| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Transductia genetica
Transductia este procesul de transfer al materialului genetic intre celulele bacteriene, mediat de bacteriofagi. Zinder si Lederberg (l952) au descoperit transductia generalizata la Salmonella typhimurium. Ei au pus in contact mutante auxotrofe, cu diferite incapacitati de sinteza si ulterior, au izolat, pe un mediu minimal, colonii recombinante prototrofe. Pentru a-l deosebi de conjugare, ei au demonstrat ca acest proces genetic nu necesita contactul fizic dintre celula donoare si cea receptoare de material genetic. Agentul mediator al transferului genic, trece prin porii filtrului, suficient de mici pentru a opri bacteriile. Procesul nu s-a datorat nici transformarii genetice, deoarece este rezistent la DN-aza. Vectorul purtator de gene, de la celula donoare la cea receptoare a fost fagul temperat P22. Profagul P22 a fost indus spontan si astfel celulele lizogene au evoluat spre liza. S-au asamblat particule virale transductoare, care au infectat celulele celei de a II-a tulpini de Salmonella din amestecul celular. Particulele virale erau purtatoare ale genei bacteriene de tip salbatic, care s-a recombinat cu cromosomul celulelor mutante (auxotrofe), rezultand astfel celule prototrofe, care cresc si se divid pe mediul minimal si formeaza colonii.
In raport cu diversitatea genelor transduse si cu gradul de manifestare a efectului transductiei asupra celulelor descendente, se descriu trei variante ale procesului;
transductie specializata
transductie generalizata
transductie abortiva.
Transductia specializata a fost descoperita de Lederberg (l953). Este caracteristica fagului λ, ca o consecinta a faptului ca genomul sau se integreaza riguros, numai intre genele gal si bio ale cromosomului de E. coli, in raport cu care prezinta omologie genetica. Integrarea este rezultatul recombinarii genetice la situsuri specifice (fig. 134).
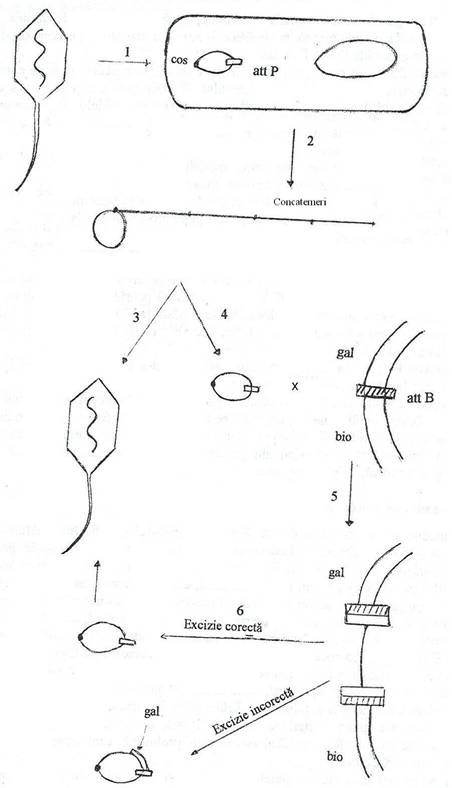
Fig. 134. Ilustrarea schematica a procesului de transductie genetica. Dupa ce fagul λ infecteaza o celula de E. coli, genomul sau se circularizeaza si cuplul evolueaza in doua directii alternative: 1) evolutia litica, in cursul careia cromosomul se replica si formeaza concatemeri ce vor fi clivati si impachetati in particule fagice progene; 2) evolutia lizogena, in cursul careia fagul se integreaza in cromosomul celulei, prin recombinare la situs specific. Sub actiunea unui factor inductor, genomul fagic se excizeaza printr-un proces invers celui de integrare. In situatii rare, procesul de excizie este aberant (incorect), adica genomul fagic incorpopreaza secvente cromosomale ale celulei gazda, dar pierde o secventa de marime echivalenta. Daca genele fagice nu sunt esentiale pentru ciclul de multiplicare, cromosomul fagic recombinat se replica si se asambleaza fagi progeni transductori
In stare integrata, profagul se comporta ca orice gena cromosomala, adica se replica sincron cu cromosomul si este transmis tuturor celulelor descendente.
Sub actiunea unor factori fizici (socuri termice, radiatii UV etc.) sau chimici, genomul viral este excizat din insertiile sale cromosomale, se circularizeaza si se replica productiv, generand fagi progeni. Procesul se numeste inductie litica, deoarece rezultatul este liza celulei bacteriene. Inductia litica este consecinta anihilarii actiunii proteinei represor, singura codificata de genomul fagc integrat. Excizia genomului fagic are loc sub actiunea unor enzime si de regula, sectionarea se face la situsurile cos care flancheaza genomul fagic, ceea ce corespunde unei excizii corecte. Proteina Rec A se activeaza si devine proteaza, substratul actiunii sale fiind represorul codificat de fagul lambda, iar excizionaza (o endonucleaza) sectioneaza genomul fagic. Excizionaza formeaza un complex functional cu integraza, ce se leaga strans de situsul POB'' Actiunea lor combinata determina excizia profagului.
Cu o frecventa mica (l/l06 celule), excizia genomului fagic este incorecta. Astfel, se excizeaza un genom fagic care poarta gene bacteriene, dar caruia ii lipseste o secventa echivalenta de nucleotide virale, la extremitatea opusa. Genomul viral modificat, se replica, este tradus la proteine virale si se asambleaza fagi transductori.
Fagul λ este excizat impreuna cu gena cromosomala gal, bio, ori cu ambele. Datorita integrarii genelor bacteriene in cromosomul fagic, are loc pierderea (prin recombinare) unei secvente echivalente ca marime, din genomul fagic. Rezulta astfel, fagi transductori defectivi, deoarece cantitatea de material genetic ce poate fi impachetata in capul fagului este constanta. ADN viral este pierdut totdeauna la extremitatea opusa genelor bacteriene dobandite de fagul transductor. Foarte rar pot fi generati fagi transductori, care contin genomul viral complet, impreuna cu secvente de ADN bacterian, derivate de la ambele extremitati ale punctelor de insertie a profagului.
In general, fagii transductori sunt defectivi si in consecinta, incapabili sa se multiplice si sa lizeze celula bacteriana. De aceea, pentru a initia ciclul de multiplicare si liza celulei bacteriene, necesita prezenta fagilor helper.
In celula infectata, de cele mai multe ori, genomul fagului transductor este supus actiunii enzimelor de restrictie si este degradat.
Cu o frecventa mica, genomul fagului transductor se poate integra in cromosomul bacterian, printr-un proces de recombinare si genele transduse pot conferi noi caractere fenotipice celulei bacteriene. De exemplu, fagul transductor defectiv lambda dgal+, dupa ce infecteaza si genomul sau se recombina cu cromosomul unei bacterii defective pentru utilizarea galactozei (gal-), ii confera acesteia capacitatea de a metaboliza galactoza. Celula bacteriana care a dobandit un fenotip nou se numeste transductant. Fragmentul de material genetic transdus de genomul fagic este adaugat genomului bacterian, si nu substituit.
Transductia fagica este un proces care se realizeaza cu o frecventa mica, dar datorita numarului mare de particule virale si a celulelor bacteriene infectate, aceasta modalitate de transfer de material genetic intre celulele bacteriene, are rol semnificativ in conditii naturale.
Transductia generalizata, nelimitata sau nerestrictiva este procesul de transfer, prin intermediul unui fag, a oricarei gene a cromosomului bacterian, indiferent de pozitia sa in genom. Procesul a fost descris pentru fagul P1 care infecteaza E. coli, pentru fagul P22 la Salmonella si pentru unii fagi care infecteaza celulele de B. subtilis.
Fagii de transductie generalizata au proprietatea, ca in ciclul litic in raport cu celula bacteriana, sa incorporeze fragmente cromosomale ale gazdei.
Virusul P1 are ca genom o molecula de ADN lineara, dublu catenara si o capsida icozaedrica. ADN, de l00 kb este permutat ciclic si are secvente repetitive terminale. Ca si fagul T4, dupa infectie molecula se circularizeaza.
Cuplul poate urma ciclul litic sau lizogenic. Varianta lizogena necesita replicarea ADN circular, in stare fizic autonoma. In stare lizogena, fagul P1, sub forma moleculei circulare este mentinut intr-un numar minim de copii (una/celula), datorita unui control stringent al replicarii, ca si in cazul plasmidei F. Mecanismul distributiei poate deveni ineficient daca un proces de recombinare omologa a creat o molecula dimerica, ce se distribuie numai in una din cele doua celule fiice.
In evolutia litica, ADN fagic se replica dupa modelul theta (θ) si al cercului rotativ. Rareori, in procesul asamblarii, sunt incorporate si fragmente de ADN ale cromosomului bacterian, rezultand fagi de transductie generalizata.
Infectia celulelor de Salmonella cu fagul P22 produce fragmentarea ADN celular. In cursul morfogenezei virale, in capside se impacheteaza uneori, exclusiv ADN bacterian si rezulta fagi transductori. In capsidele goale sunt incorporate, de regula, numai gene de provenienta bacteriana si numai in mod exceptional sunt incorporate gene celulare in asociatie cu mici fragmente de genom viral. Teoretic, oricare dintre genele cromosomale, are o sansa egala de a fi transdusa (de aceea se numeste transductie generalizata), desi, practic, unii cistroni sunt transdusi cu o frecventa mai mare. In capsida fagica sunt impachetate preferential, fragmentele de ADN care au o secventa similara secventei "pac" (packing =impachetare), care regleaza impachetarea genomului fagic. Aparatul enzimatic de impachetare a genomului fagic recunoaste secventa anologa si astfel fragmentele de ADN bacterian sunt incorporate in capsida virala. Rezulta astfel, particule fagice care poarta diferite fragmente ale cromosomului bacterian. Desi in celula exista cantitati aproximativ egale de ADN fagic si ADN celular, numai un mic procent al fragmentelor cromosomului bacterian este impachetat si un procent mic de particule virale sunt transductoare. Factorul limitant al asamblarii fagilor transductori este frecventa secventelor pac.
Particulele fagice care incorporeaza exclusiv sau preponderent gene bacteriene sunt totdeauna defective. Acesti fagi nu sunt obligatoriu lizogenizanti. Dupa ce infecteaza o noua celula bacteriana, nu se multiplica si nu au efect litic, deoarece lipsesc genele virale esentiale. Genele transduse pot fi integrate prin procese de recombinare, in cromosomul bacterian. Procesul se realizeaza prin inlocuirea unor determinanti genetici ai celulei receptoare, cu cei transdusi. Recombinarea necesita omologie genetica intre secventele de ADN participante si este dependenta de proteina Rec A.
Transductia abortiva (incompleta) se caracterizeaza prin faptul ca genomul virusului transductor nu determina liza, dar nici nu se integreaza in cromosomul celulei infectate.
Recombinarea fragmentelor de ADN transduse, de origine bacteriana, are loc cu o frecventa foarte mica. Majoritatea fragmentelor transduse raman in citoplasma celulelor receptoare. Nu se replica, deoarece nu contin o origine a replicarii si astfel sunt mostenite numai de una din cele doua celule surori care rezulta la fiecare diviziune. Totusi, sunt gene functionale si codifica produsul lor in celula receptoare, in care determina o stare de diploidie partiala. Genele transduse se dilueaza treptat in populatia celulara, prin diviziune.
Daca celula este mutanta nutritionala (datorita mutatiei unei gene cromosomale), iar fragmentul transdus isi are originea intr-o celula prototrofa, se produce complementarea genica si reversia la prototrofie a celulei receptoare. La fiecare diviziune celulara, produsul de sinteza a genei transduse poate fi distribuit in cele doua celule surori si astfel, celula care nu primeste fragmentul de ADN, fenotipic poate ramane de tip salbatic, pana cand produsul este degradat sau diluat prin diviziuni succesive. Astfel de celule produc colonii mici, spre deosebire de transductantii stabili, care produc colonii de dimensiuni normale.
Un alt exemplu al transductiei abortive este acela al genelor care determina sinteza si asamblarea flagelului. In acest caz, merodiploidul format prin transductia abotiva este mobil, iar celula sora rezultata prin diviziune, este imobila. Pe o placa cu agar moale, merodiploidul este mobil, iar celulele imobile formeaza o colonie tipica.
Transductia plasmidelor
Asa cum cromosomul bacterian poate fi impachetat in fagii de transductie generalizata, in mod asemanator ADN plasmidial poate fi inglobat in capsidele virale. Daca plasmida este mai mare decat capul fagic, se impacheteaza numai o parte a plasmidei. Plasmida R este mai mare decat capul fagului P22. Fragmentul de plasmida transdus nu se poate circulariza (nu are secvente omologe de baze) si nici nu se poate integra in cromosomul gazdei si de aceea va fi degradat. Plasmidele prea mici nu produc un cap fagic stabil, dar formele lor multimere pot fi impachetate eficient.
Transductia este un proces care are loc si la bacteriile Gram pozitive: Mycobacterium, Staphylococcus, Streptomyces si B. subtilis.
Conversia fagica
Conversia fagica este procesul aparitiei unor proprietati fenotipice noi ale celulei bacteriene, consecutiv intrarii in actiune a unor gene fagice. Fenomenul conversiei fagice se manifesta la unele bacterii infectate cu un fag temperat netransductor, care persista in celula, fie in stare de profag (integrat in cromosomul bacterian), fie in stare fizic autonoma.
Conversia fagica se deosebeste de transductia fagica specializata, prin doua particularitati esentiale:
modificarile celulei bacteriene sunt rezultatul expresiei unor gene de origine virala (si nu bacteriana, ca in cazul transductiei fagice);
fagii care realizeaza fenomenul de conversie sunt normali si poseda tot setul de gene, in timp ce fagii de transductie specializata sunt defectivi, datorita pierderii unor gene proprii, prin inlocuirea lor cu gene ale cromosomului bacterian.
Fenomenul conversiei fagice este cunoscut la Corynebacterium diphteriae. Celulele care au ca material genetic numai propriul cromosom nu sunt virulente si nu determina un proces patologic infectios. Aparitia virulentei este conditionata de infectia cu fagul beta, al carui genom se integreaza in cromosomul bacterian. Sinteza toxinei difterice este codificata de o gena fagica.
Un alt exemplu este acela al structurii chimice a lipopolizaharidului din peretele celular de Salmonella. S-a descris fenomenul de conversie serologica a unei linii de S. anatum, dupa infectia cu un fag, ca o reflectare a modificarii compozitiei chimice a polizaharidului din molecula de lipopolizaharid. Dupa pierderea profagului prin excizie, liniile bacteriene respective revin la fenotipul anterior infectiei fagice: C. diphteriae pierde capacitatea de a sintetiza toxina (isi pierde patogenitatea), iar S. anatum revine la tipul serologic initial. Se considera ca aceste gene fagice, la origine au fost gene cromosomale, dar au fost incorporate si au devenit gene fagice.
Transfectia
Transfectia este o varianta a transformarii genetice, prin intermediul acizilor nucleici de origine virala. In acest scop se poate folosi atat ADN cat si ARN.
Transfectia a fost definita de Foldes si Trautner (l964). Ei au tratat suspensia de celule bacteriene cu ADN fagic purificat. Dupa cultivarea celulelor pe suprafata placilor cu mediu solidificat, au observat aparitia plajelor de liza, al caror numar este proportional cu cantitatea de ADN adaugat suspensiei celulare.
Transfectia a fost realizata, in special, la bacterii Gram negative: E. coli, A. tumefaciens, E. aerogenes, P. aeruginosa, S. typhimurium, dar si la B. subtilis.
La E. coli, transfectia a fost descrisa de Kaiser si Hogness (l960), ca rezultat al interactiunii dintre ADN purificat al fagului λ transductor (λ dgal), cu celulele lizogenizate de fagul λ. Transfectia este usurata daca celulele sunt infectate simultan cu un fag helper netransductor. Fagul helper asigura perforarea peretelui celular si probabil ca pe masura ce fagul injecteaza propriul sau ADN, situsul cos al ADN de transfectie se asociaza cu ADN al fagului helper si astfel moleculele sunt transportate simultan in celula.
Deoarece fagii transductori contin gene cromosomale bacteriene, transfectia realizeaza concomitent si procesul transformarii genetice.
La E. coli, transfectia, similar transformarii, se poate realiza dupa tratarea celulelor cu o solutie de Ca2+ 0,025 M, care asigura o permeabilizare a invelisurilor celulare fata de ADN de diferite proveniente.
Proportia celulelor care incorporeaza ADN fagic creste semnificativ dupa tratamentul cu lizozim si EDTA, in urma caruia rezulta sferoplasti.
Fluxul vertical al informatiei este procesul de transmitere a genelor de la parinti la descendenti. Diversificarea moleculara rezulta prin procese mutationale si prin duplicatie genica, rezultatul fiind diversificarea organismelor si speciatia. Fluxul vertical al genelor este cel care determina arborele filogenetic universal sau tabloul evolutiei canonice.
La nivel inferior, arborele evolutiei canonice arata dichotomia veche intre Bacteria si Archaea si evolutia ulterioara a Eucarya din ramura Archaea. Moleculele care reflecta evolutia canonica sunt rezultatul fluxului vertical al genelor. Fluxul genetic vertical contribuie la diversificarea genetica lenta, prin procesul denumit descendenta cu modificare, care in timp duce la divergenta de mari dimensiuni.
Transferul orizontal al genelor (HGT) este transferul elementelor genetice intre organisme diferite si constituie cel de al II-lea tip de flux genetic. Efectul HGT poate fi limitat, cand are loc intre bacterii strans inrudite, dar este mult mai amplu cand are loc intre specii sau genuri diferite.
Transferul orizontal al genelor este o modalitate de amestec rapid al materialului genetic, capabil sa introduca modificari genetice rapide. HGT, foarte timpuriu in evolutie, de la o bacterie la un eucariot, poate avea efecte deosebit de ample.
Rolul transferului orizontal interspecific de gene este subliniat de faptul ca circa 17% din genele cromosomului de E. coli K12 deriva din surse colaterale neinrudite.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 3757
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved