| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Relatiile fag-bacterie
Studiul relatiilor fag-bacterie a adus o contributie majora la fundamentarea si evolutia biologiei moleculare. Istoria cercetarilor interrelatiilor fag-bacterie se confunda, pana in anii '70, cu insasi istoria biologiei moleculare. Studiul interactiunii fag-bacterie a fundamentat concepte si a permis demonstrarea unor mecanisme ale biologiei moleculare:
inductia sintezei enzimelor si proteinelor codificate de fagi;
colinearitatea gena-proteina
natura nesuprapusa a codului genetic
mecanismul semiconservativ al replicarii ADN
existenta fenomenelor de restrictie si modificare la bacterii
mecanismul traducerii informatiei genetice
mecanismele mutagenezei si actiunea fenomenelor reparatorii
reglarea activitatii genelor
mecanismele oncogenezei virale
De cele mai multe ori, infectia fagica a unei culturi bacteriene sensibile, evolueaza in sensul sintezei constituientilor fagici. Se asambleaza si se elibereaza fagi progeni, rezultatul fiind liza celulei bacteriene.
Studiul multiplicarii fagilor a inceput odata cu evidentierea formarii plajelor de liza, intr-o panza de celule bacteriene, crescute pe suprafata unui mediu agarizat. Fiecare plaja de liza este initiata prin multiplicarea unei singure particule virale fagice si este rezultatul repetarii ciclului litic al infectiei fagice, in perioada de crestere a culturii bacteriene.
A II-a modalitate de evolutie a interactiunii fag-bacterie este caracteristica fagilor temperati si corespunde fenomenului de lizogenie. In interactiunea de tip lizogen, genomul viral se integreaza in structura cromosomului bacterian si se comporta ca gene cromosomale.
Ciclul litic al interactiunii fag-bacterie. Multiplicarea bacteriofagului
Multiplicarea fagului are loc intr-o serie de etape, identificate initial pentru cuplul fag T4- E. coli. Intelegerea lor a avut un rol esential pentru studiile privind interactiunea dintre virusuri si celulele animale.
Multiplicarea propriu-zisa este precedata de stabilirea contactului fizic dintre fag si celula sensibila.
Adsorbtia si fixarea fagului sunt rezultatul ciocnirilor intamplatoare dintre particula fagica si celula bacteriana, a caror frecventa depinde de densitatea lor relativa. La densitatea de l08 bacterii/ml si l07-l09 fagi/ml, in cateva minute, 90% dintre fagi se adsorb pe suprafata bacteriilor.
Pentru fixarea fagilor din seria T-par, un rol esential par sa-l aiba crosetele placii bazale, deoarece distanta dintre placa bazala si peretele celular nu este niciodata mai mica de 5 nm. Fixarea particulei fagice pe celula bacteriana, depinde de existenta, la suprafata celulei, a unor structuri chimice complementare denumite receptori de fag.
Receptorii s-au evidentiat pe membrana externa a peretelui celular si pe stratul capsular la bacteriile Gram negative, pe acizii teichoici ai bacteriilor Gram pozitive, pe flageli si pe pili.
S-au descris 4 mecanisme principale de legare a fagilor, pe suprafata celulei bacteriene:
fagii cu coada contractila din seria T, se fixeaza reversibil, prin intermediul fibrelor cozii, pe catenele glucidice ale LPS la bacteriile Gram negative, sau de acizii teichoici parietali (legati covalent de mureina), la cele Gram pozitive. Fibrele cozii fagului T4, prin capatul lor distal, interactioneaza cu specificitate inalta, cu resturile glicozil ale LPS din membrana externa la E. coli. Fixarea ireversbila este rezultatul legarii crosetelor placii bazale, cu receptorii celulei;
fagii cu coada necontractila, se leaga de componenta glucidica a LPS din membrana externa a peretelui celular. Imediat dupa aceea, are loc degradarea partiala a receptorilor sub actiunea unei endoglicozidaze virale.
De obicei, capsula bacteriana blocheaza fixarea fagilor pe structurile subiacente, dar uneori, fagii se leaga de receptorii capsulari si infecteaza celula, dupa degradarea partiala a polizaharidelor;
fagii icozaedrici ARN "masculi" se leaga pe suprafata pilului. Sinteza proteinei componente a pilului (pilina) si asamblarea ei sunt codificate de plasmida F. Pilul este o structura caracteristica numai celulelor bacteriene cu potentialitate de donor de material genetic si de aici deriva denumirea de fagi "masculi";
fagii filamentosi se fixeaza la extremitatea pilului, prin intermediul proteinei A (Attachment, engl. = atasare) a capsidei virale. Nu se cunoaste mecanismul prin care genomul fagilor ARN "masculi" sau filamentosi ajunge in celula bacteriana. Este putin probabil ca pilul sa aiba rol de "conductor" al genomului fagic. Se pare ca pilul se retracta si astfel fagii sunt orientati pana la un receptor al suprafetei celulare.
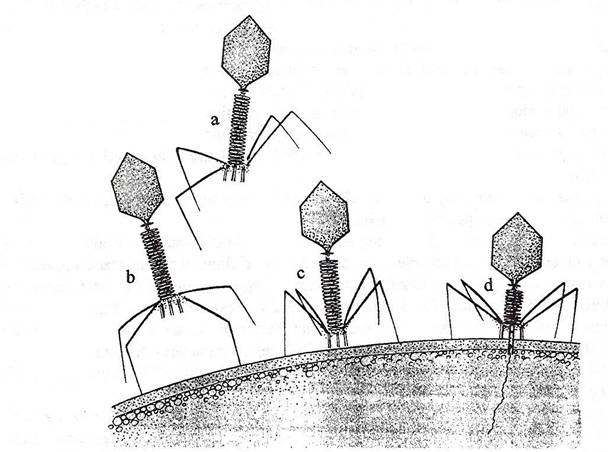
Fig. 97. Atasarea bacteriofagului T4 de peretele celular al E. coli si injectarea ADN viral. (a) Virion netasat. (b) Atasarea virionului prin fibrele cozii. (c) Fixrea ireversibila prin intermediul crosetelor placii bazale. (d) Contractia tecii cozii si injectarea ADN
(dupa Brock, 1988).
Infectia propriu-zisa. Dupa fixarea pe suprafata celulei bacteriene, placa bazala isi schimba conformatia si induce o modificare a modului de aranjare a capsomerelor tecii cozii. Ca urmare, teaca se contracta si se scurteaza, usurand patrunderea cilindrului axial al cozii, prin peretele celular, cu o lungime de circa l2 nm. ADN din capul fagului, trece in celula pe calea cilindrului axial tubular (fig. 97).
La fagii din seria T-par, coada contractila, "injecteaza" genomul in citoplasma celulei.
Capsida fagica ramane in intregime la exterior, indeplinind numai rolul unei microseringi, adaptata sa injecteze genomul fagic in celula.
Mecanismul injectarii genomului fagic nu este cunoscut cu certitudine. Membrana interna si cea externa, par sa fuzioneze local, formand un canal, sub actiunea presiunii pe care o exercita varful cilindrului axial al cozii. Acidul nucleic fagic patrunde direct, prin punctul de fuziune a membranelor. Genomul fagic ar fi orientat spre celula bacteriana, de molecule proteice "pilot" asociate genomului. Ulterior, canalul membranar se inchide cu proteinele "pilot", atasate la extremitatea proximala a ADN, cu rol in specificitatea si eficienta translocatiei ADN prin membranele celulei.
Injectarea genomului fagic se realizeaza intr-un interval foarte scurt (circa l5 secunde). In procesul transferului genomului fagic, celula bacteriana ar fi pasiva, pentru ca in capsida, ADN fagic este pliat si "impachetat" foarte strans, ceea ce ar crea o presiune datorita careia ADN, in momentul contractiei cozii si al scoaterii "dopului" proteic, ar fi propulsat energic in celula bacteriana. La aceasta s-ar adauga presiunea produsa de agitatia termica a moleculelor mari, existente in capul fagului. Alti autori considera ca celula ar exercita un efect de aspiratie, care ar facilita patrunderea genomului fagic.
Concomitent cu genomul, trec moleculele legate ionic de ADN: mici cantitati de proteine, oligopeptide bazice, poliamine.
Genomul fagic liber in celula, corespunde starii de fag vegetativ si poate fi transcris si replicat.
Infectia fagica a celulei bacteriene, determina o reorganizare profunda a activitatii sale biologice. Rezultatul interactiunii, de cele mai multe ori, este subordonarea intregului aparat de biosinteza celulara, in scopul producerii constituientilor virali. Programul genetic este transcris in doua etape: timpurie si tardiva, separate in raport de intervalul de timp al replicarii genomului.
Programul timpuriu al genomului fagic codifica urmatoarele categorii de proteine:
proteine de membrana, destinate sa "astupe" discontinuitatile produse de cozile
fagilor multipli care se fixeaza pe celula sensibila;
nucleaze care degradeaza cromosomul bacterian. Endonucleaza A creeaza brese
monocatenare la secventele cu citozina, iar endonucleaza B ataca secventele
monocatenare care contin citozina. Rezulta astfel, mici fragmente de ADN dublu
catenare, iar o exonucleaza fagica desavarseste procesul pana de dezoxi-nucleotid-
trifosfati (dNTP).
proteine care catalizeaza sinteza bazelor specifice ADN fagic (5- HMC);
proteine reglatoare ale transcrierii ADN fagic. ARNm viral timpuriu este transcris de ARN-polimeraza bacteriana, iar sinteza ARNm tardiv este catalizata de proteine virale. Unii fagi inactiveaza ARN-polimeraza celulara si codifica sinteza unei ARN-polimeraze timpurii proprii;
enzime care catalizeaza replicarea ADN fagic: ADN-polimeraza, polinucleotid-ligaza.
Sinteza ARNm celular este blocata prin schimbarea specificitatii ARN-polimerazei care trece sub controlul unui promotor viral. Sinteza proteinelor celulare este inhibata curand dupa infectie, deoarece ARNm bacterian are un timp de injumatatire foarte scurt. Astfel, ribosomii bacterieni devin disponibili pentru traducerea ARNm fagic.
Replicarea genomului fagic este mai bine cunoscuta pentru fagii T4 si l. ADN-T4 se sintetizeaza din nucleotidele rezultate din degradarea ADN celular, proces catalizat de endonuclezele den A si den B, codificate de fagul T4. Ele sunt active numai asupra ADN care contine citozina.
Hidroxi-metil-citozina (HMC) este sintetizata de doua enzime fagice. In sinteza ADN T4 trebuie prevenita incorporarea citozinei, deoarece o astfel de molecula este substratul endonucleazelor fagice care degradeaza ADN bacterian. Fagul trebuie sa codifice sinteza enzimelor care sa previna incorporarea C, in molecula de ADN.
E. coli poseda o endonucleaza(restrictaza) care cliveaza secventele cu HMC. Pentru a preveni degradarea ADN-T4 (cu HMC), resturile de HMC sunt glicozilate de doua enzime fagice (a si b-glicozil-transferaza). ADN T4 devine "imun" fata de enzimele de restrictie ale celulei bacteriene. Glicozilarea este o modificare post-replicativa, iar endonucleaza bacteriana este inactiva fata de ADN-glicozilat.
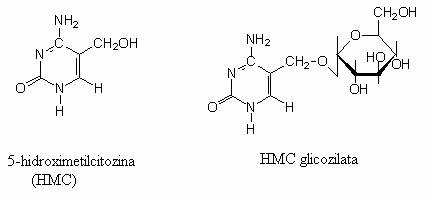
5-hidroxil-citozina neglicozilata si glicozilata. Prin inlocuirea gruparii CH2OH cu H, rezulta citozina.
ADN fagic, intr-o prima etapa, se replica dupa modelul semiconservativ bidirectional, al cercului simplu. Se sintetizeaza copii ale genomului care vor fi transcrise pentru sinteza proteinelor tardive. Ulterior, ADN fagic se replica dupa modelul cercului rotativ (fig. 98). Rezulta o molecula poligenomica de ADN (genomuri multiple asociate in conexiunea cap-coada), un genom concatemer. O endonucleaza cliveaza concatemerul, in molecule de lungimea genomului, ce vor fi incorporate in capsida, in procesul morfogenezei.
Sinteza proteinelor tardive, devine predominanta dupa circa 20 de minute, dupa ce ADN a atins rata maxima de replicare, iar transcrierea genelor timpurii este stopata. Proteinele tardive sunt grupate in trei categorii:
majoritatea sunt proteine structurale si intra in alcatuirea capului, cozii si anexelor ei;
proteinele de morfogeneza (de asamblare). Cele mai importante sunt proteinele de "cofraj", definite ca proteine necesare in timpul asamblarii unei structuri, dar lipsesc din structura virionului;
proteinele enzimatice, necesare lizei celulei bacteriene si eliberarii fagilor progeni: o muramidaza de tipul lizozimului, ce hidrolizeaza mureina peretelui bacterian si o lipaza (holina), care ataca membrana plasmatica a celulei, usurand accesul lizozimului spre structura parietala. Liza celulei este conditionata de acumularea acestor proteine.
Asamblarea si morfogeneza fagului s-a studiat la mutante fagice defective, care produc numai capete, numai cozi sau numai fibre. Fagul are o arhitectura complexa si principiul autoasamblarii nu este functional. S-au identificat 3 linii separate de asamblare a subunitatilor structurale ale fagului: linia capetelor, a cozilor, a fibrelor si a proteinei 63, care catalizeaza legarea fibrelor la nivelul placii bazale.
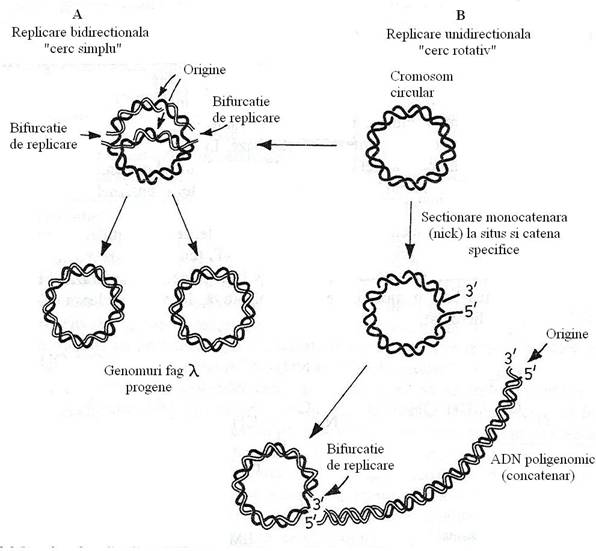
Fig. 98. Mecanismul replicarii ADN al fagului λ in cursul infectiei litice. a. Initial, replicarea se face dupa modelul "cercului simplu" (sinteza bidirectionala fata de punctul de origine al replicarii). B. Pentru asamblarea fagilor progeni, replicarea genomului se face dupa modelul "cercului rotativ" (dupa Watson, 1977).
Mai mult de 50 de gene ale fagului T4 sunt implicate in morfogeneza. Ele codifica proteinele structurale, dar si pe cele nestructurale, necesare asamblarii.
Prima regula a morfogenezei, este ca asamblarea se face pe module structurale. Capetele si cozile sunt primele care se asambleaza si formeaza complexe vizibile la microscopul electronic. Ulterior, fibrele cozii se adauga acestor complexe. Avantajul asamblarii fagului pe mai multe module, este acela ca subunitatile cu erori pot fi eliminate timpuriu, separat la fiecare linie.
A II-a regula este ca asamblarea se face de la capatul terminal, spre punctul de jonctiune cu restul virionului. De exemplu, pentru asamblarea cozii, primele sunt produse placile bazale, apoi cilindrul axial, iar in final, teaca.
In procesul asamblarii capului, prima structura care se formeaza se numeste precap tip I, ce se asambleaza pe o structura proteica centrala, formata din proteinele de "cofraj", pe care sunt inserate capsomerele si nu contine ADN. Aceasta structura este convertita in precap de tip II, de asemenea lipsit de ADN, dar mai consolidata prin aparitia legaturilor chimice intre capsomere. Structura capului isi dobandeste proprietatile structurale si functionale depline, inainte de patrunderea ADN (fig. 99).
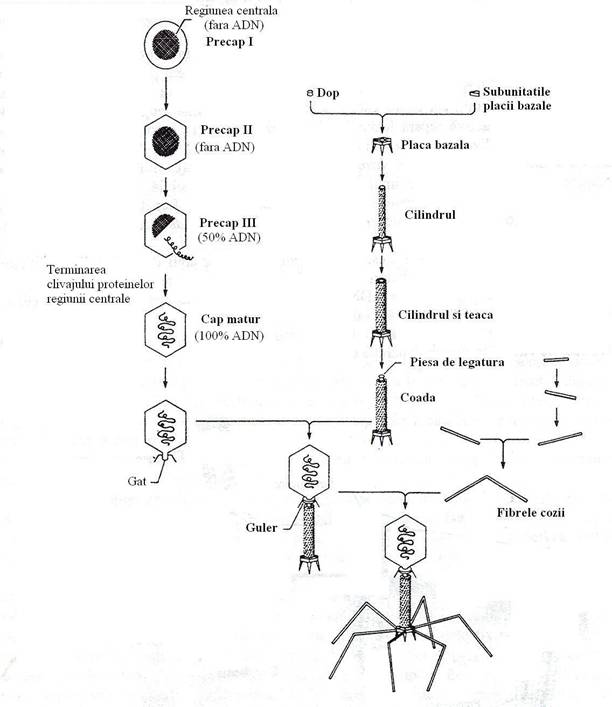
Fig. 99. Ilustrarea schematica a mecanismului asamblarii fagului T4. Asamblarea fagilor se face pe trei module: cap, coada si fibrele cozii (dupa Davis, 1990).
Impachetarea ADN, incepe prin legarea unui capat al moleculei de ADN poligenomic (concatemer), de o proteina a capului fagic. Molecula de ADN s-ar rula pe un ax proteic, intr-un mod particular, de la exteriorul spre interiorul axului. ADN patrunde pe la unul din varfurile icozaedrului, dupa dislocarea unei capsomere. La acelasi varf se asambleaza coada.
Poliaminele si o proteina de condensare, au rol in impachetarea ADN. Molecula de ADN este sectionata din concatemerii rezultati in replicare. In capul fagului T4 intra o cantitate de ADN mai mare decat un genom complet (l02%). Din aceasta cauza, in timpul impachetarii ADN, sectionarile din concatemeri, nu se fac la o secventa unica de baze, ci la pozitii determinate de cantitatea de ADN ce intra intr-un cap. Probabil, capatul liber al moleculei, intra in capul fagic si continua pana la umplere, dupa care concatemerul este sectionat. Acesta este mecanismul "capului plin". Secventa terminala impachetata este duplicata fata de secventa care a intrat initial, adica ADN este redundant terminal.
Deoarece in capsida fagului T4 este impachetata o molecula de ADN mai mare decat genomul propriu-zis, secventa nucleotidelor in genomul particulelor virale ale unei populatii, este defazata cu un numar determinat de baze. De exemplu, daca intr-o molecula de ADN genomic, secventa nucleotidelor este ABCD.. XYZAB, in altele secventa este CDEF... XYZABCD. Toate moleculele de ADN genomic contin aceiasi secventa de nucleotide, dar secventa initiala a genomului se repeta la capatul terminal si difera de la un virion la altul. Datorita redundantei terminale, genomul fagilor este permutat ciclic.
Redundanta terminala este o proprietate individuala a moleculelor de ADN fagic, iar permutarea ciclica este o proprietate a genomului populatiei de fagi.
Pe masura ce genomul este incorporat in capsida, proteinele de "cofraj" sunt eliminate prin spatiile dintre capsomere. Capul matur are un volum cu 40-l00% mai mare decat al precapului.
Fagii maturi se acumuleaza in celula bacteriana, iar cromosomul bacterian pierde treptat coordonarea activitatii celulare si apare un dezechilibru functional profund. Muramidaza se sintetizeaza foarte timpuriu, la 8 minute dupa infectie, dar este ineficienta atata timp cat celula este fiziologic activa. Enzima sectioneaza moleculele de mureina, dar celula activa repara leziunile. Cand leziunile nu mai pot fi reparate in ritmul producerii, celula se lizeaza exploziv si elibereaza fagii progeni. Liza celulei necesita actiunea sinergica a doua produse genice: lizozimul si holina (o lipaza). Holina creeaza brese in membrana interna, permitand lizozimului sa ajunga la structura sensibila.
Fiecare virion progen poate sa infecteze o noua celula. Repetarea de mai multe a ciclului litic pe o panza de celule sensibile, produce o plaja de liza. Plaja de liza este echivalentul unei colonii de fagi.
In cultura bacteriana crescuta intr-un mediu lichid, fagii determina liza celulelor, astfel incat suspensia bacteriana se clarifica, mergand pana la sterilizare.
Fagii icozaedrici cu genom ADN monocatenar au dimensiuni mici (25-35 nm), sunt totdeauna virulenti ca si cei cu genom ADN dublu catenar si lizeaza celula in circa 30 de minute. Prototipul lor este fagul fi X l74. Capsida este alcatuita din trei tipuri de proteine (F, G, H). Proteinele G si H se termina cu prelungiri in forma de spiculi, necesara atasarii pe suprafata celulei.
In celula infectata, ADN se tapeteaza cu proteina SSB (Single Stranded Binding) si incepe sa se replice. Initial, se sintetizeaza o molecula de ARN scurta, catalizata de ARN-polimeraza celulara, cu rol de primer. Complexul celular de replicare extinde primerul, iar ulterior ARN este digerat. Rezulta o molecula ADN dublu catenara, care este forma replicativa.
Fagii filamentosi (prototipul este fagul fd) au ca genom o molecula de ADN circulara, care se dubleaza, prin pliere fata de sine insasi. Bazele nu sunt complementare, ceea ce inseamna ca ADN este monocatenar.
Virionul se adsoarbe la extremitatea unui pil F. Mecanismul patrunderii ADN in celula nu se cunoaste. ADN monocatenar este tapetat cu o proteina codificata de fag, cu rol major, se pare, in impachetare si nu in replicare. Impachetarea genomului in capsida, este concomitenta cu iesirea virionului. Proteina majora a capsidei helicale este inclusa in membrana celulara. Pe masura ce ADN fagic patrunde in membrana, este acoperit de proteina capsidei. Sunt singurii fagi care nu necesita liza celulei pentru eliberare. Celula continua sa creasca, sa se divida si sa elimine cateva sute de virioni la fiecare generatie celulara.
Fagii ARN "masculi" (prototipul este MS2) au simetrie icozaedrica. Genomul este o molecula de ARN monocatenar, de 4 kb lungime si contine 3-4 gene. Receptorii se gasesc pe suprafata pilului F si de aceea infecteaza numai celulele bacteriene F+, cu potentialitate de donor de material genetic in cuplul de conjugare. De aici deriva denumirea de fagi "masculi". Cele 4 gene codifica o proteina capsidala (C), o ARN-polimeraza dependenta de ARN (P), o lizina (L) si proteina A, reglatoare a asamblarii, prezenta intr-o singura copie in virion.
Toate copiile de ARN sunt identice, fie ca au rol de genom, fie ca sunt mesagere.
Importanta fenomenului de bacteriofagie
Bacteriofagii sunt foarte raspanditi: se gasesc in mediile naturale in care traiesc bacteriile sensibile care le pot fi gazda, adica in apa, sol, in tractul digestiv al omului si animalelor. Datorita efectului litic al interactiunii cuplului, fagii regleaza densitatea populatiilor bacteriene in mediile naturale. Interactiunea litica a cuplului fag-bacterie are cateva aplicatii practice majore.
Fagul este utilizat in terapeutica, pentru tratamentul unor infectii rezistente la antibiotice si la agentii chimioterapeutici.
Specificitatea relatiei fag-bacterie, a stat la baza elaborarii unui procedeu de identificare si clasificare a bacteriilor, in functie de sensibilitatea lor la un fag sau la un grup de fagi, procedeu denumit lizotipie sau tipizare prin fag. Metoda este folosita pentru stabilirea diagnosticului bacteriologic in clinica, dar si in studiile de sistematica bacteriana.
Prin tipizare fagica se intelege identificarea unei tulpini bacteriene, utilizand un set de preparate fagice purificate si standardizate, carora li se cunoaste capacitatea de a liza anumite tulpini bacteriene. Tipizarea prin fag este o metoda foarte utila in studiile de epidemiologie, pentru a reconstitui originea si circulatia unei infectii intr-o colectivitate, in cursul unei epidemii. Practic, toate tulpinile unei specii bacteriene patogene, izolate in cursul unei epidemii de la bolnavi, de la purtatori, din alimente contaminate, din vectori sau de animalele care actioneaza ca izvor de infectie, avand origine comuna, apartin aceluiasi lizotip (aceluiasi tip fagic).
Metoda se foloseste pentru identificarea tulpinilor de Salmonella, Vibrio etc. In acest scop, tulpinile se insamanteaza in placi, dupa tehnica "in panza". Se testeaza sensibilitatea lor fata de un numar cat mai mare de preparate fagice standardizate.
In bioindustrie, fagii pot produce liza microorganismelor producatoare de substante utile, provocand pierderi economice importante.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 2597
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved