| CATEGORII DOCUMENTE |
| Alimentatie nutritie | Asistenta sociala | Cosmetica frumusete | Logopedie | Retete culinare | Sport |
IMUNITATEA ANTIVIRALA
Avand un genom extrem de simplu (intre 3 si 250 gene) virusurile nu sunt capabile sa elaboreze un numar de proteine si enzime importante (de ex. enzime necesare catabolizarii glucidelor sau enzime necesare replicarii acizilor nucleici). In consecinta, ele sunt obligate sa paraziteze diverse gazde celulare, pentru ca folosind 'masinaria biochimica' a acestora, sa poata supravietui si prolifera. Un numar de virusuri care patrund in diverse celule umane sau animale produc tulburari patologice cu un spectru clinic foarte variat: boala acuta, subclinica, recurenta, latenta sau cronica.
Raspunsul organismului gazda este si el foarte divers implicand toate componentele nespecifice ale apararii, dar cu o nuanta proprie (Fig.25): cu preponderenta imunitatii celulare legata de dezvoltarea intracelulara a virusurilor si relativa ineficienta a raspunsului umoral.
Eficienta globala a raspunsurilor poate fi deplina (ducand la eliminarea virusurilor si stabilirea unei imunizari durabile) sau partiala cu persistenta virusului in organism.
In fine, in anumite situatii insusi mecanismul imun de aparare poate avea efecte adverse, producand manifestari patologice.
In forma sa obisnuita, infectia virala incepe cu invazia unuia dintre invelisurile epiteliale de granita: tegumente, mucoase respiratorii si digestive. Unele virusuri raman cantonate la acest nivel. Altele, se raspandesc in organism pe cale sanguina (faza viremica a infectiei) si se stabilesc in organele-tinta. Pe parcursul acestui traseu aceste virusuri expun Atg. proprii si amorseaza mecanismele de aparare nespecifica impreuna cu initierea raspunsului imun umoral. In cazul infectiei primare, Atc. virali se elaboreaza insa prea tarziu pentru a putea fi operativi.
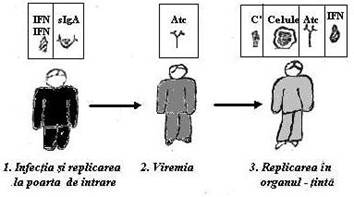
Ei vor juca un rol mai activ in infectiile secundare sau in interactiunea cu celulele care exprima Atg. virale.
La comanda stimulilor virali celulele gazda pot fabrica proteine. Drept urmare, in celulele parazitate coexista atat Atg. virale codificate de genomul viral cat si Atg. celulare codificate de gene celulare activate de virus. Toata aceasta zestre de Atg., apare risipita in nucleu, in citoplasma sau pe suprafata celulei gazda. Atg. de suprafata codificate de celula gazda au potential redus de a induce raspunsul imun. In schimb, servesc clinicianului ca markeri care dezvaluie persistenta infectiei. Atg. virale exprimate pe suprafata celulei sunt insa imunogene. Raspunsul declansat de aceste Atg. este T-dependent. (Fig.43)
Mecanismele de aparare umorala
Mecanismele de aparare umorala au rolul sa neutralizeze si sa inactiveze virusurile. Inhibitia nemijlocita a infectivitatii virale prin interactiunea anticorpilor cu antigenele virale de suprafata, este un mecanism esential de aparare a gazdei. Anticorpii previn absorbtia si penetrarea virusului `n celula gazda, iar ulterior difuzarea infectiei este limitata tot de catre anticorpi.
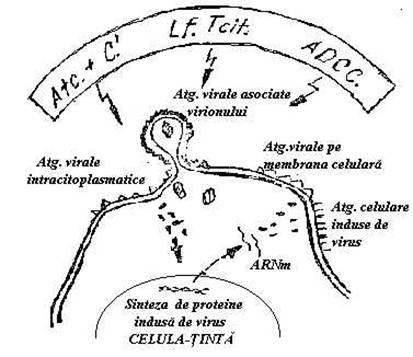
Fig. 43 Exprimarea Atg virale pe suprafata membranei celulare si recunoasterea lor de efectorii imunitatii umorale si celulare
Anticorpii antivirali sunt capabili sa se lege direct de virusuri in faza extracelulara sau pe Atg. virale exprimate pe celule. IgM si IgG intercepteaza virusurile vehiculate in plasma si fluidele tisulare, in timp ce IgA din secretii protejeaza suprafetele mucoaselor portilor de intrare. Legarea pe virusurile extracelulare cu efectul sau protectiv antiviral reprezinta baza seroterapiei profilactice.
Prin atasarea pe virusurile extracelulare, Atc. pot neutraliza capacitatea infectanta a acestora deoarece ar impiedica patrunderea lor in celule. Complementul si interferonul actioneaza sinergic cu Atc. pentru a preveni infestarea celulara.
Complementul poate sprijini actiunea neutralizanta a Atc. prin invelirea virusurilor sau prin provocarea lizei unei particule virale.
Sistemul complement activat este capabil sa medieze o serie de activitati biologice, dintre care cea mai cunoscuta este capacitatea de a media distrugerea litica a virusurilor cu `nvelis lipidic. Mecanismul este probabil legat de capacitatea sistemului de a produce un raspuns inflamator acut, care are drept scop, sa localizeze substanta care a activat complementul.
Virusurile si celulele infectate cu virusuri, `nvelite cu anticorpi, se comporta ca si complexe imune tipice, activand complementul. Mecanismele de inactivare pot fi litice si nelitice.
Mecanismele litice intervin cand sistemul complement este activat de virusuri cu `nvelisuri, care au lipide celulare `nglobate `n aceasta structura. Cercetarile de microscopie electronica au demonstrat ca modificarile structurale rezultate din actiunea sistemului "C" pot fi de la distrugeri minore, pana la pierderea integritatii virale. Aceste modificari de permeabilitate, permit liza osmotica sau influxul de proteine plasmatice si nucleaze `n interiorul virusului. Modificari mai extinse duc la pierderea `nvelisului si la eliberarea acidului nucleic viral. Dintre virusurile ce sunt sensibile la liza mediata de sistemul "C" amintim pe cele din genurile Retrovirus, Myxovirus, Paramyxovirus si Alfavirus.
Mecanismele nelitice au un rol important `n activarea virusului prin acumularea de proteine ale sistemului complement pe `nvelisul viral, care pot interfera cu adsorbtia sau cu penetrarea virusului `n celula, reducand astfel infectivitatea. S-a demosntrat ca `ndeosebi fractiunile complementului cu un mare numar de molecule (C1q, C4b, C3b) mascheaza situsurile necesare pentru atasarea virusului la suprafata celulelor potential sansibile. Un alt mecanism prin care sistemul complement incativeaza virusurile este agregarea. Un astfel de mecanisam este realizat, `ndeosebi, de componenta C3b opentru care virusul poseda receptori localizati la nivelul infectiei virale.
Interferonii. Mai multe specii moleculare de IFN (alfa si beta) sunt secretate de celulele infestate de virus, foarte rapid dupa debutul infectiei inca inaintea aparitiei Atc. Combinarea acestor molecule de IFN cu membrana celulelor neinfectate din vecinatate initiaza traducerea unui ARNm care codifica o proteina antivirala care impiedica invadarea acestor celule, iar monocitele si macrofagele dobandesc sub influenta IFN capacitatea de a se impotrivi replicarii virusurilor. In acest mod IFN contribuie la eliminarea focarelor primare ale infectiei. Mai tarziu, in cursul infectiei dupa stimularea specifica a LfT, apare IFN gama produs de acestea, care va amplifica mai multe modele de reactii citotoxice indreptate impotriva celulelor infecatate, reactii efectuate de LfT cit., celulele NK si K, Mcf. activate.
Atasarea Atc. pe Atg. exprimate pe celula infectata poate produce efecte diferite de la un tip de infectie la altul:
activarea complementului pe celula-tinta si citoliza;
atragerea si activarea celulelor K urmate de efect citotoxic indirect ('ADCC');
modularea Atg. virale sau despuierea celulelor de Atg. ar permite celulei infectate sa evite distrugerea prin mecanisme citotoxice.
ATC+C'- LIZA VIRUSURILOR - NEUTRALIZARE
CU LIPIDE MEMBRANARE
ATC+C' - LIZA CELULELOR INFECTATE -POATE DEZGOLI (CALEA VIRUSUL CU ALTERNA) PROTEJARE
CELULELOR
Schema nr. 4 Efectele antivirale ale anticorpilor
Mecanismele de aparare celulara
In erarhia formelor de aparare antivirala mecanismele imunitatii celulare joaca un rolul dominant. (Fig.44)
Imunitatea mediata celular joaca un rol important `n rezistenta fata de anumite infectii virale. Ca regula generala, virusurile care se asambleaza la nivelul celulelor infectate sau care determina `n cursul replicarii neoantigene `n zona periferica a celulei, induc imunitatea celulara.
Mecanismul efector al imunitatii celulare urmeaza un model simplu, care asigura si participarea mecanismelor nespecifice, ca auxiliar indispensabil. Limfocitele sensibilizate lansate `n circulatie, devin capabile sa sintetizeze o gama larga de produse biologice active, `n momentul contactului cu celulele infectate, tumorale sau celulele alogrefelor. Asemenea celule sunt denumite celule tinta, ele purtand pe membrana lor antigene specifice, `n raport cu care limfocitele cu fost sensibilizate.
Atg. virale exprimate pe suprafata celulelor infectate impreuna cu Atg.CMH de clasa I stimuleaza LfT cit. specifice care vor produce liza celulei infestate. Raspunsul este insa mult mai amplificat prin stimularea concomitenta si a LfT CD4+ care induce prin intermediul secretiei de citokine urmatoarele efecte:
a) recrutarea si activarea Mcf. (care dobandesc capacitati citotoxice nespecifice asupra celulelor infectate);
b) activarea si cooperarea cu LfB ( care primesc semnale helper de recunoastere a Atg. virale si produc Atc. specifici fata de acestea ca si anticorpi care pot interactiona cu particule virale eliberate din celula gazda distrusa prevenind pe aceasta cale patrunderea virusurilor in alte celule);
c) activarea LfT cit.;
d) activarea mediata a IfN si a celulelor NK.
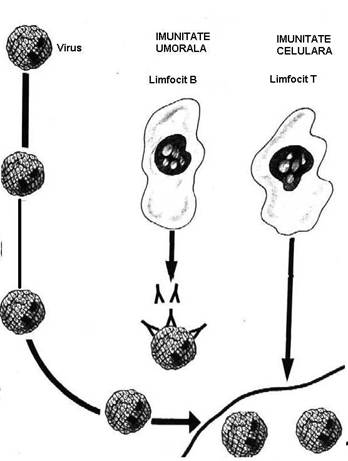
Fig. 44. Raspunsul
imun `n imunitatea antivirala
Nu intotdeauna mecanismele de aparare reusesc sa elimine complet virusul infectant care poate persista luni (sau uneori) ani de zile. Se accepta trei forme de infectii persistente: cronice, latente si lente.
infectii cronice ( modelele: infectia cu virus citomegalic, hepatita B) se caracterizeaza prin persistenta virusului in ciuda unei reactii imune destul de ample, care insa nu influenteaza evolutia cronica a bolii. Serul bolnavilor contine titruri ridicate de Atc. IgM specifici, dar imunitatea de tip celular pare serios afectata asa cum o demonstreaza scaderea raspunsurilor, in vitro, a limfocitelor T in diverse modele de stimulare. Mecanismele acestei stari de imunodeficienta nu sunt bine cunoscute.
infectiile latente (modelele: virusuri herpetice-herpes simplex I si II, varicella-zoster) se caracterizeaza clinic prin secventa: boala acuta, remisiune (vindecare aparenta), recadere, remisiune In remisiunea clinica virusul persista de fapt, in celulele organului-tinta. Exista o productie de Atc. IgG serici care insa nu confera protectie deoarece virusurile sunt localizate strict intracelular. Raspunsul celular controleaza infectia si o mentine in stare latenta dar nu o poate eradica. Recaderile sunt posibile daca apare o deficienta a controlului infectiei: unele medicamente, stari pasagere de imunosupresie sau bolile limfoproliferative cronice pot facilita recrudescenta puseelor de boala;
infectiile lente au evolutie incentinita atat in incubatie cat si in cursul bolii manifestate. Un astfel de exemplu este leucoencefalopatia multifocala (produsa de un virus extrem de raspandit in randul populatiei umane de toate varstele, denumit SV 40). Boala se dezvolta ca o infectie oportunista pe terenul unei imunodeficiente de lunga durata care insoteste bolile limfoproliferative cronice (limfoame maligne de ex.) sau imunosupresia terapeutica aplicata la purtatorii de grefa pentru prevenirea fenomenului de respingere.
In anumite imprejurari, raspunsul imun antiviral provoaca gazdei alterari patologice. Mecanismele T-dependente provoaca uneori leziuni severe si progresive celulelor infectate. Infestarea experimentala a soarecilor cu virusul coriomeningitei limfocitare produc moartea animalelor in urma unor leziuni cerebrale inversibile. Supresia cu Ciclofosfamida (CFA) a animalelor inoculate previne moartea acestora, dar duce la o infectie persistenta. Efectul produs de CFA poate fi anihilat dupa administrarea de LfT recoltate de la un soarece imunizat in prealabil, sugerand ca moartea animalelor restaurate se datoreste efectului citopatic al LfT. Un mecanism similar a fost incriminat in hepatita cronica activa umana: Lf citotoxice recunosc Atg. exprimate pe suprafata hepatocitelor parazitate si produc o hepatocitoliza persistenta.
Anumite virusuri infecteaza celulele limfoide cu efecte extrem de severe. Virusul HTLV I ('human T leukemia/lymphoma virus I') infecteaza LfT CD4+ si produce o proliferare continua a acestora ('imortalizare') a carei urmare este aparitia unui limfom malign foarte grav (limfomul T endemic). Aceleasi limfocite T CD4+ sunt tinta predilecta a virusului HIV care are efect citopatic: distrugerea progresiva a zestrei T CD4+ este unul din mecanismele de baza care explica instalarea sindromului imunodeficientei dobandite (AIDS sau SIDA).(Fig.45)
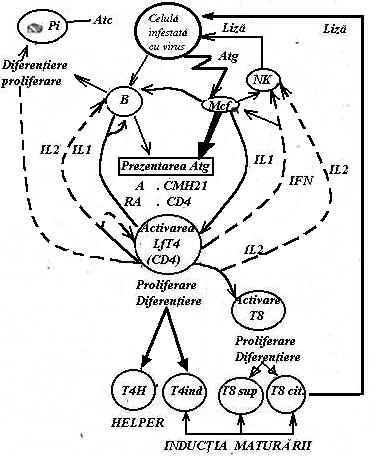 |
B = LfB; Mcf = macrofage; NK = celule 'natural killer'; T4h = LfT4 helper; T4ind = LfT inductor; T8sup= LfT8 supresor; T8cit = LfTh citotoxic; IL1-IL2= interleukina 1 si 2; IFN = interferon; FAB,FPB si FDB = factori de activare, proliferare si diferentiere ai LfB; Atg = antigen; RAtg = receptorul pentru Atg. al LfT; CMH = proteine de clasa II-a din complexul major de histocompatibilitate; CD4 = markerul LfT4
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 2839
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2026 . All rights reserved