| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Miscarea este o insusire esentiala a materiei vii. La animale miscarea este evidenta, mai ales prin deplasare. La plantele superioare fixate de substrat, miscarea nu se efectueaza prin deplasare, ci prin cresterea sau curbarea organelor sub actiunea unor factori de mediu. Miscarile plantelor sunt numite miscari de crestere.
Orientarea cresterii plantelor spre zonele cu conditii optime sau indepartarea de cele cu conditii nefavorabile se realizeaza cu participarea hormonilor de cresterte sau prin modificarea turgescentei unor grupuri de celule.
Miscarile active ale plantelor superioare se realizeaza cu consum de energie. Efectuarea acestor miscari necesita un mecanism in lant care include factorii excitanti, receptorii, sistemele de transmitere a excitatiilor si realizarea miscarilor de raspuns.
Excitantii reprezinta, in general, factorii mediului inconjurator: umiditatea, substantele minerale, temperatura, forta de gravitatie, excitatiile mecanice, lumina etc. Pentru a fi receptionate, excitatiile trebuie sa depaseasca o valoare minima de prag a intensitatii.
Receptarii sunt diferentiati in functie de factorii excitanti:
Stimulii luminosi sunt receptionati de pigmentii fotoreceptori, respectiv de fitocrom si de flavoproteine.
Forta de gravitatie determina sedimentarea amiloplastelor si a unor cristale anorganice, care excita receptorii situati in plasmalema si in reticulul endoplasmatic.
Excitatiile mecanice sunt receptionate de celele senzitive sau de papile senzitive, situate prin celulele epidermice.
Transmiterea excitatiei se face prin intermediul hormonilor, prin modificarea permeabilitatii membranelor plasmatice sau sub forma unui potential de actiune. Executarea miscarilor de orientare a organelor plantelor se realizeaza prin cresterea inegala a celulelor ca urmare a actiunii hormonilor sau prin modificarea turgescentei celulelor ca urmare a functionarii pompelor de ioni.
In cazul plantelor superioare miscarea poate fi activa., - atunci cand se realizeaza cu consum de energie metabolica si pasiva, - cand nu se consuma energie. Realizarea miscarilor presupune existenta unui mecanism complex, ce actioneaza in lant pornind de la factorul excitant la receptor prin sistemele de transmitere a impulsului pana la realizarea miscarii de raspuns conform schemei (fig. 179).
De obicei excitantii sunt reprezentati de factori ai mediului precum; lumina, temperatura, gravitatia. apa, etc. Receptorii de care dispun plantele sunt extrem de diferiti, functie de stimulii externi, iar caile de transmitere a excitatiilor sunt reprezentate de substantele bioregulatoare de crestere naturale (fitohormonii) (Milica si colab., 1982).
Mecanismele moleculare ale miscarilor plantelor sunt departe de a fi pe deplin intelese. Cercetarile recente au realizat totusi progrese remarcabile, dezvoltand modele explicative ce confera un rol esential in realizarea miscarilor intracelulare si directionale elementelor contractile celulare (microtubuli si/sau microfilamente). Un alt mecanism important in realizarea miscarilor se bazeaza pe schimbarea turgescentei tesuturilor, prin modificarea presiunii osmotice a celulelor implicate in realizarea miscarii ( Stubbs and Slabas, 1982; Versel and Pilet, 1986).
11.1. Miscarile pasive
Miscarile pasive sunt frecvent intalnite si se bazeaza pe mecanismele fizice generate de diferenta de umiditate existenta intre tesuturi vegetale invecinate sau intre tesuturi si mediul extern. Acest tip de miscari sunt importante mai ales datorita faptului ca permit perpetuarea speciei prin dispersia semintelor, a polenului, a fructelor, etc.
- miscari pasive prin imbibitie - sunt caracteristice fructelor uscate si au loc datorita modificarilor inegale a volumului diferitelor tesuturi ce intra in constructia organului, sub actiunea apei de imbibitie anizotropa din micelele celulozice ale peretilor celulari. Exemplu - pastaile de Orobus vernus ce se desfac pe timp de seceta prin spiralizare spre exterior.
- miscari pasive prin coeziune - caracteristice sporangiilor ferigilor, si au loc datorita forei mari de coeziune dintre apa si membranele celulare (peste 1000 atm).
Aceste miscari au loc si in cazul deschiderii anterelor. Celulele endoteciului anterelor prezinta ingrosari sub forma de stea pe fata interna si sub forma de cordoane pe partea externa. Atunci cand are loc deshidratarea, membranele externe subtiri si elastice sunt trase spre interior, generand deschiderea anterei.
- miscarile pasive prin aruncare - sunt caracteristice staminelor unor specii de plante din familia Urticaceae. La maturarea polenului anterele concrescute la baza filamentelor se desprind si executa o miscare brusca de intindere aruncand astfel polenul la o anumita distanta
Din cele prezentate se remarca faptul ca in cazul miscarilor pasive mecanismele miscarii au la baza modificari ale umiditatii unor tesuturi sau componente celulare.
11.2. Miscarile active
Miscarile active sunt frecvente si au loc cu consum energetic. Din aceasta categorie fac parte miscarile intracelulare, cele ale plantelor fixate de substrat si miscarile plantelor libere.
- miscarile active intracelulare - cuprind deplasarea unor organite celulare (nucleu), si citoplasmatice (cloroplastelor, hialoplasmei, veziculelor, etc). Miscarea nucleului este evidenta la nivelul perilor absorbanti ai radacinii si in tuburile polinice, unde nucleii se deplaseaza spre zonele cu activitate metabolica mai intensa, sau in zonele lezate ale celulei coordonind procesele metabolice sau reparatorii (Pop, 2002).
Miscarea cloroplastelor este prezenta atat la plantele superioare cat si la cele inferioare. Scopul miscarii cloroplastelor este de a capta in conditii optime lumina pentru realizarea fotosintezei.
Miscarea ce sta la baza miscarii cloroplastelor este reprezentat fie de catre curentii protoplasmatici, fie prin modificarea tensiunii superficiale dintre membrana externa a organitului si hialoplasma, fapt ce genereaza anumite forme de miscare.
Miscarea hialoplasmei se realizeaza prin curentii protoplasmatici. Mecanismul prin care se realizeaza miscarile intracelulare consta in contractarea si relaxarea succesiva a microfibrilelor proteice constitutive. Exista de asemenea miscari la nivelul reticului endoplasmatic (sistemul circulator celular) la nivelul diferitelor vezicule, cisterne, saculi, etc. Viteza cu care se realizeaza procesele de miscare intracelulare este direct dependenta de gradul de hidratare al protoplasmei, deci de activitatea fiziologica a celulei.
11.3 Miscarile plantelor fixate de substrat.
Majoritatea speciilor vegetale, deci plantele fixate de substrat realizeaza in tot cursul dezvoltarii miscarii active ca raspuns la actiunile unor stimuli externi. In urma actiunii stimulilor externi au loc modificari in modul de orientare al organelor si acestea pot fi paralelotrope (orientarea organului paralel cu functia excitantului) sau plagiotrope (orientarea organului perpendicular de directia excitantului). Cand organul in crestere se orienteaza spre excitant, miscarea este pozitiva, iar cand se orienteaza in sens invers, misarea este negativa
Realizarea miscarilor de crestere si orientare a diferitelor organe ale plantelor are loc prin cresterea inegala a celulelor ca urmare a actiunii hormonilor sau prin modificarea turgescentei celulelor ca urmare a functionarii pompelor de ioni. Principalele tipuri de miscare la plantele fixate sunt: tropismele, nastiile si nutatiile.
TROPISMELE
Tropismele sunt miscari orientate de crestere provocate de actiunea unilaterala a unui factor de mediu.
In functie de directia de crestere, tropismele pot fi:
pozitive, cand organul de crestere se orienteaza in directia de actiune a factorului de mediu;
negative, cand organul de crestere se orienteaza opus directiei de actiune a factorului de mediu.
Orientarea organului se face prin curbarea sa, datorita cresterii inegale a celulelor pe fetele opuse.
Dupa natura factorului de mediu, tropismele pot fi: geotropisme, fototropisme, hidrotropisme, chimiotropisme si tigmotropisme.
Geotropismul
Geotropismul (gravitropismul) este miscarea orientata de crestere provocata de actiunea unilaterala a fortei de gravitatie. Geotropismul orienteaza organele plantelor pe directia de actiune a fortei de atractie a pamantului, ceea ce explica pozitia orizontala a tulpinii aeriene si radacinii plantelor.
Plantele au dezvoltat de-a lungul timpului celule specializate (statocite) capabile sa perceapa gravitatia si sa raspunda stimulului acesteia prin semnale la nivelul zonelor de crestere. Statocitele detin, spre deosebire de alte celule un numar important de particole mobile, influentate de gravitatie ( teoria statolitilor) (Wayne si colab., 1992 Kiss si colab., 1996).
Statolitii sunt tipuri specializate de amiloplaste implicate in perceptia fortei gravitationale la plante. Amiloplastele specializate au o densitate mai mare decat hialoplasma, sedimentand sub influenta fortei gravitationale. Au rolul de a percepe si transmite stimulul gravitational prin activarea canalelor mecanosenzitive, fapt ce determina modificarea concentratiei de auxine la nivelul varfurilor de crestere ale sistemului radicular. Aceste modificari duc la cresterea diferita a tesuturilor sistemului radicular ( Evans si colab. 1986, Wang si colab. 1995).
Celulele receptoare (statociste) pentru forta de gravitatie se afla situate in meristemele apicale si in meristemele inercalare ale plantelor graminee (Kaufan s.a. 1987). Sub actiunea fortei de gravitatie, continutul statocistelor se repartizeaza neuniform, spre partea inferioara a acestora.
La modificarea pozitiei plantelor, amiloplastele si unele cristale anorganice, care au rolul unor statolite, ajung la partea bazala a celulelor in cca 10 min. Acestea exercita o presiune asupra reticulului endoplasmatic si a plasmalemei, care contin receptori de presiune (Kaufman si Dayanandan, 1985).
Realizarea miscarilor geotropice are loc in zona de extensie celulara care se afla la cativa milimetri de meristemul apical sau la baza internodurilor, in cazul plantelor graminee. Pana in prezent insa nu s-a putut preciza semnalul care se transmite de la statociste la zona de crestere si modul de transmitere a acestuia.
Aceasta crestere a organelor plantelor influentata gravitational, ce poarta denumirea de gravitropism, este un element esential in viata plantelor facilitand captarea luminii sau absorbtia apei si a elementelor minerale, depasind eventuale obstacole .
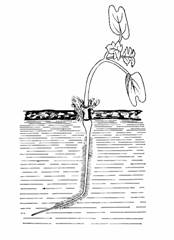
Raspunsul gravitropic al organelor plantelor se manifesta foarte devreme. Spre exemplu, semintele in curs de germinare apartinind speciei Hordeum vulgare, cu o lungime a radiculei de doar 2 cm sunt deja capabile sa raspunda actiunii fortei gravitationale (fig. 180).
Mecanismul gravitropismului la plantele superioare se bazeaza pe faptul ca acestea sesizeaza directia actiunii fortei gravitationale prin statocite, in zona elongatiei celulare. Atat calciul, cat si fitohormonii de tipul auxinei actioneaza ca raspunsuri la actiunea stimulului, iar concentrarea lor inegala in parti diferite ale aceluiasi tesut, determina cresteri inegale redirectionand sensul cresterii radacinii (Konings 1995).
In modificarea pozitiei organului, amiloplastele si unele cristale anorganice ajung in partea bazala a celulelor, presand asupra reticulului endoplasmatic si plasmalemei ce contin receptori de presiune (Kiss, 1996) ( fig. 181).
Mecanismul geotropismului este explicat prin participarea stimulatorilor de crestere.
Auxinele sunt sintetizate in frunze de unde migreaza in forma inactiva spre zonele apicale din varfurile radacinii si tulpinii. La acest nivel devin active, sub forma acidului -indolil acetic si sunt capabile sa circule in planta polar, in functie de potentialul bioelectric.
In pozitie verticala potentialul bioelectric al plantei este reprezentat de sarcini electrice negative la varfurile radacinii si tulpinii si de sarcini electrice pozitive la baza organelor, in zona numita colet. Auxinele electronegative datorita caracterului acid circula in planta in sens bazipetal, de la varful organelor spre colet. Circulatia este uniforma pe toate fetele organelor determinand o crestere uniforma a celulelor, din care rezulta pozitia verticala a radacinii si tulpinii.
In pozitie orizontala are loc modificarea potentialului bioelectric al plantei, prin repartizarea sarcinilor electrice negative pe partea superioara si a sarcinilor electrice pozitive pe partea inferioara. Auxinele electronegative se acumuleaza pe partea inferioara determinand la tulpina alungirea mai intensa a celulelor si curbarea acesteia in miscarea de geotropism negativ.
Pentru radacini concentratia auxinelor acumulate pe partea inferioara este supraoptimala, inhiband alungirea celulelor. Din aceasta cauza, celulele se alungesc mai intens pe partea superioara, determinand curbarea radacinii in miscarea de geotropism pozitiv.
Cercetarile efectuate de Bandurski, Schultz si Cohen (1980), la tulpinile de graminee asezate paralel cu suprafata solului, au evidentiat ca auxinele se acumuleaza in cantitate mai mare in zona situata in apropierea solului (349 ng/g substanta uscata). Cantitatea de auxine din aceasta zona a fost de 2,7 ori mai mare comparativ cu cea determinata in tesuturile situate in partea opusa a tulpinii. Distributia neuniforma a auxinei si anume in cantitate mai mare spre sol determina stimularea extensiei celuleor din aceasta zona si realizarea unei curburi geotrop negative.
Pharis s.a. (1981) au determinat o cantitate mai mare de gibereline libere in statocistele gravistimulate, comparativ cu cele nestimulate, iar Reid si Paris (citati de Purohit, 1985) au determinat intensificarea biosintezei etilenei.
Wlikins (1979) considera ca orientarea geotrop pozitiva a radacinilor se datoreste interactiunii dintre auxina transportata bazipetal si acidul abscisic transportat acropetal. Radacina este sensibila la actiunea auxinei care determina inhibarea cresterii acesteia. In acelasi timp, auxina accentueaza efectul inhibitor al acidului abscisic asupra cresterii.
Miscarile gravitrop au la baza un mecanism fiziologic si biochimic complicat, care include, dupa Kaufman s.a. (1987), biosinteza unor enzime ( -glucozidaza, celulaza si invertaza), functionarea pompelor de protoni, modificarea pH-ului din apoplast din apropierea si extensia celulelor. Timpul de percepere a excitatiei gravitrope este de 12-30 s, iar timpul necesar pentru reactie este de 30-40 min. La plantele de Avena si continua 46-60 ore (Kaufman si Doyanandan, 1985).
Actiunea fortei de gravitatie determina cresterea radacinii spre sol si a tulpinii in sens ortogravitrop negativ. Ramificatiile de ordinul I ale tulpinii cresc plagiotrop, iar celelalte ramificatii sunt agravitrope si cresc in toate directiile.
Tulpinile aeriene prezinta un geotropism negativ, iar radacinile un geotropism pozitiv (fig. 182) Datorita geotropismului, la o planta asezata in pozitie verticala radacina se curbeaza in jos, iar tulpina in sus.
|
|
|
Fig.183 - Frunzele plantei de Coleus in pozitie normala (a) si pe clinostatul cu ax orizontal (b) (dupa Pfeffer) |
Prin anularea actiunii unilaterale a fortei de gravitatie, geotropismul nu se manifesta. Fenomenul poate fi pus in evidenta cu ajutorul unui aparat numit clinostat. Se aseaza o planta crescuta in ghiveci in pozitie orizontala si se roteste uniform in jurul axei cu ajutorul clinostatului, printr-o miscare lenta. Se constata ca planta nu mai produce curbarea geotropica a tulpinii, ci se alungeste in pozitie orizontala (fig. 183).
Pentru radacini concentratia auxinelor acumulate pe partea inferioara este supraoptimala, inhiband alungirea celulelor. Din aceasta cauza, celulele se alungesc mai intens pe partea superioara, determinand curbarea radacinii in miscarea de geotropism pozitiv.
Rolul geotropismului in viata plantelor este foarte important deoarece realizeaza orientarea organelor in mediul care le asigura desfasurarea normala a functiilor fiziologice, si anume absorbtia si fixarea la radacina si sustinerea organelor aeriene la tulpina
Miscarile gravitrope au o importanta deosebita pentru cultura plantelor, deoarece asigura inradacinarea acestora, precum si ridicarea culturilor de cereale culcate la pamant de furtuni sau de animale.
Fototropismul
Fototropismul este miscarea orientata de crestere provocata de actiunea unilaterala a luminii.Factorul de excitatie in acest caz este reprezentat de lumina, iar receptorii sunt reprezentati de pigmentii asimilatori situati in zona apicala. Plantele tinere iluminate unilateral se curbeaza in directia sursei de lumina. Curburile fototrofice au loc in zona elongatiei celulare.
Fototropismul se manifesta pozitiv la tulpina si negativ la radacina (fig.184).
Frunzele ca organe plagiotrope orienteaza limbul in asa fel incat radiatiile luminoase sa cada perpendicular pe acesta.
|
|
Fig. 184 - Orientarea organelor unei plantule crescuta cu radacina intr-o solutie nutritiva, fata de sursa de lumina |
Exista o stransa legatura intre cantitatea de lumina si reatia fototropica. Aceasta legatura este cunoscuta in literatura de specialitate ca legea Bunsen-Roscoe sau teorema reciprocitatii. Conform acestei teoreme exista o relatie indirecta intre intensitatea luminii si durata acesteia. Teorema presupunea ca efectele sunt aceleasi prin actiunea unei lumini de intensitate redusa pe perioada indelungata sau actiunea unei lumini intense pe durata scurta. Cercetari recente releva faptul ca teorema reciprocitatii este reala intr-un sens extrem de limitat.
Fototropismul este evident la plantele tinere, dar si la muscata din fereastra (Pelargonium zonale) care se orienteaza spre sursa de lumina. Ca si in cazul geotropismului, zona de percepere a excitatiei fototropice este varful organelor, iar zona de reactie este zona de extensie celulara
Mecanismul fototropismului este explicat tot prin participarea auxinelor. In conditii de iluminare uniforma, auxinele active in varful vegetativ migreaza bazipetal, uniform pe toate fetele organului, determinand alungirea uniforma a celulelor si cresterea verticala a acestuia.
Sub actiunea iluminarii unilaterale are loc modificarea concentratiei de auxine si anume acumularea acesteia pe fata neexpusa si scaderea pe fata expusa la lumina. Scaderea concentratiei de auxine sub actiunea luminii se poate explica fie prin inactivarea acestora si transformarea in lumi-auxin lactona, fie prin modificarea potentialului bioelectric celular, cu acumularea de sarcini electrice pozitive pe fata opusa sursei de lumina
Orientarea limbului frunzei in functie de sursa de lumina este asigurata de curbarea fototropica a petiolului. Aceasta orienteaza frunzele cu limbul perpendicular pe directia razelor de lumina (fig. 185).
Fenomenul de fototropism este deosebit de important in orientarea frunzelor spre o intensitate luminoasa optima, capabila sa realizeze intensitatea maxima a fotosintezei.
Hidrotropismul
Hidrotropismul este miscarea orientata de cresterea provocata de actiunea unilaterala a apei. Zona de percepere a excitatiei hidrotropice este varful radacinii, iar zona de reactie este zona neteda. Hidrotropismul explica cresterea sitemului radicular in functie de cantitatea de apa din sol. Se spune ca 'radacinile alearga in sol dupa sursa de apa'(fig. 186).
S-a demonstrat ca hidrotropismul poate anula geotropismul. In functie de cantitatea de apa din mediu, radacinile se pot curba geotropic negativ.
Hidrotropismul asigura manifestarea functiei radacinii de absorbtie a apei si elementelor minerale din sol.
Chimiotropismul
Chimiotropismul este miscarea orientata de crestere provocata de actiunea unilaterala a unui factor chimic. Chimiotropismul poate fi pozitiv si negativ. A fost demonstrat la radacini si in cresterea tubului polenic in stil.
Chimiotropismul pozitiv sau negativ al radacinii poate fi provocat de concentratia diferitelor solutii din sol. Concentratia diferita pe cele doua fete opuse ale radacinii poate provoca curbari pozitive sau negative, in functie de valorile acesteia. Se presupune ca diferenta de concentratie ar creea o diferenta de potential bioelectric, care sta la baza curbarii.
Chimiotropismul radacinii permite cresterea acesteia in solutii cu concentratie optimala, hipotonice si evita solutiile cu concentratie supraoptimala, hipertonice.
Chimiotropismul pozitiv al tubului polenic permite cresterea acestuia datorita unui factor chimic sintetizat in ovul, favorizeaza polenizarea si fecundarea cu polenul compatibil.
Tigmotropismul
Tigmotropismul este miscarea orientata de crestere provocata de actiunea unilaterala a substratului solid.
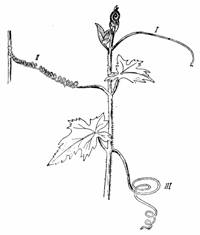
Tigmotropismul este foarte frecvent la carcei. Zona de percepere a excitatiei este fata inferioara si fetele laterale. Datorita tigmotropismului, carceii executa o miscarea de circumferinta, se rasucesc de suport si ajuta sustinerea tulpinilor agatatoare (fig. 187).
La Clematis si Tropaeolum tigmotropismul se manifesta la petiol, iar la Vanilla la radacinile adventive. La Bryonia dioica excitatia tigmotropica este preceputa de anumite celule epidermice cu peretii mai subtiri si cu o protoplasma ce contine diferite granule, bastonase si cristale. Acestea transmit excitatia la celulele fetei opuse si determina curbarea (fig. 188).
|
|
Fig. 188 - Carcei de Bryonia dioica: I - inainte de atingerea suportului; II - dupa atingerea suportului carcelul a format un resort intermediar intre suport si tulpina III - carcelul care nu a atins un suport ramane inchircit. |
NASTIILE
Nastiile sunt miscari neorientate de crestere provocate de actiunea neuniforma, cu intensitate variabila a unui factor de mediu.
Nastiile se realizeaza prin modificarea turgescentei celulelor pe doua fete opuse. Ele pot fi:
epinastii, cand turgescenta creste pe fata superioara, de exemplu la petale sau tepale, determinand deschiderea florilor;
hiponastii, cand turgescenta creste pe fata inferioara, de exemplu la petale sau tepale determinand inchiderea florilor.
Dupa natura factorului de mediu care actioneaza, nastiile pot fi: termonastii, fotonastii, nictinastii, tigmonastii si seismonastii.
Termonastiile
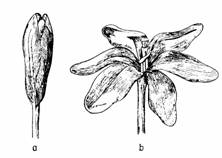
Termonastiile sunt provocate de intensitatea variabila a temperaturii. Sunt evidente la florile de lalele, Crocus etc. (fig. 189).
|
|
Fig. 189 - Floare de Crocus luteus: a - la temperatura joasa in stare inchisa b - la temperatura ridicata, in stare deschisa |
La lalele, florile inchise aduse la temperatura camerei se deschid. (fig.190) La Crocus, florile prezinta sensibilitate la o variatie de temepratura de 00- 50C.
Daca plantele sunt tinute sub o anumita temperatura critica, florile nu se deschid niciodata. Prin aceasta metoda, la lalele poate fi indusa cleistogamia respectiv fecundarea in interiorul florii inchise.
Fotonastiile
Fotonastiile sunt provocate de intensitatea variabila a luminii. Sunt miscari caracteristice inflorescentelor speciilor compozite, cat si an cazul foliolelor unor specii ca: Phaseolus, Albizzea, Mumosa, Robinia, Oxalis, Trofolium, etc. (fig. 191 ).
Miscarea este determinata de modificarea gradului de hidratare a celulelor situate de o parte si de alta a pulvinulelor, ca urmare a modificarii presiunii osmotice a sucului vacuolar.
Ele sunt evidente la Gentiana nivalis, care isi inchide florile la simpla trecere a unui nor prin fata soarelui.
Nictinastiile
Nictinastiile sunt provocate sunt provocate de alternanta zilei cu noaptea. Ele sunt evidente la frunzele de Stellaria , care in timpul noptii se ridica si se apropie de tulpina si de Impatiens parviflora si Polygonum convolvulus, care se apleaca in jos. La lumina sau intuneric continuu, nictinastiile isi pierd din intensitate. (fig. 192).
Tigmonastiile
Tigmonastiile sunt miscarile unor organe ale plantelor carnivore provocate de atingerea receptorilor. Stimularea perilor senzitivi de pe frunzele de Drosera sau Dionaea de catre insecte determina miscari ale perilor sau ale frunzelor care duc la capturarea insectei. Mecanismele de realizare a miscarii sunt extrem de complexe, dar se bazeaza an principal pe moficarea turgescentei celulelor perilor senzitivi sau a unor pulvinule situate la baza limbului foliar (fig. 193).
Seismonastiile
Seismonastiile sunt provocate de actiunea unui factor mecanic. Exemplul cel mai evident este Mimosa pudica, la care atingerea frunzelor este urmata de lasarea acestora in jos. Dupa 1/4 - 1/2 ora apare miscarea de revenire. Se presupune ca seismonastiile sunt provocate de modificarea turgescentei unor celule de la baza petiolului, numite pulvinule (fig. 194).
Aceste modificari de turgescenta apar datorita cresterii permeabilitatii plasmalemei si tonoplastului, intensificarii activitatii pompelor de ioni si cresterii rapide a concentratiei ionilor de potasiu din apoplast. Ionii de potasiu din celulele de la baza pulvinulelor sunt transpotati pe cale apoplasmica in celulele de la partea superioara a acestora, unde determina cresterea presiunii osmotice, absorbtia apei si cresterea turgescentei. Drept urmare frunza cfompusa realizeaza o miscare de coborare, iar foliolele se apropie. Dupa un anumit interval de timp frunzele si foliolele revin la pozitia initiala
Un alt exemplu il constituie filamentele senzitive ale staminelor. La Berberis vulgaris filamentele staminelor se curbeaza la atingerea unei insecte, favorizand polenizarea entomofila. La Centaurea nigra si C. montana sensibilitatea filamentului la atingerea mecanica favorizeaza eliminarea polenului din antere si de asemenea polenizarea entomofila. La Cobea scandens filamentele staminelor se retrag la nivelul stigmatului in momentul caderii corolei, favorizand autopolenizarea (fig. 195).
NUTATIILE
Nutatiile reprezinta miscarile laterale sau in jurul unei axe pe care le efectueaza plantele fixate sub influenta unor factori interni. Exista nutatii efemere si periodice.
Nutatiile efemere se produc o singura data in viata plantelor si sunt reprezentate de miscarile pe care le efectueaza cotiledoanele in timpul germinarii semintelor, pana ajung la pozitia orizontala
Nutatiile periodice sunt specifice plantelor volubile si reprezinta miscari circulare pe care le efectueaza tulpina in timpul cresterii. Durata unei miscari variaza in functie de specie, intre cateva minute si cateva ore.
Nutatiile se datoresc vitezei diferite de extensie a celulelor din peretii laterali ai tulpinii. Aceasta diferenta a ritmului de crestere se presupune ca se datoreste transportului auxinei sub forma de unde cu concentratii diferite.
La unele specii (Desmodium gyrans), miscarile de circumnutatie a foliolelor sunt cauzate de variatia turgescentei pulvinulelor de la baza foliolelor, ca urmare a transportului apei si, probabil, al ionilor.
APLICATII PRACTICE DE LABORATOR
TROPISME
Tropismele sunt mi[cri de cre[tere a plantelor determinate de ac]iunea unilateral a unui factor excitant extern. Tropismele pot fi pozitive cand organul `n cre[tere se orienteaz `n direc]ia de ac]iune a excitantului, [i negative cand organul `n cre[tere se orienteaz opus direc]iei excitantului.
Dup natura factorului extern care ac]ioneaz ca excitant al mi[crii, tropismele se clasific `n: geotropism, fototropism, hidrotropism [i chimiotropism.
Geotropismul
Este mi[carea de cre[tere a plantelor provocat de ac]iunea unilateral a for]ei de gravita]ie. Geotropismul este pozitiv `n cazul rdcinii [i negativ la tulpin.
Punerea `n eviden] a geotropismului pozitiv la rdcini
Materiale necesare plantule de mazre sau bob, camer umed, plac de sticl `nvelit `n sugativ, lam de ras.
Modul de lucru Se folose[te o plac de sticl `nvelit `n sugativ pe care se a[eaz dou plantule de mazre cu rdcinile `n pozi]ie orizontal. Una dintre plante rmane intact iar la a doua se `ndeprteaz varful vegetativ. Se introduce placa `ntr-o camer umed. Dup 24 de ore se constat c la plantula intact, rdcina a crescut [i s-a curbat `n jos pe cand la plantula la care s-a `ndeprtat varful rdcinii (piloriza [i zona neted), rdcina nu a crescut [i nu s-a curbat.
Interpretare. Sub ac]iunea gravita]iei, rdcina a reac]ionat manifestand un geotropism pozitiv. Varful vegetativ al rdcinii reprezint zona de percepere a excita]iei geotropice iar zona neted reprezint zona de reac]ie.
Durata experien]ei observare 10 minute la interval de 24 ore.
Punerea `n eviden] a geotropismului negativ la tulpin
Materiale necesare ghivece cu plante de mazre, stativ metalic cu cleme.
Modul de lucru Se a[eaz ghivecele cu plantule de mazre `n pozi]ie orizontal. Dup 24 ore se constat c tulpinile plantelor se curbeaz `n sus.
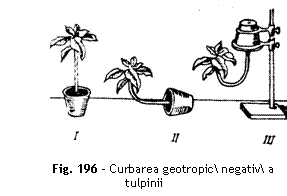 Interpretare Sub ac]iunea gravita]iei,
tulpina se curbeaz `n sus, manifestand un geotropism negativ (fig.196 ). Mecanismul geotropismului
este explicat prin participarea auxinelor. ~n pozi]ie vertical, poten]ialul
bioelectric al organelor este situat cu sarcinile negative `n varful organelor
[i cu sarcinile pozitive la baza acestora, ceea ce determin o alungire
uniform a celulelor pe toate fe]ele organelor, rezultand cre[terea vertical.
La plantele situate `n pozi]ie orizontal se modific poten]ialul bioelectric
al organelor, sarcinile negative se situeaz pe fa]a superioar iar cele
pozitive pe fa]a inferioar a tulpinii [i a rdcinii. Doza de auxine acumulat
este stimulatoare pentru alungirea celulelor din fa]a inferioar a tulpinii [i
determin curbarea geotropic negativ. Pentru rdcini, concentra]ia auxinelor
pe fa]a inferioar este supraoptimal [i inhib alungirea celulelor din fa]a
inferioar a rdcinii; `n schimb, celulele de pe fa]a superioar cresc mai
intens, rezultand curbarea geotropic pozitiv. Geotropismul joac un rol
deosebit `n realizarea func]iilor rdcinii [i tulpinii.
Interpretare Sub ac]iunea gravita]iei,
tulpina se curbeaz `n sus, manifestand un geotropism negativ (fig.196 ). Mecanismul geotropismului
este explicat prin participarea auxinelor. ~n pozi]ie vertical, poten]ialul
bioelectric al organelor este situat cu sarcinile negative `n varful organelor
[i cu sarcinile pozitive la baza acestora, ceea ce determin o alungire
uniform a celulelor pe toate fe]ele organelor, rezultand cre[terea vertical.
La plantele situate `n pozi]ie orizontal se modific poten]ialul bioelectric
al organelor, sarcinile negative se situeaz pe fa]a superioar iar cele
pozitive pe fa]a inferioar a tulpinii [i a rdcinii. Doza de auxine acumulat
este stimulatoare pentru alungirea celulelor din fa]a inferioar a tulpinii [i
determin curbarea geotropic negativ. Pentru rdcini, concentra]ia auxinelor
pe fa]a inferioar este supraoptimal [i inhib alungirea celulelor din fa]a
inferioar a rdcinii; `n schimb, celulele de pe fa]a superioar cresc mai
intens, rezultand curbarea geotropic pozitiv. Geotropismul joac un rol
deosebit `n realizarea func]iilor rdcinii [i tulpinii.
Durata experien]ei observare 5 minute la interval de 24 ore.
Anularea efectului gravita]iei la plante prin utilizarea clinostatului
Pricipiu Organele sensibile la ac]iunea gravita]iei (tulpini, rdcini), ]inute `n pozi]ie orizontal [i supuse unei rota]ii lente (1-10 ture/or) `n jurul axei lor, nu mai prezint curbur geotropic.
Materiale necesare ghivece cu plante de mazre sau Coleus.
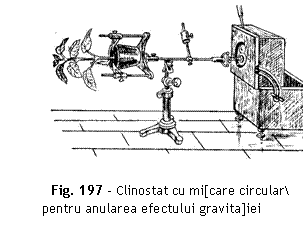 Modul de lucru Ghiveciul se
fixeaz `n pozi]ie orizontal pe axul orizontal al clinostatului, cruia i se
imprim o mi[care lent (fig. 197).
Un alt ghiveci cu aceia[i specie de plant se a[eaz alturi `n pozi]ie
orizontal, dar se men]ine `n repaus. Dup cateva ore se observ c la planta
control, aflat `n repaus, tulpina se curbeaz geotropic negativ, `n timp ce la
planta de pe clinostat, tulpina continu s creasc orizontal, fr reac]ie
geotropic.
Modul de lucru Ghiveciul se
fixeaz `n pozi]ie orizontal pe axul orizontal al clinostatului, cruia i se
imprim o mi[care lent (fig. 197).
Un alt ghiveci cu aceia[i specie de plant se a[eaz alturi `n pozi]ie
orizontal, dar se men]ine `n repaus. Dup cateva ore se observ c la planta
control, aflat `n repaus, tulpina se curbeaz geotropic negativ, `n timp ce la
planta de pe clinostat, tulpina continu s creasc orizontal, fr reac]ie
geotropic.
Interpretare Prin rota]ii lente, toate fe]ele tulpinii sau rdcinii sunt excitate succesiv de for]a gravita]ional, excita]ia nu se mai exercit unilateral, efectele se anuleaz. Dup cateva ore se constat c la planta control aflat `n repaus, tulpina se curbeaz `n sus, manifestand curbura geotropic negativ, `n timp ce la planta de pe clinostat, tulpina continu s creasc orizontal, fr reac]ie geotropic. Efectele se anuleaz reciproc iar organul vegetal continu s creasc orizontal.
Durata experientei observare 5 minute la interval de 24 ore.
Fototropismul
Este mi[carea de orientare a plantelor, determinat de ac]iunea unilateral a luminii. Dac plantele tinere, cu ]esuturi meristematice active sunt luminate unilateral, are loc curbura tulpinilor spre sursa de lumin, manifestand un fototropism pozitiv. Fa] de luminarea unilateral, rdcinile manifest un fototropism negativ iar frunzele, fiind plagiotrope, `[i orienteaz limbul spre sursa luminoas, astfel ca razele de lumin s cad perpendicular pe fa]a superioar (diatropism), ca o consecin] a ortofototropismului pozitiv al pe]iolului.
Punerea `n eviden] a fototropismului pozitiv al tulpinii
Materiale necesare camer fototropic cu pere]i intermediari cptu[i]i cu hartie neagr avand un orificiu pe un perete lateral, ghivece cu plantule de grau sau ovz, ghivece cu plante de mu[cat (Pelargonium).
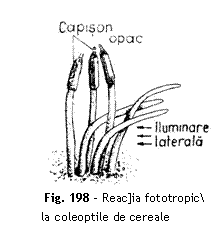 Modul de lucru ~n camera
fototropic se introduc ghivece cu plantule de grau sau ovz, `n faz de
coleoptil, [i ghivece cu plante de mu[cat. La o parte din coleoptile se
acoper varful cu un capi[on de staniol, iar la alte plantule se las
neacoperite. Timp de 24 de ore plantele primesc o iluminare unilateral, razele
de lumin ptrunzand prin orificiul aflat `n peretele camerei fotrotropice. La
grau [i ovz se constat curbarea spre sursa de lumin a coleoptilelor rmase
neacoperite, `n timp ce coleoptilele cu varful acoperit au crescut vertical [i
nu au reac]ionat la excitantul luminos lateral (fig. 198). La plantele de
mu[cat se observ curbarea spre sursa de lumin atat a varfului tulpinii cat
[i a frunzelor, orientate cu limbul perpendicular pe fasciculul luminos.
Modul de lucru ~n camera
fototropic se introduc ghivece cu plantule de grau sau ovz, `n faz de
coleoptil, [i ghivece cu plante de mu[cat. La o parte din coleoptile se
acoper varful cu un capi[on de staniol, iar la alte plantule se las
neacoperite. Timp de 24 de ore plantele primesc o iluminare unilateral, razele
de lumin ptrunzand prin orificiul aflat `n peretele camerei fotrotropice. La
grau [i ovz se constat curbarea spre sursa de lumin a coleoptilelor rmase
neacoperite, `n timp ce coleoptilele cu varful acoperit au crescut vertical [i
nu au reac]ionat la excitantul luminos lateral (fig. 198). La plantele de
mu[cat se observ curbarea spre sursa de lumin atat a varfului tulpinii cat
[i a frunzelor, orientate cu limbul perpendicular pe fasciculul luminos.
Interpretare Atat tulpinile cat [i frunzele manifest un fototropism pozitiv. Experien]a cu coleoptile de cereale, demonstreaz c perceperea excita]iei luminoase are loc `n varf, iar reac]ia de curbare este imediat sub varf, ceea ce corespunde cu zona de activitate a meristemelor de cre[tere.
Mecanismul fototropismului este corelat cu starea de activitate a auxinelor naturale din zona de cre[tere a tulpinii [i cu cre[terea inegal a celulelor de pe cele dou fe]e ale tulpinii, unde este inhibat cre[terea celulelor.
Pe fa]a opus a tulpinii, aflat `n umbr [i semiumbr, nu are loc degradarea auxinelor iar celulele prezint o cre[tere normal prin extensie.
Durata experien]e: observare 15 minute la interval de 24 ore.
Hidrotropismul
Este mi[carea orientat de cre[tere a organelor plantelor provocat de repartizarea neuniform a umidit]ii `n mediul extern. Rdcinile plantelor prezint un hidrotropism accentuat avand zona de percepere a diferen]ei de umiditate situat `n varf, iar zona de reac]ie `n subzona auxezis caracteristic prin alungirea celulelor.
Punerea `n eviden] a hidrotropismului pozitiv al rdcinilor
Materiale necesare dou plcu]e de sticl, dou pahare Berzelius, hartie sugativ, semin]e de in sau mu[tar.
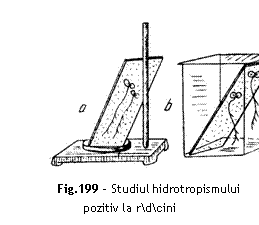 Modul de lucru Se folosesc dou
plcu]e de sticl `nvelite `n sugativ, pe care se a[eaz semin]e de in sau de
mu[tar. Plcu]ele cu semin]e pe fa]a inferioar, sunt introduse oblic `n cate o
camer umed (pahar Berzelius cu strat de ap de 1-2 cm). Una din camere este
acoperit iar cealalt este descoperit. Dup 48 ore se constat c `n camera
acoperit, rdcinile plantelor cresc vertical iar `n camera descoperit ader
la plcu]e, lipindu-se de sugativa umed (fig. 199).
Modul de lucru Se folosesc dou
plcu]e de sticl `nvelite `n sugativ, pe care se a[eaz semin]e de in sau de
mu[tar. Plcu]ele cu semin]e pe fa]a inferioar, sunt introduse oblic `n cate o
camer umed (pahar Berzelius cu strat de ap de 1-2 cm). Una din camere este
acoperit iar cealalt este descoperit. Dup 48 ore se constat c `n camera
acoperit, rdcinile plantelor cresc vertical iar `n camera descoperit ader
la plcu]e, lipindu-se de sugativa umed (fig. 199).
Interpretare ~n camera acoperit se creaz o umiditate ridicat [i uniform [i de aceea factorul ap ac]ioneaz cu egal intensitate din toate direc]iile. Rdcinile cresc perfect vertical dup o reac]ie pozitiv geotropic. ~n camera descoperit, umiditatea este neuniform iar rdcina manifest un hidrotropism pozitiv orientandu-se spre hartia sugativ `mbibat cu ap.
~n sol, rdcinile se orienteaz spre sursa de ap [i de aceea sistemul radicular este inegal repartizat, `n dependen] de varia]iile de umiditate ale solului.
Durata experien]ei 10 minute dup 48 ore de la montare.
Chimiotropismul
Este o mi[care de orientare a organelor plantelor provocat de concentra]ia inegal a unor substan]e din mediul extern. Factorul chimic ac]ioneaz unilateral determinand o reac]ie pozitiv la doze optime [i o reac]ie negativ la doze toxice.
Observarea curburii chimiotrope a rdcinilor
Principiu. Reac]ia pozitiv sau negativ chimiotrop depinde atat de concentra]ia substan]elor cat [i de natura lor chimic.
Materiale necesare cristalizor sau vas Petri, solu]ie 10% de gelatin, eprubet, solu]ie 2% de KNO3, solu]ie 2% de NaCl, plantule de mazre cu rdcini drepte.
Modul de lucru ~ntr-un cristalizor mic sau un vas Petri se toarn o plac de gelatin `n care se fac 3 excava]ii cu ajutorul unei eprubete `nclzite. ~n prima excava]ie se toarn ap, a doua se umple cu solu]ie 2% de KNO3, iar `n a 3-a se pune solu]ie 2% de NaCl. ~n fiecare din cele 3 solu]ii se introduc rdcinile a cate unei plantule de mazre [i se las `n repaus, la `ntuneric. Se observ dup 24 ore c rdcinile au rmas scufundate `n ap [i `n solu]ia de KNO3 continuand procesele de cre[tere, `n timp ce la a 3-a excava]ie rdcinile ies din solu]ia de NaCl, se orienteaz `ntr-o direc]ie opus [i `nceteaz procesele de cre[tere.
Interpretare. Rdcinile manifest un chimitropism pozitiv pentru ap [i solu]ia diluat de KNO3 [i un chimiotropism negativ pentru solu]ia de NaCl
Experien]a poate fi repetat [i cu alte solu]ii de zaharoz [i fosfat acid de amoniu pentru chimiotropism pozitiv, acizi minerali [i sulfat de Mg pentru chimiotropism negativ.
Un exemplu de chimiotropism pozitiv este orientarea grunciorilor de polen `n stil spre ovar.
Durata experien]ei 10 minute dup 24 ore de la montare.
NASTIILE
Sunt mi[cri neorientate ale organelor plantelor, provocate de varia]ia de intensitate a unor factori excitan]i, fizici sau chimici. Dup natura excitantului se deosebesc mai multe tipuri de nastii: fotonastii, termonastii, seismonastii, nictinastii.
Fotonastiile
Sunt mi[cri nastice provocate de varia]ia intensit]ii luminii, independente de direc]ia fasciculului de lumin.
Observarea mi[crilor fotonastice
Materiale necesare ghivece cu plante de Bellis perennis [i Taraxacum officinalis.
Modul de lucru Plantele florifere de Bellis perennis sau Taraxacum officinalis, se ]in la lumin intens [i se constat deschiderea florilor. Trecute la `ntuneric, florile se `nchid `ntr-un interval de timp relativ scurt.
Interpretare. Datorit varia]iei `n intensitate a luminii are loc o cre[tere inegal a celulelor pe cele dou fe]e ale `nveli[urilor florale.
Durata experien]ei 30 minute (observare).
Nictinastiile
Sunt mi[cri nastice prin care se modific pozi]ia frunzelor `n func]ie de alternan]a zi-noapte. ~n natur, fenomenul de nictinastie este cunoscut sub denumirea de 'somn' al plantelor.
Observarea nictinastiilor la plante
Principiu Prin modificarea turgescen]ei la pulvinulele de la baza pe]iolului are loc declan[area lent a mi[crii nictinastice la trecerea de la zi la noapte.
Materiale necesare plante leguminoase (fasole, salcam).
Modul de lucru Speciile de plante leguminoase (fasole, salcam) se urmresc `n orele de sear [i se constat c paralel cu reducerea intensit]ii luminii la `nserare, foliolele frunzelor se a[eaz fa] `n fa], iar axul frunzelor se `nclin `n jos.
Interpretare. Prin reducerea intensit]ii luminii la trecerea de la zi la noapte scade treptat turgescen]a ]esuturilor din pulvinulele de la baza foliolelor [i a pe]iolului, determinand mi[carea nictinastic.
Durata experien]ei: 1 or seara.
Seismonastiile.
Sunt mi[cri neorientate, provocate de excita]ii mecanice externe care determin modificri `n turgescen]a celulelor.
Provocarea seismonastiei la Mimosa pudica
Materiale necesare ghivece cu plante de Mimosa pudica.
 Modul de lucru Prin atingerea
frunzelor de Mimosa cu un obiect tare, se constat c imediat foliolele frunzei
se a[eaz fa] `n fa], iar frunza `ntreag se apleac `n jos (fig. 200).
Modul de lucru Prin atingerea
frunzelor de Mimosa cu un obiect tare, se constat c imediat foliolele frunzei
se a[eaz fa] `n fa], iar frunza `ntreag se apleac `n jos (fig. 200).
Interpretare. {ocul mecanic determin modificarea turgescen]ei ]esuturilor crnoase de la baza foliolelor [i de la baza frunzei. Excita]ia perceput la nivelul foliolelor se transmite de la celul la celul, prin intermediul plasmodesmelor [i provoac o reac]ie la baza pe]iolului, ceea ce determin aplecarea `n jos a `ntregii frunze. ~n vehicularea excita]iei contribuie acidul traumatic produs de celulele atinse.
Mi[cri seismonastice se constat de asemenea la filamentul staminal al florii de Berberis vulgaris, la frunzele de Oxalis acetosella [i la inflorescen]ele de Bellis perennis.
Durata experien]ei: 10 minute
Termonastiile
Sunt mi[cri neorientate de cre[tere datorate intensit]ii variabile a temperaturii.
Observarea mi[crilor termonastice la lalele
Materiale necesare flori de Tulipa gesneriana (lalea), termometru, 2 flacoane Erlenmeyer cu ap rece [i cald.
Modul de lucru Prin mutarea florilor de lalea de la temperatur sczut (80C) la temperatur ridicat (250C) se constat deschiderea rapid a tepalelor.
Interpretare. Mi[carea termonastic este provocat de o cre[tere inegal a fe]elor opuse ale tepalelor la flori. La temperaturi sczute cresc mai intens celulele de pe fa]a extern a tepalelor, iar la temperaturi optime (25-300C) cresc mai intens celulele de pe fa]a intern a tepalelor, ceea ce declan[eaz deschiderea florilor.
Durata experien]ei 15 minute.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 8468
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved