| CATEGORII DOCUMENTE |
| Alimentatie nutritie | Asistenta sociala | Cosmetica frumusete | Logopedie | Retete culinare | Sport |
ETIOLOGIE - TRICHINELOZA
Clasificarea zoologica
Trichineloza este o boala parazitara produsa de viermi din specii diferite,dar care apartin aceluiasi gen-Trichinella.
Subincrengatura: Nemathelmintes
Clasa: Nematoda
Subclasa: Adenophorea
Ordinul: Enoplida
Suprafamilia: Trichinelloidea
Familia: Trichinellidae
Subfamilia: Trichinellinae
Genul: Trichinella- Raillet 1985,sin. Trichina
Trichina-Owen 1835
Specii: Trichinella spiralis (Raillet, 1985);
Trichinella pseudospiralis (Garkovi, 1972);
Trichinella nativa (Britov si Boev, 1972);
Trichinella nelson (Britov si Boev, 1972);
Trichinella britovi (Britov si Boev, 1972);
Trichinella murelli;
Trichinella papuae;
Trichinella zimbabwensis.
Mult timp s-a considerat ca genul Trichinella cuprinde o singura specie Trichinella spiralis- si ca trichineloza la om si animale este provocata de aceasta unica specie. In ultimele decenii s-a conturat ideea ca genul Trichinella cuprinde patru specii surori ("sibling species") sau doua specii - Trichinella spiralis si Trichinella pseudospiralis - prima specie - Trichinella spiralis - avand trei variante intraspecie: Trichinella spiralis domestica, Trichinella spiralis nelson si Trichinella spiralis nativa.
Mai recent, unii cercetatori au considerat ca genul Trichinella cuprinde doua specii complet diferite: Trichinella spiralis si Trichinella pseudospiralis, iar in cadrul speciei Trichinella spiralis ar exista 7 subspecii sau variante notate cu siglele T1, T2, T3, T4, T5, T6, T7 si T8. Sigla T4 este rezervata pentru Trichinella pseudospiralis.
1.2 Specia genului Trichinella
In present sunt recunoscute official 8 specii de Trichinella, din care trei specii (Trichinella pseudospiralis, Trichinella papuae si Trichinella zimbabwensis) nu produc capsula.
Trichinella spiralis ( numita de altfel T1 ) este raspandita in regiunile temperate ale globului si este in general asociata porcului domestic cu un grad inalt de infectivitate pentru porci, soareci si sobolani.
Trichinella nativa (T2) si subspeciile sale, Trichinella T6, gasita in America de Nord, este o specie adaptata la climatul rece. Are capacitate (limitata) scazuta de infectie la porci, dar este frecvent intalnita la carnasierele salbatice ursi si morsa; in plus se distinge prin rezistenta la temperaturi scazute (inghet).
Trichinella britovi (T3) este intalnita predominant (cu frecventa ) la animalele salbatice, desi este gasita ocazional la porci si cai. Se gaseste in regiunile temperate din Europa si Asia. Trichinella britovi si genotipurile inrudite, Trichinella T8 din Africa de Sud si Trichinella T9 din Japonia, cu cateva caracteristici intermediare cu alte specii, incluzand slaba rezistenta la frig, infectivitate marita la suine si o slaba reprezentare a capsulei (larvele pot fi confundate cu speciile neincapsulate in unele cazuri).
Trichinella murelli (T5) este o specie Nord Americana intalnita la animalele salbatice ocazional la cai si oameni. Are capsula, putere de infectivitate la porcii domestici, insa reprezinta un risc ridicat pentru oamenii ce consuma carne de vanat.
Trichinella nelsoni (T7) a fosy izolata sporadic la animalele salbatice din Africa. Se caracterizeaza prin rezistenta mate la temperaturi ridicate comparativ cu alte specii de Trichinella.
Trei specii de Trichinella nu formeaza capsula de colagen in musculatura si anume:
Trichinella pseudospiralis (T4) cu o distributie cosmopolita, a fost descoperita la pasarile rapitoare, carnivore si omnivore salbatice, sobolani si marsupial din Asia, America de Nord, Europa si subcontinentul Australian.
Trichinella papuae (T10) este a doua specie care nu formeaza capsula. A fost descoperita doar la porcii salbatici si la oamenii din Papua Noua Guinee. Este rezistenta la congelare, are infestivitate scazuta pentru porci, a fost intalnita la o varietate de mamifere salbatice si a fost implicate in imbolnaviri la om.
Trichinella zimbabwensis (T11) a fost recent descoperita (pusa in evidenta) la crocodilii din crescatorie din Zimbabwe. Speciile pot infesta porcii si sobolanii.
Toate speciile si genotipurile de Trichinella produc imbolnaviri la om.
Criteriile principale care au stat la baza sustinerii ideii de pluralitate a speciilor in cadrul genului Trichinella au fost urmatoarele:
a. Capacitatea lor de incrucisare, dar cu diferente intre ele. Incrucisarea intre Trichinella Spiralis si Trichinella pseudospiralis, determina descendenti incapsulati si neinfectati pentru pasari. Incrucisarile au reusit de asemenea si intre celelalte specii.
b. Capacitatea lor de a se reproduce in anumite gazed, apreciata prin indicele capacitatii de reproducere (ICR) definit ca raportul dintre numarul de larve intramusculare si numarul de larve administrate. Acest character nu este insa legat numai de parazit, el depinde, de asemenea, de capacitatea imunologica a gazdelor.
c. Profilul lor enzimatic care se poate modifica in cursul pasajelor pe animale, asa incat un criteriu care ar permite diferentierea a doua specii devine identic la acele doua specii.
d. Analiza electroforetica a proteinelor si polipeptidelor larvare si prepararea de anticorpi monoclonali care recunosc aceste elemente.
e. Analiza AND-ului lor obtinut prin amplificarea a 10 baze AND
f. Njmarul de larve 1 nou - nascute (L In.n) produse de femeie.
g. Durata necesara dezvoltarii celulei hranitoare.
In legatura cu specificatia la Trichinella se pot face urmatoarele observatii privind distributia diferitelor specii.
In Europa se gasesc Trichinella spiralis si Trichinella britovi. Trichinella britovi se gaseste in principal in Franta unde 7% din vulpi sunt parazitate iar in Italia si Elvetia nu se cunoaste decat Trichinella britoci, cu exceptia mistretilor care sunt infestati cu Trichinella spiralis de provenienta franceza.
In Spania Trichinella spiralis este raspandita in toata tara, iar Trichinella britovi nu s-a semnalat decat la mistreti la altitudini mai mari de 800 m.
In SUA nu se cunoaste decat Trichinella spiralis.
In Africa sud-sahariana exista doua specii Trichinella nelson ecuatoriana si Trichinella britovi (T8) in Africa de Sud.
In zonele arctice Trichinella native s-a gasit la ursul alb (Thalarctos maritimus) si la alte mamifere salbatice, terestre si marine.
In Romania specia obisnuita este Trichinella spiralis intalnita destul de frecvent la numeroase specii de animale domestice si salbatice, ca si la om.
1.3 Particularitatile morfologice ale genului Trichinella
Toate speciile sau subspeciile (variante) genului Trichinella au caractere morfologice identice.
Trichinella spiralis este un nematode inegal calibrat, avand partea anterioara mai subtire decat cea posterioara. Corpul parazitului este compus dintr-o cavitate generala bine dezvoltata si un perete musculo-membranos. In cavitatea generala se gaseste lichidul celomic, in care se afla dispuse organele interne. Invelisul extern este format dintr-o cuticula care contine in structura sa albumin, fibroina, substante mucoide, colagen si cheratina. Este transparent si elastic.
Invelisul intern situate sub cuticula format din hypoderm. In grosimea acestuia se gasesc patru ingrosari dispuse pe toata lungimea corpului, proeminand in interior sub forma a patru cordoane (cele situate lateral se numesc campuri laterale, iar cele dispuse dorsal si ventral se numesc "linii"). In aceste cordoane se gasesc unele cellule cu rol in sistemul nervos.
Musculatura, de tip holomiar, situate sub hypoderm este la randul ei impartita de cele patru cordoane hipodermice in patru "campuri". Fiecare camp muscular este alcatuit din 4-5 celule alungite a caror portiune bazala este contractila. La partea superioara a corpului celulele muscular au o forma semilunara si sunt situate cu convexitatea spre zona caudal. Prin forma si dispozitia lor, celulele fac legatura intre campul muscular dorso-lateral sic el ventro-lateral. Cuticula, hipodermul si musculature formeaza peretele corpului, care delimiteaza in interior cavitatea generala in care se afla tubul digestive si organelle de reproducere.
Sistemul nervos este de tip ganglionar fiind format dintr-un inel periesofagin alcatuit din sase ganglion. Se mentioneaza existent unui inel cephalic din care pleaca patru nervi principali: doi laterali, unul dorsal si unul ventral. Celulele nervoase sunt mai numeroase in partea anterioara a parazitului, iar in restul corpului in special in zona orificiului vulvar si la extremitatea posterioara a corpului sunt isolate.
Esofagul este de tip stongiloid si este format dintr-o portiune anterioara scurta si o zona bulbar captusita de o rasfrangere a cuticulei. Urmeaza un conduct ingust care poate fi flexat sub forma literei S. Apoi esofagulse dilate formand un pseudobulb alungit, cu peretii grosi si musculosi. Dupa aceasta formatiune esofagul apare ingustat si inconjurat de stichosoma sau corpul celular. Pe intreaga sa lungime esofagul are un lumen triradiat, delimitat de un perete musculos. Corpul cellular este alcatuit dintr-un numar variabil de cellule, in medie 55, denumite "stichocite". Stichocitele contin granule alfa si beta, iar continutul este evacuat printr-un canalicul in esofag. La adulti, stichocitele prezinta o activitate constanta in multiplicarea numarului mitocondriilor si in dezvoltarea reticulului endoplasmatic.
Intestinul mediu continua esofagul si are prima portiune dilatata in forma de para, constituind "stomacul". In continuare lumenul apare ingustat. La limita dintre esofag si intestinal mediu, se gasesc doua glande unicelulare, asimetrice, cu rol in digestive. Intestinul este captusit de un epiteliu unistratificat ale carui cellule trimit spre lumen prelungiri protoplasmatice care au rol in absorbtie. Intestinul terminal difera ca alcatuire de la sex la sex. La mascul, acesta se uneste cu canalul deferent formand cloaca, care are orificiul terminal situate intre cele doua perechi de papile. La femela, intestinal terminal se deschide in exterior prin orificiul anal.
1.3.1 Masculul de Trichinella spiralis
Masculul are o lungime de 1.0 - 1.6 mm si o grosime de 0.033 - 0.040 mm (circa 40 microni). In conditii de parazitism masiv, dimensiunile pot fi reduse. Corpul are o forma cilindro-conica, mai subtire la capatul anterior si mai ingrosat la cel posterior. La partea anterioara se afla orificiul bucal, cu un diametru de 2 microni. La ambele sexe se remarca prezenta unui stilet bucal retractile, lung de 7 microni si lat de un micron. La capatul posterior, masculul prezinta o pensa copulatoare alcatuita din doi apendici conici, chitinosi si curbati ventral. Intre ei se afla orificiul cloacal flancat de patru papile, din care doua sunt hemisferice si sunt situate ventral, iar doua au forma conica si sunt amplasate dorsal.
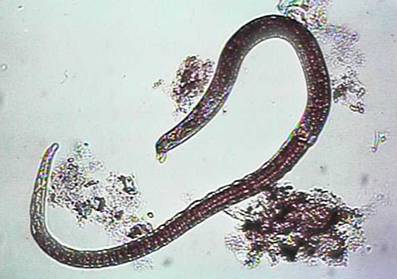
Aparatul reproducator, de tip tubular, este alcatuit din testicul, canal deferent si vezicula seminal. Partea terminal a intestinului se uneste cu portiunea terminala a veziculei seminal si creeaza canalul cloacal, care se deschide printr-un orificiu situate intre cei doi apendici copulatori.
Testiculul este captusit de un tesut epiterial cu rol germinativ din cellule reproducatoare. Spermatozoizii sunt adunati in vezicula seminal care se destined mult in grosime si sunt eliminate prin orificiul cloacal. Lungime maxima a testicolului este de 653 microni, iar grosimea de 35 microni.
1.3.2 Femela de Trichinella spiralis
Femela are o lungime de 2.2 - 4.0 mm si o grosime de 60 - 7- micron.
Corpul are forma cilindro-conica, fiind efilat la partea anterioara si mai gros la capatul posterior, unde se termina brusc in unghi obtuz.
Aparatul reproducator este de tip tubular si este alcatuit dintr-un singur ovar ingust, situate in apropierea orificiului anal, un oviduct scurt, uter si vagin. Ultimul se deschide in cincimea anterioara a corpului prin orificiul vulvar. Ovarul, lung de maxim 836 microni, este tapisat pe toata lungimea interna de epitelui germinativ care are rol in producerea si eliminarea ovocitelor.

Uterul prezinta o dilatatie numita receptacol seminal si are rol in a adaposti spermatozoizii dupa actul copulator. In receptacolul seminal are loc fecundatia ovocitelor si formarea oualor, din care ies larvele, care elterior trec in canalul vaginal si apoi in exterior.
Oul are forma ovalara, fiind acoperit de o membrane extreme de fina, transparent si elastic. Practic nu are coaja, asa cum se intalneste la celelalte nematode. Lungimea oului este cuprinsa intre 15 si 20 de micron. Prezinta in interiorul embrionul care se elibereaza de invelis in miscari active, fiind eliminate in mediul exterior. Trichinella pseudospiralis are dimensiuni mai mici decat Trichinella spiralis : 0.6 - 1.03 mm la female.
1.4 Ciclul evolutiv (biologic)
Trichinella spiralis imbogateste ciclul biologic evolutiv al clasei Nematoda printr-un mod propriu de comportament, care da nastere in biologia acestei clase, ciclului autoheteroxen.
Evolutia Trichinelei spiralis in momentul in care larva infestanta de obicei incapsulata, dar si libera, in stare viabila este ingeratade organismul in care acest parazit permanent patrunde si care devine in acelasi timp atat gazda definitive (prin formele adulte ce se gasesc in intestine unde se dezvolta si are loc ponta larvelor), cat si gazda intermediara (in muschiul striat, unde se cantoneaza formele larvare infestate).
Gazda definitive a trichinelelor devine, invariabil si obligatoriu gazda intermediara.
Astfel, ciclul evolutiv la Trichinella spiralis cuprinde urmatoarele faze:
Faza intestinala
Faza de migrare sau diseminare
Faza de invazie si localizare musculara
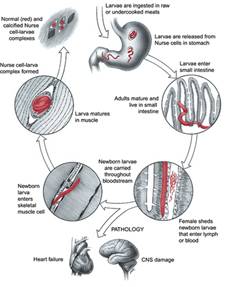
1.4.1 Faza intestinala
In decurs de 3- 4 ore din momentul ingerarii carnii infectata cu larve sau numai a larvelor infectate, in stomacul noii gazed incepe digerarea fibrelor muscular si a capsule ce inconjoara larva, devenind libera. Digestia acestora are loc sub actiunea sucului gastric, si probabil si de catre unele enzyme secretate de larva inchistata, enzyme care pot fi active sub influenta conditiilor existente in stomacul gazdei.
Deschiderea capsule se face in zona in care se afla extremitatea posterioara a larvei, justificand in felul acesta afirmatia de mai sus. O data eliberate, larvele libere trec in intestinal subtire si patrund printer vilozitatile mucoasei doudenale si jejunale astfel ca la 3 ore de la infestatie, o parte din larve se gasesc deja in epiteliul intestinal dupa care patrund si sunt cuibarite in niste formate prin fuzionarea a peste 100 de cellule formand in epiteliul intestinal un sincitiu.
In aceste conditii larvele se hranesc cu exsudat cellular si continut intestinal, dezvoltandu-le rapid. Larvele vii encapsulate trec neafectate prin stomac si urmeaza aceeasi cale de dezvoltare.
Caracteristic Trichinellei spiralis este dezvoltarea ei extrem de rapida, care dureaza numai 48 de ore pana ce larva devine adult, ca apoi, in 2-3 zile sa aiba loc dezvoltarea organelor reproducatoare, devenind adult apt pentru reproducere. Evolutia de la larva la adult are loc in etape, marcate prin cate un process de naparlire de regula patru naparliri ultima naparlire avand loc atunci cand toate componentele organelor de reproductive sunt complet dezvoltate.
In mucoasa intestinala adultii tineri se copuleaza in nisa celulara formata in epiteliul intestinal, femelele ajung la glandele limfatice fiind in numar de 1.5-2 ori mai mare decat cel al masculilor, astfel incat un mascul insamanteaza in medie doua femele.
Comportamentul sexual al trichinelelor are la baza atractia mediata de feromoni. In vitro s-a dovedit ca viermii femela produc o molecula hidrosolubila cu proprietati atractante pentru mascul. Masculii pot produce un repellent care tine alti masculi la oarecare distanta.
Numarul de larve depuse de femela variaza in functie de gazda. Astfel la rozatoare la care femela depune 200 - 1600 larve. La porc pentru fiecare larva infestata administrate se produc 105 - 112 larve musculare.
De interes pentru immunodiagnostic sunt stichocitele si secretia lor. Stichocitele sunt cellule secretoare ce contin granule de diferite dimensiuni, cu morfologie variata. Aceste granule sunt secretate, si de intestinal in dezvoltare al viermelui si sunt puse in libertate la aproximativ 30 de ore de la imfestare, fiind puternic antigenice. Se pare ca aceste secretii ajuta viermele la invadarea tesuturilor si in nutritive. Stichocitele contin fosfataze acide si ATP-aza, dar nu si fosfataza alcanica.
Durata de viata este estimate intre 18-40 zile, in cazul infectiilor massive longevitatea parazitilor fiind mai mare. Se apreciaza ca dupa copulare masculii mor, fiind eliminate, femelele ramanand vii inca 4-6 saptamani. Copularea adultilor se face la 36-48 de ore dupa infectie iar la eliminarea primelor larve tinere are loc la 4-5 zile de la infectare la animale si 7 zile la om.
Spermatozoizii sunt lipsiti de flagel si contin 2-3 cromozomi, cu toate ca celelalte cellule somatic ale lor contin 5 cromozomi.
Ponta larvelor are loc din 30 in 30 de minute, iar prin viviparitate larvele eliminate de o femela pe parcursul vietii sale variaza intre 345-15000, cu o medie de 1500-2000 de larve, fiind subliniat faptul ca 70% din larve sunt depuse in primele 3 zile de la inceperea pontei, numarul larvelor obtinute de la femelele situate in douden fiind numeric crescute fata de cele care se gasesc in jejun si ileon.
Ponta are loc in grosimea mucoasei intestinale, mai ales la nivelul glandelor Lieberkuhn.
Fecunditatea femelelor este apreciata dupa numarul de larve muscular existente la 30 de zile de la infectare, iar indexul capacitatii de reproductive este reprezentat de raportul dintre numarul de larve L1 infestate ingerate si numarul de larve regasite in musculatura.
1.4.2 Faza de migrare
Se accepta existent a doua cai de migrare a larvelor:
Migrarea active prin care larvele tinere strabat peretele intestinal, trec in cavitatea abdominal si migreaza prin tesutul conjunctiv spre muschii striate (fenomen variabil pentru 5% din totalul larvelor nou-nascute) dupa Soule si colab;
Migrarea pasiva, in care larvele depuse in peretele intestinului pe cale limfatica si sangvina ajung in muschii striate. Considerata ca fiind cea mai obisnuita cale de diseminare ea se prezinta astfel:
a. Vasele limfatice eferente - ganglionii limfatici mezenterici - canalul toracic - vena cava craniala - atriul drept - ventriculul drept - artera pulmonara - pulmon - vena pulmonara - atriul stang - aorta - artere - capilare - tesut conjunctiv interfascicular (uscular) - fibra musculara;
b. Calea circulatiei sangvine entero-hepatica: vena porta - vena hepatocardiaca (sushepatica si vena cana) - atriul drept - etc.
c. Peretele intestinal - cavitatea peritoneala - canal toracic - vena cava cranial - atriul drept - s.a.m.d.
Migrarea larvelor nou-nascute are loc atat prin miscarile active proprii, cat si prin cele limfohemodinamica, pana ajung sa se localizeze in muschii striate ai noii gazed, dar indifferent de calea de migrare a larvelor, atriul drept reprezinta o etapa obligatory si invariabila.
Cu toate ca larvele tranziteaza prin cord, creier, ficat si alte organe, ele se localizeaza si se dezvolta numai in musculature striata. Patrunderea in vasele limfatice si sangvine la o ora de la nasterea larvelor tinere se apreciaza ca are loc prin traversarea laminei propria a vilozitatilor cu ajutorul stiletului. In timpul acestei faze, ficatul si pulmonul pot retine numeroase larve un timp oarecare (peste, ore la sobolan), organe in care produc, prin actiuni conjugate o gama larga de leziuni.
1.4.3 Faza de invazie musculara
Miscarile proprii ale larvelor si preluarea lor pe cale limfohemodinamica le asigura acestora posibilitatea de a ajunge in final in locurile de asezare definitiva, reprezentate de musculature striata. O data ajunse aici, cu ajutorul stiletului bucal, dar si cu participarea unor enzyme histolitice se insinueaza sub sarcolema, in lungul fibrelor muscular. Aparitia larvelor in musculature se poate petrece intre a sasea zi pana la a optsprezecea zi de infectie, si determina organismul astfel parazitat o stare de miozita acuta.
Larvele tinere care nu reusesc sa penetreze sarcolema fibrei muscular, ca si cele care se localizeaza in alte tesuturi si organe si nu reusesc sa se incapsuleze, mor si sunt distruse prin procesele de aparare locala a gazdelor.
In timpul circulatiei catre tesutul muscular striat, larvele nu suporta evenimente de dezvoltare, pastrandu-si lungimea intre 80-120 microni, iar grosimea intre 5-6 microni.
Larvele L1 ajunse in fibra muscular incep sa determine modificari morfopatologice concomitent cu dezvoltarea lor, asa incat, in a 10-a zi de la infectare lungimea lor este de 450 de micron si diametrul 17-20 microni, la 17 zile organelle de reproducere ajung la 204 microni, putandu-se deosebi sexele.
Soule (1991) arata ca dupa 24 de ore de la patrunderea larvei in celula muscular, volumul acestei se dubleaza (184). Celula muscular astfel parazitata isi pierde propria structura organizatorica, capatand proprietati de celula hranitoare. Cuticula larvei se ingroasa si numarul stichocitelor creste de la 1 la 50, diferentiindu-se in celule ce contin granule alfa (10-15) si cellule ce contin granule beta (35-40), fenomen precedat in a 9-a si a 10-a zi de sinteza si secretie de antigen.
In general se considera mai frecventa prezenta parazitului in musculature scheletica puternic irigata (muschii maseteri, muschii laringieni, muschii intercostali, muschii diafragmatici, muschii psoasi), larvele instalandu-se de regula la nivelul insertiilor musculare, la limita deci dintre muschi si tendon sau aponevroza, structure ce constituie bariere natural in calea migrarii larvelor.
1.4.3.1 Stadiul de spiralare
In a 17-a zi de la infectie larva se curbeaza sub form literei "U", devine rezistenta la digestia peptic si este infectanta. Dupa 30 de zile, larvele capata o lungime de 10000 de micron si o grosime de 35 de microni, procesul de spiralare avansand astfel incat intre a 21-a si a 35-a zi de la infectare, larva prezinta aspectul unui tirbuson si este rasucita de doua ori si jumatate.
Examenul trichineloscopic efectuat, depisteaza greu infectia, deoarece component muscular si larva nu au un contrast evident, larvele putand parasi fibra muscular si trece in zonele de suc de carne, unde pot produce miscari de rasucire si incolacire. Astfel de larve ingerate rezista actiunii sucului gastric, sunt capabile de dezvoltare si inmultire si sunt deci infectante, de unde si necesitatea de a se examina sucul de carne din jurul fragmentului de carne etalat.
1.4.3.2 Stadiul de incapsulare
La 28-30 de zile de la infectare, actiunea patogena a larvei la locul de cantonare este exprimata printr-un process inflamator subacut. Dispar striatiunile transversal ale fibrei musculare, iar tesutul conjunctiv interfibrilar se organizeaza in jurul larvei, provocand o izolare a acesteia de organismul gazdei, si constituind astfel chistul de Trichinella spiralis.
Chistul, delimitat net de tesuturile din jur este alcatuit dintr-o capsula omogena, hialina, sticlos translucida, lamelata si cu contur dublu, in interior existand o magma albuminoasa, cu resturi de sarcoplasma si nucleu ce provin din fibrele muscular degenerate. Acest process de inchistare se realizeaza in 3 - 7 saptamani, depinzand de factori legati de parazit, ca si de gazda.
Se apreciaza ca procesul de formare a capsule nu este unul de necroza al celulei muscular, ci dimpotriva aceasta devine mai bogata in structura ergoplasmatica, prin existent unui mare numar de nuclei care constituie o celula uriasa, in a carei plasma se gaseste larva. Pe fata interna a capsule se gaseste un tesut conjunctiv, care face legatura cu partile extreme ale larvei, concluzionandu-se ca nu se distinge nici larva nici celula parazitata.
Se mentioneaza existent unei retele de capilare in peretele chistului, considerandu-se ca aceasta retea vasculara face legatura intre larva si gazda.
Prin criosectiune se demonstreaza contrariul, si anume ca larva este situate intracitoplasmatic si este lipsita de membrama periferica. Unii autori arata ca in fibra muscular parazitata, activitatea metabolica creste, dispar miofilamentele si segmentatia, nucleul intra in amitoza, nucleoli se largesc, mitocondriile se multiplica si se dezvolta o membrane groasa ca reactive parenchimatoasa de iritatie.
Tot referitor la formarea capsule, se mentioneaza ca in primele faze dupa patrunderea larvelor in fibra muscular se constata inmultirea si activarea nucleilor sarcolomei, cu sporirea cantitativa de sarcoplasma si aglomerarea ei in jurul parazitului.
Initial, nuclei sunt mari, veziculosi si dispusi central, ulterior migreaza spre periferie, se aplatizeaza si se picnozeaza. Se constituie astfel in jurul larvei, de obicei spiralate, o capsula bistratificata, formata intern din sarcoplasma, iar extern din precolagen si ulterior din colagen. Pe masura ce procesul evolueaza in jurul capsule se instituie o reactive mezenchimala reprezentata in cazurile incipiente prin 1-2 randuri de histiocite, iar ulterior adevarate granuloame bogate in cellule gigante si epitelioide.
In concluzie, formarea capsule este rezultanta unei adaptari a fibrei muscular parazitate, la prezenta larvei de Trichinella spiralis. In lipsa capsule, larva nu ar putea supravietui, din cauza raspunsului inflamator foarte puternic care se creeaza in jurul sau.
Incapsularea larvelor face sa scada stimularea antigenic, iar nivelul de anticorpi scade progresiv, pana la a dispare in cateva luni sau ani, desi parazitul este inca viu. Incapsularea larvei, permite reducerea reactiilor peroxidazice, si supravietuirea larvei timp de ani, prin utilizarea resurselor energetice.
In timp, infiltratul inflamator din jurul chisturilor capata un tablou histologic de forma cronica 5 cu preponderenta formelor limfoide. Ulterior, se instaleaza procese de scleroza cu zone variabile de hialinizari, iar infiltratul patrunde in capsula si conduce la liza parazitului.
Chistul de Trichinella spiralis are forma clasica ovalar alungita, cu cate un tubercul la cei doi poli, conferindu-i aspect de lamaie. Marele ax este dispus invariabil in lungul fibrelor musculare. Lungimea chistului este cuprinsa intre 400 - 700 microni, iar latimea, la mijlocul chistului este cuprinsa intre 250 - 300 microni.
Dimensiunile si forma chisturilor variaza in fnctie de regiunea anatomica in care se afla muschiul parazitat, precum si de specia care este gazda.
Legat de modul lor de grupare, se pot decela chisturi isolate, alaturate sau dispuse in siraguri, mai numeroase in apropierea tendoanelor si aponevrozelor musculare.
De subliniat este faptul ca in acest stadiu larvele de Trichinella spiralis nu au cea mai mare capacitate infectanta. Atat la om cat si la animale, infestarea se poate realize cu doze mici de larve: 50 - 75 larve pentru om, 40 - 50 larve pentru porc, 15 - 20 larve pentru carnasiere, 2 - 3 larve pentru un cobia. Luand in discutie numarul chisturilor in masa musculara a unui animal parazitat, s-au constatat variatii mari ce depind si de zona musculara cercetata.
1.4.3.3 Stadiul de calcificare
Calcificarea chisturilor de Trichinella spiralis este un proces specific granuloamelor parazitate si consta in depunerea sarurilor de calciu pe chistul cu larva. Calcificarea chisturilor de Trichinella spiralis este un fenomen tipic, in sensul ca depunerile de saruri de calciu incep la poli si continua spre central chistului, fiind regresiv in timp comstatandu-se decalcificarea chisturilor si apoi resorbtia acestora. Dupa Gh. Olteanu (134) calcificarea incepe la 5 - 6 luni, dupa 7 luni este de 1%, dupa 12 luni 30%, iar dupa 18 - 24 luni este complete.
Etapele procesului de calcificare au mai multe substadii cu importanta in diagnosticul trichineloscopic al carnurilor (Cristescu si Popa (1, 63)) astfel dupa stadiul de calcificare, chisturile de Trichinella spiralis se pot imparti dupa cum urmeaza :
a. Chisturi cu inceput de calcificare. Acestea prezinta :
- capsula cu depuneri de saruri de calciu la poli; aceasta este transparenta neomogen, sau este tulbure la poli;
- larva, bine vizibila este vie si infectanta.
b. Chisturile in plin proces de calcificare prezinta^
capsula cu depuneri reduse de saruri de calciu pe intreaga suprafata, fund cu predominanta tulbure la poli.
- larva, mai putin vizibila, este vie si infectanta.
c. Chisturile calcificate partial prezinta :
- capsula cu intense si uniforme depuneri de calciu este partial tr ansparenta spre opacitate uniforma;
- larva, foarte greu sau deloc vizibila, este practic vie si infestanta.
- capsula ultratransparenta, opaca, puternic calcificata;
larva, calcificata, vizibila numai dupa clarificare, este impregnata cu saruri de calciu, pare moarta, fiind practic neinfectanta.
d. Chisturile complet calcificate au:
- capsula ultratransparenta, opaca, puternic calcificata
- larva, calcificata, vizibila numai dupa calcificare, este impregnata cu saruri de calciu, pare moarta, fiind practic neinfestanta.
e. Chisturile calcificate atipic
- apar relativ rar, larvele calcificandu-se fara calcificare capsulei;
- la examenul trichineloscopic apar doar fragmente de larva, sau resturi tio tesut conjunctiv;
- in toate situatiile larvele sunt pe moarte.
f. Chisturile calcificate patologic
- apar in mod exceptional;
procesul de calcificare incepe de la centru spre poli (ca in cazul chisturilor de Sarcocystis
- dupa clarificare se poate vedea larva sau fragmente din aceasta;
- se observa lipsa striatiunilor transversale din fibrele musculare;
- la poli sunt prezente inca celule adipoase.
g. Chisturile decalcifiate, pe cale de resorbtie, prezinta:
capsula opaca, usor transparenta, cu depuneri de calciu in parte resorbit;
- larva , (de cele mai multe ori resturi), cu impregnatie de saruri de calciu, mai mult sau mai putin evidenta si care se afla pe cale de resorbtie, deci moarta.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 4249
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved