| CATEGORII DOCUMENTE |
| Bulgara | Ceha slovaca | Croata | Engleza | Estona | Finlandeza | Franceza |
| Germana | Italiana | Letona | Lituaniana | Maghiara | Olandeza | Poloneza |
| Sarba | Slovena | Spaniola | Suedeza | Turca | Ucraineana |
|
The Cell and Its Functions |
|
|
Each of the 100 trillion cells in a human being is a living structure that can
survive for months or many years, provided its surrounding fluids contain
appropriate nutrients. To understand the function of organs and other
structures of the body, it is essential that we first understand the basic
organization of the cell and the functions of its component parts.
|
Organization of the Cell |
|
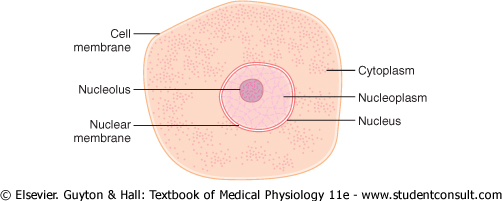
A typical cell, as seen by the light microscope, is shown in Figure 2-1. Its two major parts are the nucleus and the cytoplasm. The nucleus is separated from the cytoplasm by a nuclear membrane, and the cytoplasm is separated from the surrounding fluids by a cell membrane, also called the plasma membrane. |
|
The different substances that make up the cell are collectively called protoplasm. Protoplasm is composed mainly of five basic substances: water, electrolytes, proteins, lipids, and carbohydrates. |
|
Water. The principal fluid medium of the cell is water, which is present in most cells, except for fat cells, in a concentration of 70 to 85 per cent. Many cellular chemicals are dissolved in the water. Others are suspended in the water as solid particulates. Chemical reactions take place among the dissolved chemicals or at the surfaces of the suspended particles or membranes. |
|
Ions. The most important ions in the cell are potassium, magnesium, phosphate, sulfate, bicarbonate, and smaller quantities of sodium, chloride, and calcium. These are all discussed in more detail in Chapter 4, which considers the interrelations between the intracellular and extracellular fluids. |
|
The ions provide inorganic chemicals for cellular reactions. Also, they are necessary for operation of some of the cellular control mechanisms. For instance, ions acting at the cell membrane are required for transmission of electrochemical impulses in nerve and muscle fibers. |
|
Proteins. After water, the most abundant substances in most cells are proteins, which normally constitute 10 to 20 per cent of the cell mass. These can be divided into two types: structural proteins and functional proteins. |
|
Structural proteins are present in the cell mainly in the form of long filaments that themselves are polymers of many individual protein molecules. A prominent use of such intracellular filaments is to form microtubules that provide the 'cytoskeletons' of such cellular organelles as cilia, nerve axons, the mitotic spindles of mitosing cells, and a tangled mass of thin filamentous tubules that hold the parts of the cytoplasm and nucleoplasm together in their respective compartments. Extracellularly, fibrillar proteins are found especially in the collagen and elastin fibers of connective tissue and in blood vessel walls, tendons, ligaments, and so forth. |
|
page 11 |
|
|
|
page 12 |
|
|
|
Figure 2-1 Structure of the cell as seen with the light microscope. |
|
The functional proteins are an entirely
different type of protein, usually composed of combinations of a few
molecules in tubular-globular form. These proteins are mainly the enzymes
of the cell and, in contrast to the fibrillar proteins, are often mobile in
the cell fluid. Also, many of them are adherent to membranous structures
inside the cell. The enzymes come into direct contact with other substances
in the cell fluid and thereby catalyze specific intracellular chemical
reactions. For instance, the chemical reactions that split glucose |
|
Lipids. Lipids are several types of substances that are grouped together because of their common property of being soluble in fat solvents. Especially important lipids are phospholipids and cholesterol, which together constitute only about 2 per cent of the total cell mass. The significance of phospholipids and cholesterol is that they are mainly insoluble in water and, therefore, are used to form the cell membrane and intracellular membrane barriers that separate the different cell compartments. |
|
In addition to phospholipids and cholesterol, some cells contain large quantities of triglycerides, also called neutral fat. In the fat cells, triglycerides often account for as much as 95 per cent of the cell mass. The fat stored in these cells represents the body's main storehouse of energy-giving nutrients that can later be dissoluted and used to provide energy wherever in the body it is needed. |
|
Carbohydrates. Carbohydrates have
little structural function in the cell except as parts of glycoprotein
molecules, but they play a major role in nutrition of the cell. Most human
cells do not maintain large stores of carbohydrates; the amount usually
averages about 1 per cent of their total mass but increases to as much as 3
per cent in muscle cells and, occasionally, 6 per cent in liver cells.
However, carbohydrate in the form of dissolved glucose |
|
Physical Structure of the Cell |
|
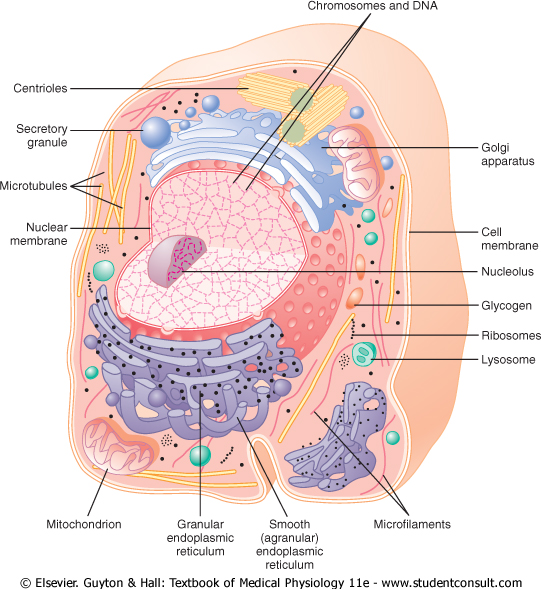
The cell is not merely a bag of fluid, enzymes, and chemicals; it also contains highly organized physical structures, called intracellular organelles. The physical nature of each organelle is as important as the cell's chemical constituents for cell function. For instance, without one of the organelles, the mitochondria, more than 95 per cent of the cell's energy release from nutrients would cease immediately. The most important organelles and other structures of the cell are shown in Figure 2-2. |
|
Membranous Structures of the Cell |
|
Most organelles of the cell are covered by membranes composed primarily of lipids and proteins. These membranes include the cell membrane, nuclear membrane, membrane of the endoplasmic reticulum, and membranes of the mitochondria, lysosomes, and Golgi apparatus. |
|
The lipids of the membranes provide a barrier that impedes the movement of water and water-soluble substances from one cell compartment to another because water is not soluble in lipids. However, protein molecules in the membrane often do penetrate all the way through the membrane, thus providing specialized pathways, often organized into actual pores, for passage of specific substances through the membrane. Also, many other membrane proteins are enzymes that catalyze a multitude of different chemical reactions, discussed here and in subsequent chapters. |
|
Cell Membrane |
|
The cell membrane (also called the plasma membrane), which envelops the cell, is a thin, pliable, elastic structure only 7.5 to 10 nanometers thick. It is composed almost entirely of proteins and lipids. The approximate composition is proteins, 55 per cent; phospholipids, 25 per cent; cholesterol, 13 per cent; other lipids, 4 per cent; and carbohydrates, 3 per cent. |
|
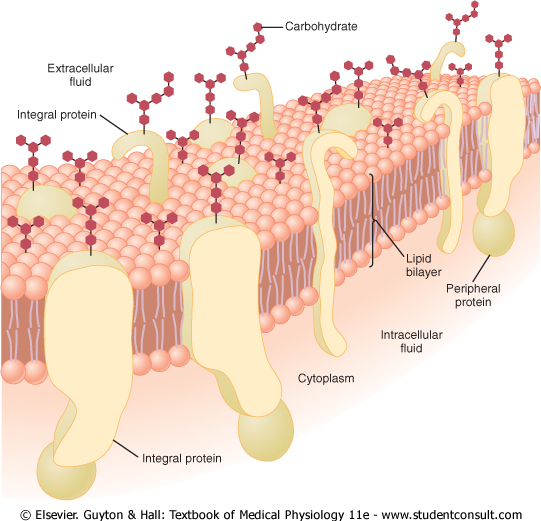
Lipid Barrier of the Cell Membrane Impedes Water Penetration. Figure 2-3 shows the structure of the cell membrane. Its basic structure is a lipid bilayer, which is a thin, double-layered film of lipids-each layer only one molecule thick-that is continuous over the entire cell surface. Interspersed in this lipid film are large globular protein molecules. |
|
page 12 |
|
|
|
page 13 |
|
||
|
|
|
Figure 2-2 Reconstruction of a typical cell, showing the internal organelles in the cytoplasm and in the nucleus. |
|
The basic lipid bilayer is composed of phospholipid molecules. One end of each phospholipid molecule is soluble in water; that is, it is hydrophilic. The other end is soluble only in fats; that is, it is hydrophobic. The phosphate end of the phospholipid is hydrophilic, and the fatty acid portion is hydrophobic. |
|
Because the hydrophobic portions of the phospholipid molecules are repelled by water but are mutually attracted to one another, they have a natural tendency to attach to one another in the middle of the membrane, as shown in Figure 2-3. The hydrophilic phosphate portions then constitute the two surfaces of the complete cell membrane, in contact with intracellular water on the inside of the membrane and extracellular water on the outside surface. |
|
The lipid layer in the middle of the membrane
is impermeable to the usual water-soluble substances, such as ions, glucose |
|
The cholesterol molecules in the membrane are also lipid in nature because their steroid nucleus is highly fat soluble. These molecules, in a sense, are dissolved in the bilayer of the membrane. They mainly help determine the degree of permeability (or impermeability) of the bilayer to water-soluble constituents of body fluids. Cholesterol controls much of the fluidity of the membrane as well. |
|
Cell Membrane Proteins. Figure 2-3 also shows globular masses floating in the lipid bilayer. These are membrane proteins, most of which are glycoproteins. Two types of proteins occur: integral proteins that protrude all the way through the membrane, and peripheral proteins that are attached only to one surface of the membrane and do not penetrate all the way through. |
|
Many of the integral proteins provide structural channels (or pores) through which water molecules and water-soluble substances, especially ions, can diffuse between the extracellular and intracellular fluids. These protein channels also have selective properties that allow preferential diffusion of some substances over others. |
|
Other integral proteins act as carrier proteins for transporting substances that otherwise could not penetrate the lipid bilayer. Sometimes these even transport substances in the direction opposite to their natural direction of diffusion, which is called 'active transport.' Still others act as enzymes. |
|
page 13 |
|
|
|
page 14 |
|
|||
|
|
|
Figure 2-3 Structure of the cell membrane, showing that it is composed mainly of a lipid bilayer of phospholipid molecules, but with large numbers of protein molecules protruding through the layer. Also, carbohydrate moieties are attached to the protein molecules on the outside of the membrane and to additional protein molecules on the inside. (Redrawn from Lodish HF, Rothman JE: The assembly of cell membranes. Sci Am 240:48, 1979. Copyright George V. Kevin.) |
|
Integral membrane proteins can also serve as receptors for water-soluble chemicals, such as peptide hormones, that do not easily penetrate the cell membrane. Interaction of cell membrane receptors with specific ligands that bind to the receptor causes conformational changes in the receptor protein. This, in turn, enzymatically activates the intracellular part of the protein or induces interactions between the receptor and proteins in the cytoplasm that act as second messengers, thereby relaying the signal from the extracellular part of the receptor to the interior of the cell. In this way, integral proteins spanning the cell membrane provide a means of conveying information about the environment to the cell interior. |
|
Peripheral protein molecules are often attached to the integral proteins. These peripheral proteins function almost entirely as enzymes or as controllers of transport of substances through the cell membrane 'pores.' |
|
Membrane Carbohydrates-The Cell 'Glycocalyx.' Membrane carbohydrates occur almost invariably in combination with proteins or lipids in the form of glycoproteins or glycolipids. In fact, most of the integral proteins are glycoproteins, and about one tenth of the membrane lipid molecules are glycolipids. The 'glyco' portions of these molecules almost invariably protrude to the outside of the cell, dangling outward from the cell surface. Many other carbohydrate compounds, called proteoglycans-which are mainly carbohydrate substances bound to small protein cores-are loosely attached to the outer surface of the cell as well. Thus, the entire outside surface of the cell often has a loose carbohydrate coat called the glycocalyx. |
|
The carbohydrate moieties attached to the outer surface of the cell have several important functions: (1) Many of them have a negative electrical charge, which gives most cells an overall negative surface charge that repels other negative objects. (2) The glycocalyx of some cells attaches to the glycocalyx of other cells, thus attaching cells to one another. (3) Many of the carbohydrates act as receptor substances for binding hormones, such as insulin; when bound, this combination activates attached internal proteins that, in turn, activate a cascade of intracellular enzymes. (4) Some carbohydrate moieties enter into immune reactions, as discussed in Chapter 34. |
|
Cytoplasm and Its Organelles |
|
The cytoplasm is filled with both minute and
large dispersed particles and organelles. The clear fluid portion of the
cytoplasm in which the particles are dispersed is called cytosol; this
contains mainly dissolved proteins, electrolytes, and glucose |
|
page 14 |
|
|
|
page 15 |
|
Dispersed in the cytoplasm are neutral fat globules, glycogen granules, ribosomes, secretory vesicles, and five especially important organelles: the endoplasmic reticulum, the Golgi apparatus, mitochondria, lysosomes, and peroxisomes. |
|
Endoplasmic Reticulum |
|
Figure 2-2 shows a network of tubular and flat vesicular structures in the cytoplasm; this is the endoplasmic reticulum. The tubules and vesicles interconnect with one another. Also, their walls are constructed of lipid bilayer membranes that contain large amounts of proteins, similar to the cell membrane. The total surface area of this structure in some cells-the liver cells, for instance-can be as much as 30 to 40 times the cell membrane area. |
|
The detailed structure of a small portion of endoplasmic reticulum is shown in Figure 2-4. The space inside the tubules and vesicles is filled with endoplasmic matrix, a watery medium that is different from the fluid in the cytosol outside the endoplasmic reticulum. Electron micrographs show that the space inside the endoplasmic reticulum is connected with the space between the two membrane surfaces of the nuclear membrane. |
|
Substances formed in some parts of the cell enter the space of the endoplasmic reticulum and are then conducted to other parts of the cell. Also, the vast surface area of this reticulum and the multiple enzyme systems attached to its membranes provide machinery for a major share of the metabolic functions of the cell. |
|
|||
|
|
|
Figure 2-4 Structure of the endoplasmic reticulum. (Modified from DeRobertis EDP, Saez FA, DeRobertis EMF: Cell Biology, 6th ed. Philadelphia: WB Saunders, 1975.) |
|
Ribosomes and the Granular Endoplasmic Reticulum. Attached to the outer surfaces of many parts of the endoplasmic reticulum are large numbers of minute granular particles called ribosomes. Where these are present, the reticulum is called the granular endoplasmic reticulum. The ribosomes are composed of a mixture of RNA and proteins, and they function to synthesize new protein molecules in the cell, as discussed later in this chapter and in Chapter 3. |
|
Agranular Endoplasmic Reticulum. Part of the endoplasmic reticulum has no attached ribosomes. This part is called the agranular, or smooth, endoplasmic reticulum. The agranular reticulum functions for the synthesis of lipid substances and for other processes of the cells promoted by intrareticular enzymes. |
|
Golgi Apparatus |
|
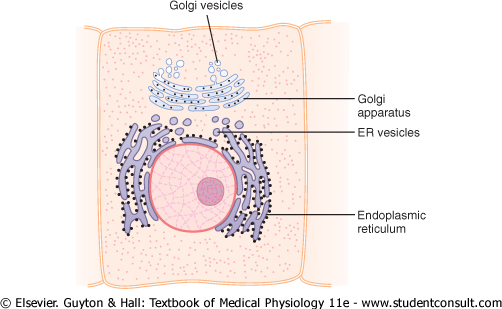
The Golgi apparatus, shown in Figure 2-5, is closely related to the endoplasmic reticulum. It has membranes similar to those of the agranular endoplasmic reticulum. It usually is composed of four or more stacked layers of thin, flat, enclosed vesicles lying near one side of the nucleus. This apparatus is prominent in secretory cells, where it is located on the side of the cell from which the secretory substances are extruded. |
|
The Golgi apparatus functions in association with the endoplasmic reticulum. As shown in Figure 2-5, small 'transport vesicles' (also called endoplasmic reticulum vesicles, or ER vesicles) continually pinch off from the endoplasmic reticulum and shortly thereafter fuse with the Golgi apparatus. In this way, substances entrapped in the ER vesicles are transported from the endoplasmic reticulum to the Golgi apparatus. The transported substances are then processed in the Golgi apparatus to form lysosomes, secretory vesicles, and other cytoplasmic components that are discussed later in the chapter. |
|
||
|
|
|
Figure 2-5 A typical Golgi apparatus and its relationship to the endoplasmic reticulum (ER) and the nucleus. |
|
page 15 |
|
|
|
page 16 |
|
Lysosomes |
|
Lysosomes, shown in Figure 2-2, are vesicular
organelles that form by breaking off from the Golgi apparatus and then
dispersing throughout the cytoplasm. The lysosomes provide an intracellular
digestive system that allows the cell to digest (1) damaged cellular
structures, (2) food particles that have been ingested by the cell, and (3)
unwanted matter such as bacteria. The lysosome is quite different in
different types of cells, but it is usually 250 to 750 nanometers in
diameter. It is surrounded by a typical lipid bilayer membrane and is filled
with large numbers of small granules 5 to 8 nanometers in diameter, which are
protein aggregates of as many as 40 different hydrolase (digestive)
enzymes. A hydrolytic enzyme is capable of splitting an organic compound
into two or more parts by combining hydrogen from a water molecule with one
part of the compound and combining the hydroxyl portion of the water molecule
with the other part of the compound. For instance, protein is hydrolyzed to
form amino acids |
|
Ordinarily, the membrane surrounding the lysosome prevents the enclosed hydrolytic enzymes from coming in contact with other substances in the cell and, therefore, prevents their digestive actions. However, some conditions of the cell break the membranes of some of the lysosomes, allowing release of the digestive enzymes. These enzymes then split the organic substances with which they come in contact into small, highly diffusible substances such as amino acids and glucose. Some of the more specific functions of lysosomes are discussed later in the chapter. |
|
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|||||||
|
|
|
|
|
|
|
Peroxisomes |
|
Peroxisomes are similar physically to lysosomes, but they are different in two important ways. First, they are believed to be formed by self-replication (or perhaps by budding off from the smooth endoplasmic reticulum) rather than from the Golgi apparatus. Second, they contain oxidases rather than hydrolases. Several of the oxidases are capable of combining oxygen with hydrogen ions derived from different intracellular chemicals to form hydrogen peroxide (H2O2). Hydrogen peroxide is a highly oxidizing substance and is used in association with catalase, another oxidase enzyme present in large quantities in peroxisomes, to oxidize many substances that might otherwise be poisonous to the cell. For instance, about half the alcohol a person drinks is detoxified by the peroxisomes of the liver cells in this manner. |
|
Secretory Vesicles |
|
||
|
|
|
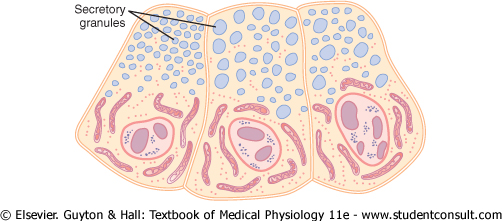
Figure 2-6 Secretory granules (secretory vesicles) in acinar cells of the pancreas. |
|
|
|
|
|
Figure 2-7 Structure of a mitochondrion. (Modified from DeRobertis EDP, Saez FA, DeRobertis EMF: Cell Biology, 6th ed. Philadelphia: WB Saunders, 1975.) |
|
One of the important functions of many cells is secretion of special chemical substances. Almost all such secretory substances are formed by the endoplasmic reticulum-Golgi apparatus system and are then released from the Golgi apparatus into the cytoplasm in the form of storage vesicles called secretory vesicles or secretory granules. Figure 2-6 shows typical secretory vesicles inside pancreatic acinar cells; these vesicles store protein proenzymes (enzymes that are not yet activated). The proenzymes are secreted later through the outer cell membrane into the pancreatic duct and thence into the duodenum, where they become activated and perform digestive functions on the food in the intestinal tract. |
|
Mitochondria |
|
The mitochondria, shown in Figures 2-2 and 2-7, are called the 'powerhouses' of the cell. Without them, cells would be unable to extract enough energy from the nutrients, and essentially all cellular functions would cease. |
|
page 16 |
|
|
|
page 17 |
|
Mitochondria are present in all areas of each cell's cytoplasm, but the total number per cell varies from less than a hundred up to several thousand, depending on the amount of energy required by the cell. Further, the mitochondria are concentrated in those portions of the cell that are responsible for the major share of its energy metabolism. They are also variable in size and shape. Some are only a few hundred nanometers in diameter and globular in shape, whereas others are elongated-as large as 1 micrometer in diameter and 7 micrometers long; still others are branching and filamentous. |
|
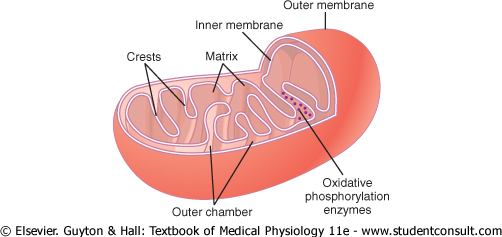
The basic structure of the mitochondrion,
shown in Figure 2-7, is composed mainly of two lipid bilayer-protein
membranes: an outer membrane and an inner membrane. Many
infoldings of the inner membrane form shelves onto which oxidative
enzymes are attached. In addition, the inner cavity of the mitochondrion is
filled with a matrix that contains large quantities of dissolved
enzymes that are necessary for extracting energy from nutrients. These
enzymes operate in association with the oxidative enzymes on the shelves to
cause oxidation of the nutrients, thereby forming carbon dioxide and water
and at the same time releasing energy. The liberated energy is used to
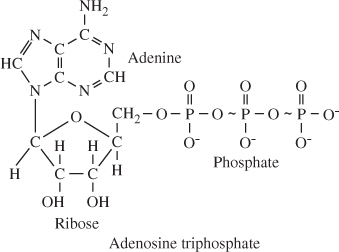
synthesize a 'high-energy' substance called adenosine |
|
Mitochondria are self-replicative, which means that one mitochondrion can form a second one, a third one, and so on, whenever there is a need in the cell for increased amounts of ATP. Indeed, the mitochondria contain DNA similar to that found in the cell nucleus. In Chapter 3 we will see that DNA is the basic chemical of the nucleus that controls replication of the cell. The DNA of the mitochondrion plays a similar role, controlling replication of the mitochondrion itself. |
|
Filament and Tubular Structures of the Cell |
|
The fibrillar proteins of the cell are usually organized into filaments or tubules. These originate as precursor protein molecules synthesized by ribosomes in the cytoplasm. The precursor molecules then polymerize to form filaments. As an example, large numbers of actin filaments frequently occur in the outer zone of the cytoplasm, called the ectoplasm, to form an elastic support for the cell membrane. Also, in muscle cells, actin and myosin filaments are organized into a special contractile machine that is the basis for muscle contraction, as discussed in detail in Chapter 6. |
|
A special type of stiff filament composed of polymerized tubulin molecules is used in all cells to construct very strong tubular structures, the microtubules. Figure 2-8 shows typical microtubules that were teased from the flagellum of a sperm. |
|
Another example of microtubules is the tubular skeletal structure in the center of each cilium that radiates upward from the cell cytoplasm to the tip of the cilium. This structure is discussed later in the chapter and is illustrated in Figure 2-17. Also, both the centrioles and the mitotic spindle of the mitosing cell are composed of stiff microtubules. |
|
Thus, a primary function of microtubules is to act as a cytoskeleton, providing rigid physical structures for certain parts of cells. |
|
|
|
Figure 2-8 Microtubules teased from the flagellum of a sperm. (From Wolstenholme GEW, O'Connor M, and The publisher, JA Churchill, 1967. Figure 4, page 314. Copyright the Novartis Foundation formerly the Ciba Foundation.) |
|
Nucleus |
|
The nucleus is the control center of the cell. Briefly, the nucleus contains large quantities of DNA, which are the genes. The genes determine the characteristics of the cell's proteins, including the structural proteins, as well as the intracellular enzymes that control cytoplasmic and nuclear activities. |
|
The genes also control and promote reproduction of the cell itself. The genes first reproduce to give two identical sets of genes; then the cell splits by a special process called mitosis to form two daughter cells, each of which receives one of the two sets of DNA genes. All these activities of the nucleus are considered in detail in the next chapter. |
|
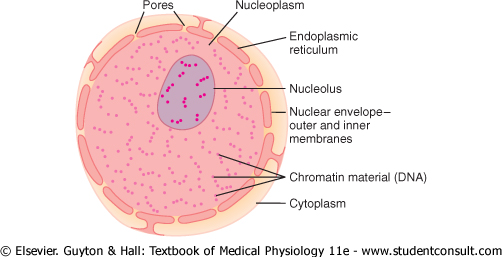
Unfortunately, the appearance of the nucleus under the microscope does not provide many clues to the mechanisms by which the nucleus performs its control activities. Figure 2-9 shows the light microscopic appearance of the interphase nucleus (during the period between mitoses), revealing darkly staining chromatin material throughout the nucleoplasm. During mitosis, the chromatin material organizes in the form of highly structured chromosomes, which can then be easily identified using the light microscope, as illustrated in the next chapter. |
|
Nuclear Membrane |
|
The nuclear membrane, also called the nuclear envelope, is actually two separate bilayer membranes, one inside the other. The outer membrane is continuous with the endoplasmic reticulum of the cell cytoplasm, and the space between the two nuclear membranes is also continuous with the space inside the endoplasmic reticulum, as shown in Figure 2-9. |
|
page 17 |
|
|
|
page 18 |
|
||
|
|
|
Figure 2-9 Structure of the nucleus. |
|
The nuclear membrane is penetrated by several thousand nuclear pores. Large complexes of protein molecules are attached at the edges of the pores so that the central area of each pore is only about 9 nanometers in diameter. Even this size is large enough to allow molecules up to 44,000 molecular weight to pass through with reasonable ease. |
|
Nucleoli and Formation of Ribosomes |
|
The nuclei of most cells contain one or more highly staining structures called nucleoli. The nucleolus, unlike most other organelles discussed here, does not have a limiting membrane. Instead, it is simply an accumulation of large amounts of RNA and proteins of the types found in ribosomes. The nucleolus becomes considerably enlarged when the cell is actively synthesizing proteins. |
|
Formation of the nucleoli (and of the ribosomes in the cytoplasm outside the nucleus) begins in the nucleus. First, specific DNA genes in the chromosomes cause RNA to be synthesized. Some of this is stored in the nucleoli, but most of it is transported outward through the nuclear pores into cytoplasm. Here, it is used in conjunction with specific proteins to assemble 'mature' ribosomes that play an essential role in forming cytoplasmic proteins, as discussed more fully in Chapter 3. |
|
|
|
|
|
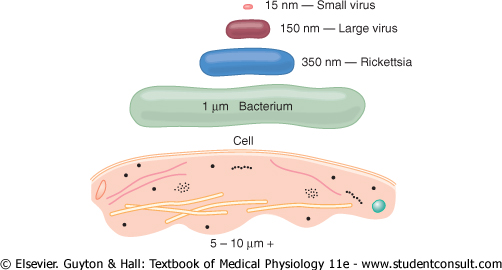
Many of us think of the cell as the lowest level of life. However, the cell is a very complicated organism that required many hundreds of millions of years to develop after the earliest form of life, an organism similar to the present-day virus, first appeared on earth. Figure 2-10 shows the relative sizes of (1) the smallest known virus, (2) a large virus, (3) a rickettsia, (4) a bacterium, and (5) a nucleated cell, demonstrating that the cell has a diameter about 1000 times that of the smallest virus and, therefore, a volume about 1 billion times that of the smallest virus. Correspondingly, the functions and anatomical organization of the cell are also far more complex than those of the virus. |
|
The essential life-giving constituent of the small virus is a nucleic acid embedded in a coat of protein. This nucleic acid is composed of the same basic nucleic acid constituents (DNA or RNA) found in mammalian cells, and it is capable of reproducing itself under appropriate conditions. Thus, the virus propagates its lineage from generation to generation and is therefore a living structure in the same way that the cell and the human being are living structures. |
|
As life evolved, other chemicals besides nucleic acid and simple proteins became integral parts of the organism, and specialized functions began to develop in different parts of the virus. A membrane formed around the virus, and inside the membrane, a fluid matrix appeared. Specialized chemicals then developed inside the fluid to perform special functions; many protein enzymes appeared that were capable of catalyzing chemical reactions and, therefore, determining the organism's activities. |
|
In still later stages of life, particularly in the rickettsial and bacterial stages, organelles developed inside the organism, representing physical structures of chemical aggregates that perform functions in a more efficient manner than can be achieved by dispersed chemicals throughout the fluid matrix. |
|
Finally, in the nucleated cell, still more complex organelles developed, the most important of which is the nucleus itself. The nucleus distinguishes this type of cell from all lower forms of life; the nucleus provides a control center for all cellular activities, and it provides for exact reproduction of new cells generation after generation, each new cell having almost exactly the same structure as its progenitor. |
|
Functional Systems of the Cell |
|
In the remainder of this chapter, we discuss several representative functional systems of the cell that make it a living organism. |
|
Ingestion by the Cell-Endocytosis |
|
If a cell is to live and grow and reproduce, it must obtain nutrients and other substances from the surrounding fluids. Most substances pass through the cell membrane by diffusion and active transport. |
|
Diffusion involves simple movement through the membrane caused by the random motion of the molecules of the substance; substances move either through cell membrane pores or, in the case of lipid-soluble substances, through the lipid matrix of the membrane. |
|
Active transport involves the actual carrying of a substance through the membrane by a physical protein structure that penetrates all the way through the membrane. These active transport mechanisms are so important to cell function that they are presented in detail in Chapter 4. |
|
Very large particles enter the cell by a specialized function of the cell membrane called endocytosis. The principal forms of endocytosis are pinocytosis and phagocytosis. Pinocytosis means ingestion of minute particles that form vesicles of extracellular fluid and particulate constituents inside the cell cytoplasm. Phagocytosis means ingestion of large particles, such as bacteria, whole cells, or portions of degenerating tissue. |
|
Pinocytosis. Pinocytosis occurs continually in the cell membranes of most cells, but it is especially rapid in some cells. For instance, it occurs so rapidly in macrophages that about 3 per cent of the total macrophage membrane is engulfed in the form of vesicles each minute. Even so, the pinocytotic vesicles are so small-usually only 100 to 200 nanometers in diameter-that most of them can be seen only with the electron microscope. |
|
Pinocytosis is the only means by which most large macromolecules, such as most protein molecules, can enter cells. In fact, the rate at which pinocytotic vesicles form is usually enhanced when such macromolecules attach to the cell membrane. |
|
|
|
|
|
Figure 2-11 Mechanism of pinocytosis. |
|
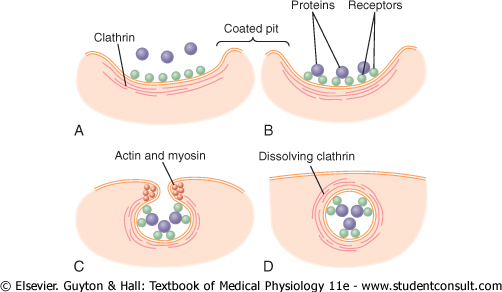
Figure 2-11 demonstrates the successive steps of pinocytosis, showing three molecules of protein attaching to the membrane. These molecules usually attach to specialized protein receptors on the surface of the membrane that are specific for the type of protein that is to be absorbed. The receptors generally are concentrated in small pits on the outer surface of the cell membrane, called coated pits. On the inside of the cell membrane beneath these pits is a latticework of fibrillar protein called clathrin, as well as other proteins, perhaps including contractile filaments of actin and myosin. Once the protein molecules have bound with the receptors, the surface properties of the local membrane change in such a way that the entire pit invaginates inward, and the fibrillar proteins surrounding the invaginating pit cause its borders to close over the attached proteins as well as over a small amount of extracellular fluid. Immediately thereafter, the invaginated portion of the membrane breaks away from the surface of the cell, forming a pinocytotic vesicle inside the cytoplasm of the cell. |
|
What causes the cell membrane to go through the necessary contortions to form pinocytotic vesicles remains mainly a mystery. This process requires energy from within the cell; this is supplied by ATP, a high-energy substance discussed later in the chapter. Also, it requires the presence of calcium ions in the extracellular fluid, which probably react with contractile protein filaments beneath the coated pits to provide the force for pinching the vesicles away from the cell membrane. |
|
Phagocytosis. Phagocytosis occurs in much the same way as pinocytosis, except that it involves large particles rather than molecules. Only certain cells have the capability of phagocytosis, most notably the tissue macrophages and some of the white blood cells. |
|
Phagocytosis is initiated when a particle such as a bacterium, a dead cell, or tissue debris binds with receptors on the surface of the phagocyte. In the case of bacteria, each bacterium usually is already attached to a specific antibody, and it is the antibody that attaches to the phagocyte receptors, dragging the bacterium along with it. This intermediation of antibodies is called opsonization, which is discussed in Chapters 33 and 34. |
|
page 19 |
|
|
|
page 20 |
|
Phagocytosis occurs in the following steps:
|
|
Digestion of Pinocytotic and Phagocytic Foreign Substances Inside the Cell-Function of the Lysosomes |
|
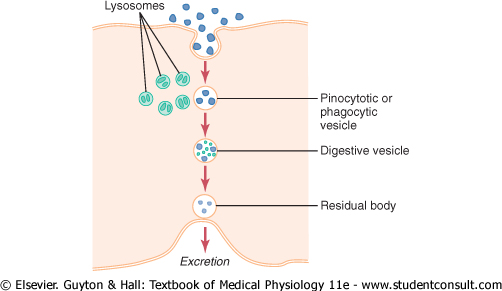
Almost immediately after a pinocytotic or
phagocytic vesicle appears inside a cell, one or more lysosomes become
attached to the vesicle and empty their acid hydrolases to the inside
of the vesicle, as shown in Figure 2-12. Thus, a digestive vesicle is
formed inside the cell cytoplasm in which the vesicular hydrolases begin
hydrolyzing the proteins, carbohydrates, lipids, and other substances in the
vesicle. The products of digestion are small molecules of amino acids |
|
Thus, the pinocytotic and phagocytic vesicles containing lysosomes can be called the digestive organs of the cells. |
|
|
|
|
|
Figure 2-12 Digestion of substances in pinocytotic or phagocytic vesicles by enzymes derived from lysosomes. |
|
Regression of Tissues and Autolysis of Cells. Tissues of the body often regress to a smaller size. For instance, this occurs in the uterus after pregnancy, in muscles during long periods of inactivity, and in mammary glands at the end of lactation. Lysosomes are responsible for much of this regression. The mechanism by which lack of activity in a tissue causes the lysosomes to increase their activity is unknown. |
|
Another special role of the lysosomes is removal of damaged cells or damaged portions of cells from tissues. Damage to the cell-caused by heat, cold, trauma, chemicals, or any other factor-induces lysosomes to rupture. The released hydrolases immediately begin to digest the surrounding organic substances. If the damage is slight, only a portion of the cell is removed, followed by repair of the cell. If the damage is severe, the entire cell is digested, a process called autolysis. In this way, the cell is completely removed, and a new cell of the same type ordinarily is formed by mitotic reproduction of an adjacent cell to take the place of the old one. |
|
The lysosomes also contain bactericidal agents that can kill phagocytized bacteria before they can cause cellular damage. These agents include (1) lysozyme, which dissolves the bacterial cell membrane; (2) lysoferrin, which binds iron and other substances before they can promote bacterial growth; and (3) acid at a pH of about 5.0, which activates the hydrolases and inactivates bacterial metabolic systems. |
|
Synthesis and Formation of Cellular Structures by Endoplasmic Reticulum and Golgi Apparatus |
|
Specific Functions of the Endoplasmic Reticulum |
|
The extensiveness of the endoplasmic reticulum and the Golgi apparatus in secretory cells has already been emphasized. These structures are formed primarily of lipid bilayer membranes similar to the cell membrane, and their walls are loaded with protein enzymes that catalyze the synthesis of many substances required by the cell. |
|
Most synthesis begins in the endoplasmic reticulum. The products formed there are then passed on to the Golgi apparatus, where they are further processed before being released into the cytoplasm. But first, let us note the specific products that are synthesized in specific portions of the endoplasmic reticulum and the Golgi apparatus. |
|
Proteins Are Formed by the Granular Endoplasmic Reticulum. The granular portion of the endoplasmic reticulum is characterized by large numbers of ribosomes attached to the outer surfaces of the endoplasmic reticulum membrane. As we discuss in Chapter 3, protein molecules are synthesized within the structures of the ribosomes. The ribosomes extrude some of the synthesized protein molecules directly into the cytosol, but they also extrude many more through the wall of the endoplasmic reticulum to the interior of the endoplasmic vesicles and tubules, that is, into the endoplasmic matrix. |
|
page 20 |
|
|
|
page 21 |
|
Synthesis of Lipids by the Smooth Endoplasmic Reticulum. The endoplasmic reticulum also synthesizes lipids, especially phospholipids and cholesterol. These are rapidly incorporated into the lipid bilayer of the endoplasmic reticulum itself, thus causing the endoplasmic reticulum to grow more extensive. This occurs mainly in the smooth portion of the endoplasmic reticulum. |
|
To keep the endoplasmic reticulum from growing beyond the needs of the cell, small vesicles called ER vesicles or transport vesicles continually break away from the smooth reticulum; most of these vesicles then migrate rapidly to the Golgi apparatus. |
|
Other Functions of the Endoplasmic Reticulum. Other significant functions of the endoplasmic reticulum, especially the smooth reticulum, include the following:
|
|
Specific Functions of the Golgi Apparatus |
|
Synthetic Functions of the Golgi Apparatus. Although the major function of the Golgi apparatus is to provide additional processing of substances already formed in the endoplasmic reticulum, it also has the capability of synthesizing certain carbohydrates that cannot be formed in the endoplasmic reticulum. This is especially true for the formation of large saccharide polymers bound with small amounts of protein; the most important of these are hyaluronic acid and chondroitin sulfate. |
|
A few of the many functions of hyaluronic acid and chondroitin sulfate in the body are as follows: (1) they are the major components of proteoglycans secreted in mucus and other glandular secretions; (2) they are the major components of the ground substance outside the cells in the interstitial spaces, acting as filler between collagen fibers and cells; and (3) they are principal components of the organic matrix in both cartilage and bone. |
|
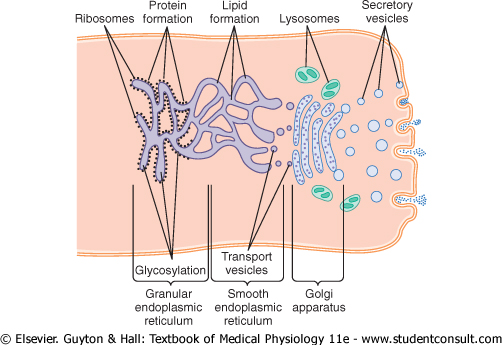
Processing of Endoplasmic Secretions by the Golgi Apparatus-Formation of Vesicles. Figure 2-13 summarizes the major functions of the endoplasmic reticulum and Golgi apparatus. As substances are formed in the endoplasmic reticulum, especially the proteins, they are transported through the tubules toward portions of the smooth endoplasmic reticulum that lie nearest the Golgi apparatus. At this point, small transport vesicles composed of small envelopes of smooth endoplasmic reticulum continually break away and diffuse to the deepest layer of the Golgi apparatus. Inside these vesicles are the synthesized proteins and other products from the endoplasmic reticulum. |
|
|
|
|
|
Figure 2-13 Formation of proteins, lipids, and cellular vesicles by the endoplasmic reticulum and Golgi apparatus. |
|
The transport vesicles instantly fuse with the Golgi apparatus and empty their contained substances into the vesicular spaces of the Golgi apparatus. Here, additional carbohydrate moieties are added to the secretions. Also, an important function of the Golgi apparatus is to compact the endoplasmic reticular secretions into highly concentrated packets. As the secretions pass toward the outermost layers of the Golgi apparatus, the compaction and processing proceed. Finally, both small and large vesicles continually break away from the Golgi apparatus, carrying with them the compacted secretory substances, and in turn, the vesicles diffuse throughout the cell. |
|
To give an idea of the timing of these
processes: When a glandular cell is bathed in radioactive amino acids |
|
Types of Vesicles Formed by the Golgi Apparatus-Secretory Vesicles and Lysosomes. In a highly secretory cell, the vesicles formed by the Golgi apparatus are mainly secretory vesicles containing protein substances that are to be secreted through the surface of the cell membrane. These secretory vesicles first diffuse to the cell membrane, then fuse with it and empty their substances to the exterior by the mechanism called exocytosis. Exocytosis, in most cases, is stimulated by the entry of calcium ions into the cell; calcium ions interact with the vesicular membrane in some way that is not understood and cause its fusion with the cell membrane, followed by exocytosis-that is, opening of the membrane's outer surface and extrusion of its contents outside the cell. |
|
Some vesicles, however, are destined for intracellular use. |
|
page 21 |
|
|
|
page 22 |
|
Use of Intracellular Vesicles to Replenish Cellular Membranes. Some of the intracellular vesicles formed by the Golgi apparatus fuse with the cell membrane or with the membranes of intracellular structures such as the mitochondria and even the endoplasmic reticulum. This increases the expanse of these membranes and thereby replenishes the membranes as they are used up. For instance, the cell membrane loses much of its substance every time it forms a phagocytic or pinocytotic vesicle, and the vesicular membranes of the Golgi apparatus continually replenish the cell membrane. |
|
In summary, the membranous system of the endoplasmic reticulum and Golgi apparatus represents a highly metabolic organ capable of forming new intracellular structures as well as secretory substances to be extruded from the cell. |
|
Extraction of Energy from Nutrients-Function of the Mitochondria |
|
|||
|
|
|
Figure 2-14 Formation
of adenosine |
|
The principal substances from which cells
extract energy are foodstuffs that react chemically with
oxygen-carbohydrates, fats, and proteins. In the human body, essentially all
carbohydrates are converted into glucose |
|
Briefly, almost all these oxidative reactions occur inside the mitochondria, and the energy that is released is used to form the high-energy compound ATP. Then, ATP, not the original foodstuffs, is used throughout the cell to energize almost all the subsequent intracellular metabolic reactions. |
|
Functional Characteristics of ATP |
|
|
|
When ATP releases its energy, a phosphoric
acid radical is split away, and adenosine |
|
To reconstitute the cellular ATP as it is used up, energy derived from the cellular nutrients causes ADP and phosphoric acid to recombine to form new ATP, and the entire process repeats over and over again. For these reasons, ATP has been called the energy currency of the cell because it can be spent and remade continually, having a turnover time of only a few minutes. |
|
page 22 |
|
|
|
page 23 |
|
Chemical Processes in the Formation of
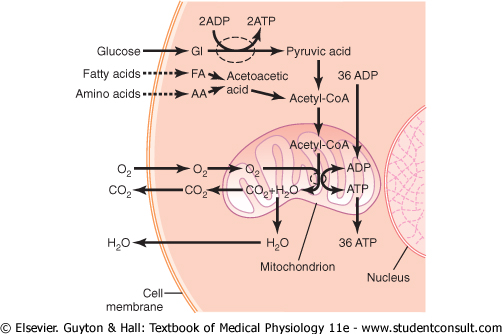
ATP-Role of the Mitochondria. On entry into the cells, glucose |
|
By far, the major portion of the ATP formed in
the cell, about 95 per cent, is formed in the mitochondria. The pyruvic acid
derived from carbohydrates, fatty acids from lipids, and amino acids |
|
In this citric acid cycle, acetyl-CoA is split into its component parts, hydrogen atoms and carbon dioxide. The carbon dioxide diffuses out of the mitochondria and eventually out of the cell; finally, it is excreted from the body through the lungs. |
|
The hydrogen atoms, conversely, are highly reactive, and they combine instantly with oxygen that has also diffused into the mitochondria. This releases a tremendous amount of energy, which is used by the mitochondria to convert very large amounts of ADP to ATP. The processes of these reactions are complex, requiring the participation of large numbers of protein enzymes that are integral parts of mitochondrial membranous shelves that protrude into the mitochondrial matrix. The initial event is removal of an electron from the hydrogen atom, thus converting it to a hydrogen ion. The terminal event is combination of hydrogen ions with oxygen to form water plus the release of tremendous amounts of energy to large globular proteins, called ATP synthetase, that protrude like knobs from the membranes of the mitochondrial shelves. Finally, the enzyme ATP synthetase uses the energy from the hydrogen ions to cause the conversion of ADP to ATP. The newly formed ATP is transported out of the mitochondria into all parts of the cell cytoplasm and nucleoplasm, where its energy is used to energize multiple cell functions. |
|
This overall process for formation of ATP is called the chemiosmotic mechanism of ATP formation. The chemical and physical details of this mechanism are presented in Chapter 67, and many of the detailed metabolic functions of ATP in the body are presented in Chapters 67 through 71. |
|
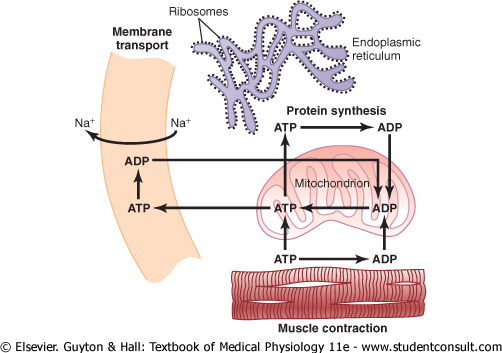
Uses of ATP for Cellular Function. Energy from ATP is used to promote three major categories of cellular functions: (1) transport of substances through multiple membranes in the cell, (2) synthesis of chemical compounds throughout the cell, and (3) mechanical work. These uses of ATP are illustrated by examples in Figure 2-15: (1) to supply energy for the transport of sodium through the cell membrane, (2) to promote protein synthesis by the ribosomes, and (3) to supply the energy needed during muscle contraction. |
|
|||
|
|
|
Figure 2-15 Use of adenosine |
|
In addition to membrane transport of sodium, energy from ATP is required for membrane transport of potassium ions, calcium ions, magnesium ions, phosphate ions, chloride ions, urate ions, hydrogen ions, and many other ions and various organic substances. Membrane transport is so important to cell function that some cells-the renal tubular cells, for instance-use as much as 80 per cent of the ATP that they form for this purpose alone. |
|
In addition to synthesizing proteins, cells
synthesize phospholipids, cholesterol, purines, pyrimidines, and a host of
other substances. Synthesis of almost any chemical compound requires energy.
For instance, a single protein molecule might be composed of as many as
several thousand amino acids |
|
The final major use of ATP is to supply energy for special cells to perform mechanical work. We see in Chapter 6 that each contraction of a muscle fiber requires expenditure of tremendous quantities of ATP energy. Other cells perform mechanical work in other ways, especially by ciliary and ameboid motion, which are described later in this chapter. The source of energy for all these types of mechanical work is ATP. |
|
page 23 |
|
|
|
page 24 |
|
In summary, ATP is always available to release its energy rapidly and almost explosively wherever in the cell it is needed. To replace the ATP used by the cell, much slower chemical reactions break down carbohydrates, fats, and proteins and use the energy derived from these to form new ATP. More than 95 per cent of this ATP is formed in the mitochondria, which accounts for the mitochondria being called the 'powerhouses' of the cell. |
|
Locomotion of Cells |
|
By far the most important type of movement that occurs in the body is that of the muscle cells in skeletal, cardiac, and smooth muscle, which constitute almost 50 per cent of the entire body mass. The specialized functions of these cells are discussed in Chapters 6 through 9. Two other types of movement-ameboid locomotion and ciliary movement-occur in other cells. |
|
Ameboid Movement |
|
Ameboid movement is movement of an entire cell in relation to its surroundings, such as movement of white blood cells through tissues. It receives its name from the fact that amebae move in this manner and have provided an excellent tool for studying the phenomenon. |
|
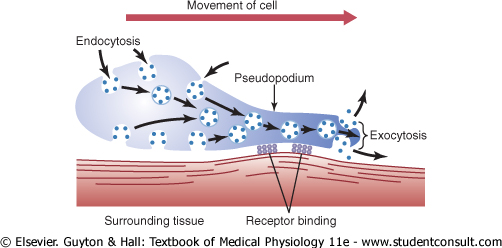
Typically, ameboid locomotion begins with protrusion of a pseudopodium from one end of the cell. The pseudopodium projects far out, away from the cell body, and partially secures itself in a new tissue area. Then the remainder of the cell is pulled toward the pseudopodium. Figure 2-16 demonstrates this process, showing an elongated cell, the right-hand end of which is a protruding pseudopodium. The membrane of this end of the cell is continually moving forward, and the membrane at the left-hand end of the cell is continually following along as the cell moves. |
|
||
|
Figure 2-16 Ameboid motion by a cell. |
|
Mechanism of Ameboid Locomotion. Figure 2-16 shows the general principle of ameboid motion. Basically, it results from continual formation of new cell membrane at the leading edge of the pseudopodium and continual absorption of the membrane in mid and rear portions of the cell. Also, two other effects are essential for forward movement of the cell. The first effect is attachment of the pseudopodium to surrounding tissues so that it becomes fixed in its leading position, while the remainder of the cell body is pulled forward toward the point of attachment. This attachment is effected by receptor proteins that line the insides of exocytotic vesicles. When the vesicles become part of the pseudopodial membrane, they open so that their insides evert to the outside, and the receptors now protrude to the outside and attach to ligands in the surrounding tissues. |
|
At the opposite end of the cell, the receptors pull away from their ligands and form new endocytotic vesicles. Then, inside the cell, these vesicles stream toward the pseudopodial end of the cell, where they are used to form still new membrane for the pseudopodium. |
|
The second essential effect for locomotion is to provide the energy required to pull the cell body in the direction of the pseudopodium. Experiments suggest the following as an explanation: In the cytoplasm of all cells is a moderate to large amount of the protein actin. Much of the actin is in the form of single molecules that do not provide any motive power; however, these polymerize to form a filamentous network, and the network contracts when it binds with an actin-binding protein such as myosin. The whole process is energized by the high-energy compound ATP. This is what happens in the pseudopodium of a moving cell, where such a network of actin filaments forms anew inside the enlarging pseudopodium. Contraction also occurs in the ectoplasm of the cell body, where a preexisting actin network is already present beneath the cell membrane. |
|
Types of Cells That Exhibit Ameboid Locomotion. The most common cells to exhibit ameboid locomotion in the human body are the white blood cells when they move out of the blood into the tissues in the form of tissue macrophages. Other types of cells can also move by ameboid locomotion under certain circumstances. For instance, fibroblasts move into a damaged area to help repair the damage, and even the germinal cells of the skin, though ordinarily completely sessile cells, move toward a cut area to repair the rent. Finally, cell locomotion is especially important in development of the embryo and fetus after fertilization of an ovum. For instance, embryonic cells often must migrate long distances from their sites of origin to new areas during development of special structures. |
|
Control of Ameboid Locomotion-Chemotaxis. The most important initiator of ameboid locomotion is the process called chemotaxis. This results from the appearance of certain chemical substances in the tissues. Any chemical substance that causes chemotaxis to occur is called a chemotactic substance. Most cells that exhibit ameboid locomotion move toward the source of a chemotactic substance-that is, from an area of lower concentration toward an area of higher concentration-which is called positive chemotaxis. Some cells move away from the source, which is called negative chemotaxis. |
|
But how does chemotaxis control the direction of ameboid locomotion? Although the answer is not certain, it is known that the side of the cell most exposed to the chemotactic substance develops membrane changes that cause pseudopodial protrusion. |
|
Cilia and Ciliary Movements |
|
page 24 |
|
|
|
page 25 |
|
||
|
|
|
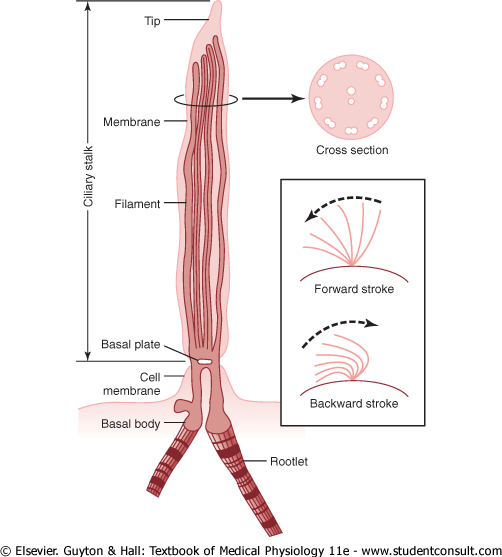
Figure 2-17 Structure and function of the cilium. (Modified from Satir P: Cilia. Sci Am 204:108, 1961. Copyright Donald Garber: Executor of the estate of Bunji Tagawa.) |
|
A second type of cellular motion, ciliary movement, is a whiplike movement of cilia on the surfaces of cells. This occurs in only two places in the human body: on the sufaces of the respiratory airways and on the inside surfaces of the uterine tubes (fallopian tubes) of the reproductive tract. In the nasal cavity and lower respiratory airways, the whiplike motion of cilia causes a layer of mucus to move at a rate of about 1 cm/min toward the pharynx, in this way continually clearing these passageways of mucus and particles that have become trapped in the mucus. In the uterine tubes, the cilia cause slow movement of fluid from the ostium of the uterine tube toward the uterus cavity; this movement of fluid transports the ovum from the ovary to the uterus. |
|
As shown in Figure 2-17, a cilium has the appearance of a sharp-pointed straight or curved hair that projects 2 to 4 micrometers from the surface of the cell. Many cilia often project from a single cell-for instance, as many as 200 cilia on the surface of each epithelial cell inside the respiratory passageways. The cilium is covered by an outcropping of the cell membrane, and it is supported by 11 microtubules-9 double tubules located around the periphery of the cilium, and 2 single tubules down the center, as demonstrated in the cross section shown in Figure 2-17. Each cilium is an outgrowth of a structure that lies immediately beneath the cell membrane, called the basal body of the cilium. |
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 2561
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved