numar egal cu cromozomii speciei, aceste regiuni se numesc procromozomi, fiecare dintre acestea apartinand unui cromozom diferit.
Structura cromozomiala persista si in nucleul interfazic, ei fiind doar mascati prin intindere, deformare, impletire. Existenta procromozomilor ca si prezenta cromozomilor politenici sunt argumente in favoarea permanentei cromozomilor si la nivelul nucleului interfazic.

a b c d
Fig. 70: Structura nucleului in repaus dupa densitatea retelei de cromatina si prezenta sau absenta cromocentrelor. a- structura eureticulata cu doi nucleoli, fara cromocentre; b- structura reticulata cu cromocentre; c- structura semireticulata cu cromocentre; d- structura areticulata cu procromozomi.
Structura fibrei de cromatina. In nucleul eucariotelor, ADN-ul este asociat cu doua categorii de proteine: histonice si nehistonice. Din aceasta asociere sau impachetare rezulta fibra de cromatina. Histonele sunt proteine cu masa moleculara mica si contin aminoacizi bazici si cu functii alcoolice (lizina, serina) incarcati pozitiv, ceea ce le pemlite legarea lor de ADN la nivelul radicalilor fosfat, incarcati negativ.
Din interactiunea ADN cu proteinele histonice, care au o forma sferica sau de cilindru turtit rezulta o structura numita nucleozom (Fig. 71). In jurul unei proteine histonice se infasoara doua lanturi de ADN formate din 146 perechi de nucleotide.
Nucleu
nucleosomic Histone
nucleosomice
110 A 57 A
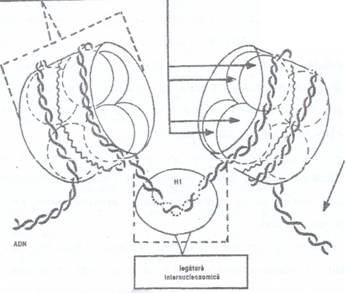
Fig. 71: Schema structurii moleculare a nucleosomilor
Regiune
AND in dublu helix (5 Ture) in interfaza Portiune
a cromatinei formata din AND si proteine histonice Cromatina
de 30 nm in profaza cu nucleosomii impachetati strans Fibra
de cromatina in cromozomul profazic de 300 nm Fibra
de cromatina din cromozomul metafazic de 700 nm Cromozom
metafazic
Aceasta fibra
primara se impacheteaza si se spiralizeaza formand o
fibra de cromatina de 30 nm diametru, caracteristica pentru
nucleul interfazic. In timpul diviziunii, aceasta se contracta si mai
mult printr-o supraspiralizare formand o cromatina de 300 nm pentru
cromozomi profazici si de 700 nm pentru o cromatida a unui cromozom
metafazic (1400 nm) (Fig. 72,73).
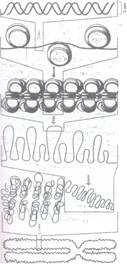
Fig. 72: Transformarile fibrei de cromatina in interfaza si in timpul diviziunii.
Proteinele nehistonice sunt proteine cu masa moleculara ridicata si indeplinesc o functie de reglare genetica intre care cea de initiere a replicarii ADN, controlul transcriptiei genelor, s.a.
3. Nucleolul caracterizeaza nucleul in repaus al tuturor celulelor eucariote.
Numarul nucleolilor din nucleu este proportional cu numarul cromozomilor astfel ca determinarea lor reprezinta un indiciu comod de stabilire a gradului de poliploidie. Nucleolul este lipsit de membrana proprie fiind delimitat doar de cromatina asociata perinucleolar, alte regiuni ale cromatinei patrund in interiorul nucleului si se numeste cromatina asociata intranucleolar (Fig. 74).

Fig. 73: Ciclul de spiralizare a cromonemei in timpul diviziunii. 1- interfaza; 2, 3, 4 profaza (se observa cele doua cromatide dispuse in spirala minora); 5- prometafaza (cele doua cromatide incep sa se separe una de alta); 6- metafaza in care se observa o spirala majora care da un aspect condensat; 7- anafaza in care se separa cele doua cromatide in cromozomi fii; 8- telofaza, in care se produce despiralizarea; C-centromer.
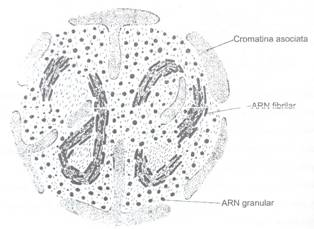
Fig. 74: Schema organizarii ultrastructurale a nucleolului
In interiorul nucleolului s-au evidentiat prin ME, urmatoarele 4 componente:
fibrile grupate cu structura dubla
granule dispersate
matrice amorfa de natura proteica
cromatina asociata
Pentru ca fibrilele si granulele sunt distruse de ribonucleaza se considera ca acestea sunt formulate din ARN. Nucleolul este locul de sinteza a ARN ribozomal care se face in contact cu cromatina asociata. In nucleol mai au loc si sinteze proteice dar nu se poate preciza daca proteinele din constitutia ribozomilor se sintetizeaza in nucleol sau in citoplasma, Asamblarea ribozomilor are loc totusi in nucleol de unde sunt apoi exportati in citoplasma, unde are loc maturarea lor.
4. Nucleoplasma sau sucul nuclear contine ARN, proteine, glicoproteine si enzime. Ea reprezinta sediul unor importante reactii metabolice, sinteza nucleotidelor si a proteinelor nucleare.
Compozitia chimica a nucleului. Cel mai important component este ADN localizat in cromatina nucleara. Continutul sau este caracteristic si constant pentru o specie, dar variaza mult cu specia, fara a exista o relatie intre continutul de ADN si gradul de evolutie. Continutul de ADN din nucleu se exprima in pg (picograme) (1 pg = 10-12 g).
Cateva exemple pentru nucleii unor celule diploide:
Allium cepa 40 pg
Vicia faba 18 pg
Pisum sativum 10 pg
Zea mays 5 pg
Homo sapiens 5,6 pg
Al doilea component al nucleului este ARN localizat in nucleoli si in sucul nuclear. Din cantitatea totala de ARN celular, ARN-ul nuclear reprezinta doar 5-10%, restul se gaseste in citoplasma la nivelul ribozomilor.
Proteinele reprezinta cea mai importanta (cantitativ) componenta chimica a masei uscate a nucleului. Proteinele au insa mai multe fractiuni intre care proteinele solubile din nucleoplasma, ribonucleoproteine, histone bazice, proteine nehistonice, unele legate altele nelegate de ADN.
Intre enzime se remarca cele ale sintezelor acizilor nucleici, ARN si ADN-polimeraza, NAD-sintetaza, nucleotidtrifosfataza, histonacetilaza. Majoritatea sunt localizate in nucleoplasma. Exista apoi enzime ale glicolizei anaerobe si ale metabolismului nucleotidic. Lipidele se gasesc in componenta membranelor nucleare si sunt reprezentate de lecitina si colesterol.
2. Nucleul in diviziune
In timpul diviziunii celulare, nucleul sufera modificari importante, membrana si nucleolii se dezorganizeaza iar cromatina devine intens colorabila si va da nastere la cromozomi.
Vom prezenta in continuare numai aspectele legate de cromozomi, procesul de diviziune va fi studiat in capitolul de fiziologie celulara.
Morfologia cromozomilor. Cromozomii celulelor vegetale prezinta o forma si marime variabila. Ei pot fi in forma de granule sferice, bastonase scurte sau alungite, in forma literei U sau V. Pe fiecare cromozom se observa o zona care nu fixeaza colorantii cromatinei si care se numeste centromer si corespunde unei zone de strangulare a cromozomului din care cauza se mai numeste si constrictie primara. Cromozomii in forma de U sau V au centromerul in zona curburii lor.
Centromerul imparte cromozomul in doua brate, superior si inferior centromerului fiecare fiind clivat longitudinal in doua cromatide (Fig. 75 A, B, C).
Dupa pozitia centromerului cromozomii pot fi metacentrici, cu centromerul situat central, submetacentrici cu centromerul situat in apropierea zonei mediane, telocentrici, cu centromerul situat spre varful cromozomului si acrocentric cu centromerul situat in varf (Fig. 76). Unii cromozomi pot prezenta si o a doua constrictie denumita secundara, la o extremitate a cromozomului, separand o mica portiune de cromozom numita satelit (sau trabant) (Fig. 75 B).
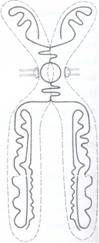
Fig. 75 A: Interpretarea structurii unui cromozom metafazic.
Cromozomii in forma de bastonas, cei mai frecventi, au o lungime de 2-20 μm si grosimea de 1-4μm. Cei mai mari cromozomi se intalnesc la unele specii de Monocotyledonate (Trillium, Tradescantia),
Toate speciile se caracterizeaza printr-un numar definit de cromozomi, constant si par in celulele vegetative sau somatice. Acesta este numarul diploid sau 2n. Sporii, gametofilul si gametii prezinta un numar de cromozomi redus la jumatate adica un numar haploid sau n de cromozomi. In aceste celule este prezent doar unul din cei doi cromozomi ai fiecarei perechi si se repartizeaza astfel in urma diviziunii reductionale.
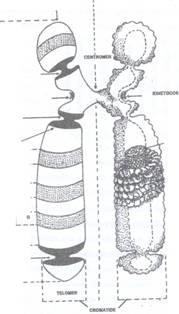
Fig. 75 B: Organizarea cromozomului metafazic la eucariote.
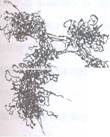
Fig. 75 C: Aspect EM al unui cromozom.

Fig. 76. Tipuri de cromozomi dupa pozitia centromerului
A - metacentric, B - submetacentric, C - telocentric, D - acrocentric
Numarul de cromozomi este variabil pentru diferite specii, neexistand o relatie intre numarul de cromozomi si gradul de evolutie. Cateva exemple (2 n) sunt prezentate mai jos:
|
Crepis capillaris | |
|
Triticum monococcum |
|
|
T. durum | |
|
T. aestivum (T. vulgare) | |
|
Allium cepa | |
|
Zea mays | |
|
Lilium candidum | |
|
Datura stramonium | |
|
Nicotiana tabacum | |
|
Prunus laurocerasus | |
|
Ferigi |
Unele specii pot avea un numar multiplicat de cromozomi (4n, 6n, 8n) adica intregul genom si se numesc poliploide. Poliploidia poate sa apara spontan sau poate sa fie indusa pe cale chimica (cu colchicina) sau fizica (cu radiatii). Acest procedeu se utilizeaza in ameliorarea unor soiuri de plante cultivate, poliploizii fiind valorosi din punct de vedere economic si au vitalitate sporita.
Multiplicarea doar a unor cromozomi se numeste aneuploidie si apare frecvent cu ocazia diviziunii reductionale, in timpul formarii gametilor astfel ca unul din gameti poate avea unul (n+l) sau doi (n+2), cromozomi in plus si altii in minus (n-l, n-2). Dupa fecundare cu astfel de gameti, se vor forma nuclei cu numar de cromozomi 2n+ 1 sau 2n-1.
Fenomenul de politenie apare in urma multiplicarii, fara diviziune, a filamentelor de ADN in interiorul nucleului conducand la aparitia cromozomilor uriasi.
Ansamblul cromozomilor metafazici, ordonati dupa marimea lor descrescanda si aliniati la nivelul centromerului, cu bratul scurt la partea superioara se numeste cariotip. In celulele diploide exista cate o pereche pentru fiecare cromozom. Acestia sunt cromozomii omologi sau autozomi. La speciile vegetale dioice (canepa, urzica, hamei, salcie, plop) unul din sexe se caracterizeaza printr-un cromozom diferit sau absent. Acestia se numesc cromozomi sexuali sau heterozomi. Fiecare pereche de cromozomi se noteaza cu cifre sau litere.
Reprezentarea grafica, schematizata, a cariotipului se numeste cariograma sau idiograma si se foloseste in studiile de genetica si pentru caracterizarea unei specii (Fig. 77).
Compozitia chimica a cromozomilor este foarte apropiata de a cromatinei nucleare din care provin. Continutul de ADN este proportional cu talia cromozomilor. Dintre proteine cele histonice sunt proportionale cu cantitatea de ADN, ele intrand in structura nucleozomilor, care persista si in cromozomi, fiind puternic condensati.
Alte proteine acide, nehistonice intra in
constitutia matricei cromozomiale si pot fi legate strans sau lax cu
In conceptia clasica, structura cromozomului este formata din matrice in care se gaseste cromonema ce poate avea cromomere, adica zone mai concentrate ale cromonemei. Cromonema reprezinta helixul dublu al ADN, capabil de intindere sau contractare iar cromomerele ar reprezenta genele active, informationale. Dupa teoria clasica, in timpul diviziunii se produce clivarea longitudinala a cromozomului in cele doua cromatide si totodata diviziunea longitudinala a cromonemei. In timpul telofazei dispare matricea iar cromonema se despiralizeaza, se intinde, ramanand contractate doar zonele heterocromatice care persista si in interfaza si care formeaza cromocentrele. In interfaza cromonemele despiralizate se impletesc cu alte cromoneme si formeaza reteaua de cromatina.
Fig. 77; Idiograma la Triticum durum (211 = 28).
Descoperirea, dupa 1953, a ciclului
celular si impartirii interfazei in cele 3 stadii (G1,
S si G2) a condus la interpretarea modului de duplicare a ADN
dupa modelul semiconservativ care are loc in perioada S (de
sinteza) astfel ca un cromozom profazic are o cantitate dubla de
ADN (4C) care se va repartiza in cele doua cromatide in
metafaza, fiecare din ele avand o cantitate de cromatina si ADN
(2C) corespunzatoare unei celule diploide, deci clivarea
longitudinala a cromozomului conduce la repartizarea uniforma,
egala a cantitatii de ADN in cele 2 cromatide care, din
anafaza devin cromozomi fii. Asa se poate explica pastrarea
In ME nu a putut fi observata nici matricea, nici cromonema nici cromomerele. S-au pus in evidenta insa structuri fibrilare care formeaza o retea foarte densa. Cromozomii intregi, nesectionati, apar formati dintr-o impletire neregulata de fibre de cromatine de 200-500 A in diametru. Aceste fibre pot trece dmtr-o cromatida in alta, numai la nivelul centromerului. Aceasta presupune ca in fiecare cromatida se gaseste o fibra foarte lunga, repliata de foarte multe ori si transversal si longitudinal in aceeasi cromatida (Fig. 76 B).
Alte descoperiri au adus precizari asupra structurii si rolului centromerului si kinetocorului care erau, anterior, considerate ca apartinand unei singure structuri. Centromerul si kinetocorul reprezinta doua componente diferite ale cromozomului dar care sunt strans legate intre ele. Centromerul reprezinta zona dintre cele doua cromatide pe care le leaga, iar kinetocorii sunt structuri localizate in dreptul centromerului la care se vor atasa microtubulii kinetocorici din fusul de diviziune, deci structuri care asigura cromozomului pe fusul de diviziune, centromerul nefiind implicat in acest proces. La nivelul kinetocorului se exercita forte care asigura deplasarea cromozomilor spre cei doi poli. Totusi centromerul asigura atasarea kinetocorului la cromozom in timpul mitozei.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 5845
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved