| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Glicoliza
Metabolismul lactozei sub actiunea bacteriilor lactice
Aproximativ 98% din continutul de lactoza din lapte trece in zer sub forma de lactoza sau acid lactic, iar partea retinuta in branzeturi este metabolizata pana la lactat, partial in timpul fabricarii branzeturilor si partial in timpul primelor stadii de maturare, in mod obisnuit de catre culturile starter. Acidul lactic aduce o contributie importanta la formarea aromei branzeturilor cu coagularea acida si contribuie semnificativ la formarea aromei branzeturilor tinere coagulate enzimatic.
Lactococcus lactis ssp. lactis si Lactococcus lactis ssp. cremoris metabolizeaza lactoza la acid lactic L(+): jumatatea reprezentand glucoza este metabolizata pe calea Embden-Meyerhof, in timp ce galactoza este metabolizata pe calea tagatozei. In branzeturile in care se utilizeaza culturi starter termofile, lactoza este consumata de Streptococcus termophillus (care se dezvolta primul, pe masura ce casul se raceste) si este hidrolizata de β-galactozidaza la glucoza si galactoza. Glucoza este metabolizata pe calea Embden-Meyerhof la L-lactat, Streptococcus termophillus nefiind capabil sa metabolizeze decat galactoza pe care o produce. Atunci cand casul este suficient de rece se dezvolta Lactobacillus. Lactobacilii galactozo-pozitivi metabolizeaza galactoza pe calea Leloir pana la glucozo-6-fosfat, care este apoi metabolizata la D,L -lactat pe calea Embden-Meyerhof. Oricum multe tulpini de Lactobacillus delbrueckii ssp lactis si Lactobacillus delbrueckii ssp bulgaricus sunt galactozo negativi.
Culturile starter utilizate pentru unele branzeturi includ specii de Leuconostoc care izomerizeaza galactoza proveita din hidroliza lactozei, via Leloir, la glucozo-6-fosfat si apoi metabolizeaza si glucoza via calea fosfocetolaza la acid lactic, etanol si CO2.
Metabolismul acidului lactic in branza
Concentratia lactatului in branzeturile tip Camembert, Elvetiana si Cheddar este de aproximativ 1, 1,4 si respectiv 1,5% (Fox, 1990). Metabolizarea acidului lactic depinde de sortiment.
In branza Cheddar, Olanda si sortimente similare, L-lactatul este izomerizat la D-lactat de bacteriile lactice care nu provin din cultura starter (BLNS), Thomas si Crow (1983).
Racemizarea L-lactatului nu este semnificativa din punct de vedere al aromei, dar D-lactatul poate avea consecinte nutritionale nedorite la inceput si D-lactatul de calciu, care este mai putin solubil decat L-lactatul de calciu poate cristaliza in branza, cauzand pete albe, nedorite, in special pe suprafetele taiate (Fox, 1990).
Lactatul poate fi oxidat de BLNS in branza Cheddar si probabil in branzeturile similare la acetat si CO2 (fig.16
Aceasta reactie depinde de oxigenul disponibil, care este determinat de dimensiunea branzei si de permeabilitatea la oxigen a materialului de ambalaj (Thomas,1987). Acetatul care poate fi produs si de catre bacteriile starter, din lactoza (Thomas,1985), sau citrat, sau din aminoacizi de catre bacteriile starter si lactobacili (Nake, Elliott, 1965) este prezent in concentratii destul de mari in branza Cheddar si se considera ca participa la aroma sa, desi concentratii mari pot determina lipsa aromei ( Astor si Dulley, 1982). De aceea oxidarea lactatul la acetat poate contribui la aroma branzei Cheddar.
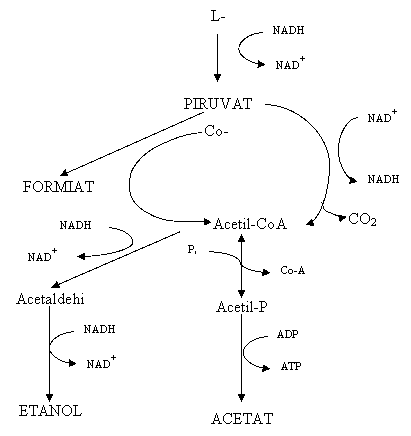
Fig. 8 Transformarile lactatului in prezenta BLNS
Fermentarea lactozei si a lactatului in branzeturile Elvetiene au fost descrise de Turner (1983). De obicei branza Emmental contine aproximativ 1,2% amestec D si L-lactat, la 14 zile, timp in care zaharurile au fost metabolizate complet. La transferul in incaperi calde se dezvolta propionibacteriile care metabolizeaza preferential L-lactatul la propionat, acetat si CO2.
CO2 este responsabil de formarea ochiurilor de fermentare, o trasatura caracteristica a acestor sortimente de branzeturi, acetatul si in principal propionatul contribuind la aroma.
Dintre toate sortimentele de branza, metabolismul lactatului este probabil cel mai extensiv in sortimentele maturate, cu mucegai (Camembert, Brie). Dupa prelucrare, aceste branzeturi contin 1% acid lactic produs in principal, sau exclusiv de mezofilii starter. Organismele secundare colonizeaza rapid si domina la suprafata acestor branzeturi- la inceput Geotrichum candidum si drojdii, urmate de Penicillium caseicolum si la fabricarea traditionala, de Brevibacterium linens .
Geotrichum candidum si Penicillium caseicolum metabolizeaza rapid lactatul la dioxid de carbon si apa, producand o crestere a pH-ului. Brevibacterium linens nu se dezvolta la pH mai mic de 5,8 si nu colonizeaza la suprafata branzeturilor, pana cand pH-ul nu are o valoare ridicata. Dezacidifierea apare la inceput la suprafata, rezultand un gradient de pH de la suprafata spre centru, ceea ce determina o difuzie a lactatului in exterior. Cand lactatul a fost epuizat de Penicillium caseicolum , acesta metabolizeaza proteinele producand amoniac, care difuzeza in interior, favorizand cresterea pH-ului.
Concentrarea fostatului de calciu la supafata creste solubilitatea la un pH mai mare si duce la aparitia precipitatului sub forma de stat, determinand aparitia unui gradient de concentratie al Ca3 (PO4)2 in masa branzei. Reducerea concentratiei Ca 3(PO4)2 in interior ajuta la inmuierea branzei.
Cresterea pH-ului favorizeaza actiunea plasminei, care impreuna cu enzimele coagulante reziduale sunt responsabile de proteoliza in aceasta branza mai mult decat proteinazele secretate de microorganismele de suprafata, care, desi sunt foarte active difuzeaza in branza la un nivel foarte redus, cu toate ca produsii actiunii lor specifice la suprafata branzeturilor pot difuza in branza. Actiunea combinata a cresterii pH-ului, pierderea Ca si proteoliza este necesara pentru procesele de inmuiere (catifelare) a branzeturilor Brie si Camembert.
Metabolismul lactatului si cresterea pH-ului influenteaza aroma acestor branzeturi, care au preponderent gust de crema. Metabolizarea lactatului poate fi realizata de drojdiile prezente la suprafata branzeturilor grase maturate. Desi acest proces are un slab efect direct asupra aromei, cresterea pH-ului este esentiala pentru dezvoltarea lui Brevibacterium linens, produsii de metabolism ai acestei bacterii dominand aroma acestor branzeturi.
Un defect obisnuit ce poate apare din metabolismul lactatului se datoreaza producerii de butirat din lactat sau glucoza de catre specii de Clostridium (fig.9).
Aceasta reactie duce la balonarea tarzie si la lipsa aromei in multe sortimente de branzeturi, cu exceptia cazurilor cand se iau masuri de precautie (GHP, adaugare de NO3-, lizozim, bactofugare sau microfiltrare).
In termeni cantitativi aceste transformari sunt printre pricipalele procese metabolice. Totusi, in comparatie cu alte transformari biochimice care au loc in timpul maturarii branzeturilor, transformarea lactozei in lactat are un efect relativ salb asupra aromei branzei mature, dar din moment ce ii determina pH-ul are o importanta majora in reglarea altor functii biochimice care apar in timpul maturarii.
Izomerizerea lactatului are un efect redus asupra aromei, dar transformarea in propionat si/sau acetat este probail semnificativa daca apare, metabolismul lactatului la butirat avand un efect negativ asupra calitatii branzei.
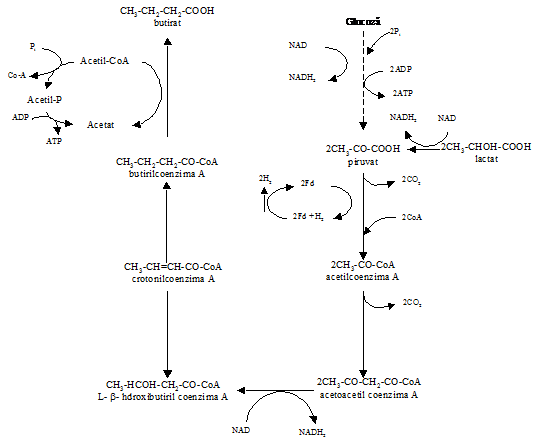
Fig. 9 Calea formarii butiratului din glucoza sau lactat
Metabolismul citratului
Concentratia relativ redusa de citrat in lapte, aproximativ 8 mM poate induce in eroare in ceea ce priveste metabolismul citratului pentru multe sortimete de branzeturi obtinute prin utilizarea culturilor mezofile (Cogan,1985, Cogan, Hill, 1993). Aproximativ 90% din citratul din lapte este solubil si trece in zer. Totusi concentratia citratului in faza apoasa a branzei este de 3 ori mai mare decat cea din zer (Fryer, 1970), reflectand concentratia citratului coloidal.
Citratul nu este metabolizat de Lactococcus lactis sau Lactococcus cremoris, dar este metabolizat de Lactococcus lactic ssp. lactis biovar diacetylactis si specii de Leuconostoc cu producere de diacetil si dioxid de carbon. Citratul nu este metabolizat de Streptococcus termophillus sau de lactobacilii termofili (Hickey, 1983), dar multe specii de lactobacili mezofili metabolizeaza citratul cu producere de diacetil si formiat (Fryer, 1970).
Producerea de dioxid de carbon din citrat este responsabila de formarea ochiurilor de fermentare in branza Olanda si de deschiderile nedorite si de casul moale in branza Cheddar si Cottage. Diacetilul este foarte important in formarea aromei branzei Cottage si Quarg si contribuie la formarea aromei branzei Olanda. Branza Cheddar contine 0,2-0,5% vol/vol. citrat care este metabolizat in funtie de microflora BLNS. Diacetilul apare intr-o concentratie ridicata in branza Cheddar si reprezinta un constituent important al aromei. Acetatul format din citrat poate de asemenea contribui la aroma branzei. Diacetilul poate fi transformat in acetoina, 2,3-butilen glicol si 2-butanona.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 2682
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved