| CATEGORII DOCUMENTE |
| Alimentatie nutritie | Asistenta sociala | Cosmetica frumusete | Logopedie | Retete culinare | Sport |
MECANISMELE RUPERII TOLERANTEI
Raspunsurile autoimune par a fi identice in derularea lor cu raspunsurile dirijate impotriva Atg.conventinale. Din acest punct de vedere pot fi discutate:
1. Recunoasterea auto-Atg. Asa cum am ilustrat in exemplul referitor la rasa de pui obezi, auto-Atg. nu sunt 'anormale' la subiectii cu tulburari autoimune. Mai mult, in mod normal prezenta auto-Atg. este necesara pentru mentinerea tolerantei fata de self. Afirmatia se sprijina pe exemplul asa-ziselor Atg sechestrate: Atg. cristalinului si Atg. spermatozoizilor (aceste Atg. sunt izolate prin bariere anatomice de restul organismului: eliberarea lor in organism produce imediat o reactie imuna). Majoritatea auto-Atg. 'coabiteaza pasnic' cu celulele imunologic competente si probabil sunt de tip timo-dependent, adica solicita pentru a declansa un raspuns prezentarea lor de catre Mcf. si cooperarea dintre celulele T auxiliare si LfB autoreactive specifice. Mecanismul implica obligatoriu o prezentare restrictiva impreuna cu moleculele CMH de clasa a II-a.
Importanta moleculelor CMH de clasa II-a in geneza BAI este sugerata de doua fapte de observatie:
exprimarea aberanta a acestor Atg. pe tesuturi care in mod normal nu le exprima. Aceasta exprimare ar putea fi indusa de unele infectii virale (prin intermediul sintezei de IFN) si ar putea explica aparitia unor BAI.
prevenirea inducerii unor BAI experimentale (ex. encefalomielita sau miastenia gravis) prin injectarea in vivo de Atc. anti-Ia (anti CMH DR). Unii agenti infectiosi pot prezenta determinanti comuni cu epitopi ai auto-Atg. Infectiile respective provoaca un raspuns autoimun printr-o reactivitate incrucisata. De exemplu, unele tulpini de streptococ hemolitic au determinanti comuni cu miocardul, cu endocardul sau cu celulele cerebrale. Pe aceste baze se explica atingerea cardiaca sau de choree care se observa in reumatismul acut streptococic.
2. Activarea Lf. Existenta clonelor autoreactive este in prezent unanim acceptata. Circa 1% dintre Lf normale circulante prezinta molecule de tireoglobulina atasate pe receptorii lor specifici fara sa existe semne de agresiune autoimuna. Mai mult, s-au decelat in circulatia subiectilor normali diversi auto-Atc. 'naturali'. Aceste observatii, culese dintre multe altele asemanatoare, sugereaza existenta LfB autoreactive dar nefunctionale la indivizii normali. Raportata la ansamblul 'masinariei' imune aceasta 'inertie' a LfB autoreactive ar implica inhibitia globala a mecanismului de activare a LfB, inhibitie care intereseaza in special LfTh si care ar fi controlata de LfTs.
Activarea sistemului (reactie autoimuna) ar putea surveni in urmatoarele imprejurari:
suprimarea supresiei: administrarea Ciclofosfamidei ( inhibitor selectiv al diferentierii LfTs) la soarecii din suse nativ-rezistente la inducerea de BAI experimentale, permite obtinerea unei tiroidite alergice;
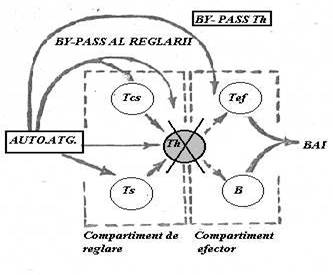
ocolirea (by -pas-ul) supresiei
(Fig.61)
Fig. 61 Ipoteza lui I.M.Roitt (1985) prin care considera ca ruperea tolerantei poate aparea prin ocolirea mecanismelor supresoare de reglare
activarea directa a LfT;
activarea directa a LfB si T efectoare;
activarea LfT contrasupresoare;
3. Dereglarea productiei de AAC. AAC care se asociaza cu BAI sunt extrem de heterogeni: IgM, IgG, IgA sau chiar IgE. Ei poseda diverse idiotipuri publice si private ca diferente intre speciile de molecule ale unei clase de Ig care sunt plasate in regiunile variabile - Fab - ale situsului combinativ.
Controlul sintezei de AAC este exercitat de LfT auxiliare si de LfTs iar dereglarile acestui control au fost amintite in sectiunea precedenta. Reamintind 2 elemente fundamentale (a)sIg- receptorul pentru Atg. al LfB -are aceeasi compozitie moleculara in portiunea variabila cu Atc. produsi de celula respectiva si (b) receptorii pentru Atg. ai LfB si T prezinta portiuni variabile, precizand ca atat receptorii celulelor limfoide autoreactive cat si AAC exprima determinanti idiotipici privati (proprii fiecarei clone de AAC) si publici ( comuni mai multor clone de molecule care reactioneaza cu aceleasi auto-Atg). Aceste idiotipuri reprezinta de fapt o varietate imensa de autoantigene care ar putea constitui motivele unei autoimunizari. Recent s-a demonstrat existenta unor idiotipuri comune intre Atc. anti-Klebsiella si auto-Atc. anti-ADN. Aceasta duce la presupunerea posibilitatii instalarii unei reactii incrucisate intre idiotipul unui Atc. specific pentru un Atg. exogen si un determinant corespunzator din structura unui auto-Atg.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 2204
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved