| CATEGORII DOCUMENTE |
| Bulgara | Ceha slovaca | Croata | Engleza | Estona | Finlandeza | Franceza |
| Germana | Italiana | Letona | Lituaniana | Maghiara | Olandeza | Poloneza |
| Sarba | Slovena | Spaniola | Suedeza | Turca | Ucraineana |
DOCUMENTE SIMILARE |
|||||
|
|||||
TERMENI importanti pentru acest document |
|||||
| : | |||||
Paleontological
Institute RAS, Profsoyuznaya 123,
Bacterial paleontology is rather young branch of paleontological studies. Bacteria (and microbes in general) could be perfectly preserved as fossils. The major part of the sedimentary rocks formed in the photic zone of epicontinental basins of the past originated under influence of microorganisms. After special studies the number of minerals formed with the participation of microorganisms increase and already is above a hundred. Bacterial-paleontological data accompanied by the data on the first origin of eukariots, metazoa, etc. significantly enrich our knowledge on biosphere evolution, and reveal the long period of transitional biosphere prior to the appearance of typical eukaryotic biosphere of modern type. The bacterial-paleontological data on carbon chondrites make foundation for the possibility of presence of extraterrestrial bacterial life.
Key words: bacterial paleontology, mineralogy, sedimentation, evolution of Biosphere.
The important role of bacteria and other microbes in the geological processes was suggested long ago. Already in the end of the XIXth century N.I. Andrusov in his lectures spoke about the bacterial role in origin of some Cenozoic deposits of iron and sulfur. Later numerous attempts of microscopic study were aimed to prove the presence of fossilized bacteria in rocks. On of the most prominent achievement of that period were A.G. Vologdins report and publication [37].
However, the inexpressive picture obtained by specialists using relatively small magnification could not convince the scientific public. Moreover, many specialists of that period claimed the tremendous role of bacteria in the formation of the sedimentary minerals resources, like bauxite, iron ores, etc. They were incurred a fierce criticism or even derision by the leading scientists of that time.
The significant breakthrough of the post-war science was discovering of fossil microorganisms in cherty rocks. A pioneering work in this field belong to T. Barghoorn and his students [5]. His studies was developed in a variety of countries [16, 28, 29, 30, 31, 32]. A vast material have been collected permitting to reconstruct not only the origin and development of microorganisms on the Earth [27, 30, 33], but also the general pattern of biosphere evolution in the Precambrian. These studies also considered the data obtained by the traditional method of maceration commonly used for extraction of acritarches, spores and pollen. In such cases chiefly the forms with the organic shells were extracted.
However, the most significant breakthrough was made after introducing of the electron microscopy to the study of microbial remains. New technology revealed that almost all sedimentary rocks contain finely preserved bacteria [14, 20, 21, 22, 23, 26, 45, 46, 50]. The perfect preservation could not be ensured without high rates of fossilization [10, 13, 20, 21, 23, 35, 40], and it was proved experimentally.
The intensive study of fossil bacteria - bacterial paleontology [23] starts and goes on during the last decade. From one hand, it is tightly connected with the extensive investigation in the field of geomicrobiology, science which studies the interrelation of recent microbial biota with rocks, minerals and other materials [1, 4]. From another hand, it is greatly influenced by the study of cyanobacterial mats (CBM), the recent and fossil as well [12, 21, 43]. These studies convinced us in the extreme antiquity of CBM and their highly important role in the formation of oxygenic atmosphere and whole biosphere in general. Here we should mention the significance of the studies of bacteria in the extreme environments - perhaps part of such conditions are similar to those present in the Archean.
The main task for bacterial paleontology is to study the fossil microbes. The traditional paleontology deals with remains of ancient organisms. It creates systematics based on organisms morphological features considering their phylogenetic relations. The main users of paleontology are the evolutionary theory, the history of organic world and the biostratigraphy. Recently the paleontology brought us priceless data for study of the biosphere evolution. Paleogeography also gains significant profit from the paleontology.
Bacterial paleontology is limited in its input data by the simplicity of the objects morphology and specificity in systematization of the bacterial material (at first, of the recent one). So the morphological approach is an important but not enough criteria for diagnostic and systematization of the fossil objects. The products of bacterial vital activity expressed by lithology and geochemistry of rocks have got the special significance. The study of the carbon isotopes and structures of different biomarkers has the highest importance.
The main spheres of utility of results of the traditional paleontology and bacterial paleontology also differ strikingly. Firstly, the bacterial paleontology is very important for sedimentology, and consequently for study of the geneses of sedimentary mineral resources, including oil and gas. Also it has importance for paleogeography in the study of epicontinental basins. The highest significance of bacterial paleontological data is obvious for study of biosphere evolution, especially in the Precambrian and the Early Paleozoic. And finally, bacterial paleontology is one of the principle aspect for the astromaterials study.
For more precise understanding of what the bacterial paleontology is, we should face the term geomicrobiology [1, 4]. The main task for any geomicrobiologist is to study the interrelation between rocks and minerals from one side and bacteria (to be correct, microorganisms) from another side. Certainly, the main biological objects of such works are the recent organisms.
The geomicrobiological studies are extremely necessary not only for elucidation of the influence of recent microorganisms on geological environment, but also for actualistic interpretation of fossil objects. The geomicrobiological studies revealed that:
1) microorganisms actively affect on all minerals without exception forming biofilms on their surfaces, which in turn are the essential features of the weathering process in a broad sense;
2) microorganisms can accumulate various metals;
3) microorganisms assist the formation of a variety of minerals composing many sedimentary rocks (including dolomite, layer silicates, etc., and obviously calcium carbonate and phosphates).
Bacterial paleontology and geomicrobiology share some sphere of actions Actually, some geomicrobiological investigations are bacterial-paleontological ones and vice versa. These relationships are quite natural between closely related sciences.
The recent electron-microscopic studies of rocks varying both in chemical composition and age prove that fossilized microorganisms practically can be found in almost all sedimentary rock. The ancient phosphorites have become the classic object for such studies. It

Fig. 1 a. Fragments of
cyano-bacterial mat from Phosphorites (

Fig. 1 b. Cyanobacterial mat
and purpul bacteria in phosphorits (
is this study, that has revealed fantastic preservation of diverse cyanobacteria and other microorganisms [22, 45] (Fig. 1) (also see Zhegallo et al., this volume). Parallel investigations treated rocks aged from the Archean to Quaternary. The microorganisms of uncertain nature were found in the Archean [29, 38, 41, 51]. However, it has evoked some strongly negative valuations [6] repudiating some of the finds. But I assume, that the critics of the Shopffs work have done it in a not completely correct way.
The microorganisms from the Lower Proterozoic were found in the rocks of various types. The most interest finds were made in jaspilites (BIF) (Fig. 2a), shungites, cherty shales and, certainly, in stromatolites. The presence of cyanobacteria in all mentioned rocks could be considered as almost proven fact.
Going higher up the stratigraphical column, I should mention the finds of the fossil bacteria in the Vendian of the Russian Platform (Fig. 2b), in the Lower Cambrian of Australia (Fig. 2c), Middle and Late Cambrian of the Siberian Platform (Fig. 3) [20], Ordovician Dyctionema Shales of the NW Russian Platform, Domanik-like rock of Ural region, and thin clay layers in the Carboniferous of the Moscow Syneclise (for details see below).

Fig. 2 a. Coccoids bacteria in
BIF (

Fig. 2 b. Cyanobacteria mat in vendian of East European platform.

Fig. 2 c. Cyanobacteria in
Lower Cambrian (
Almost unexpected and unique results were obtained by the bacterial-paleontological study of graphite from the Botogol Deposit of East Sayan (South Siberia). Graphite of this deposit was considered to be magmatogenic for a long period, since the graphite bodies are hosted by nepheline-syenite. At first, the tubular structures similar in shape and size to cyanobacteria were found in the graphite [50]. But it could not convince the specialists on nepheline-syenites actually, the micron tubes have very simple morphology. The study of graphites had been continued [26] and in a short period of time numerous fungal-like remains and fragments of loricate ticks with perfectly preserved trachybotria were found (Fig. 4). The finds of ticks strongly supported the bacterial-paleontological results and closed the discussion on the graphite origin. It is obvious now, that graphites were formed in the result of conversion of highly carbonic sedimentary carbonate rocks.

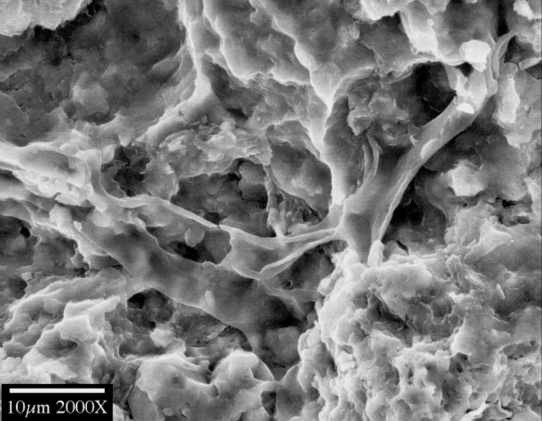
Fig. 3. Cyanobacterial mats with filaments and coccoids (Lower Cambrian, Synyaya formation, Siberian platform).
The deposits of sedimentary mineral resources give us another perfect example of the tremendous bacterial role in the geological history. Besides the above mentioned phosphorites,

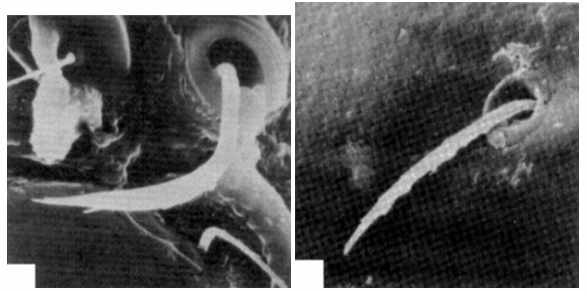
Fig. 4. a fungi-bacterial assosiation in graphites of Botogol deposites (Devonian, E. Sayan, Siberia); b-d fragments of oribatid mites with trichobotrias in fungi-bacterial assosiation (Devonian graphites, Siberia); e-f trichobotria of mites (Holocen, Siberia) (Krivolutsky et al., 1990).
jaspilites (BIF), graphites and rare metals ores, the following rocks are noteworthy: iron ores, manganese ores (including oceanic manganese concretions), bauxites, sulfides, gold, etc.
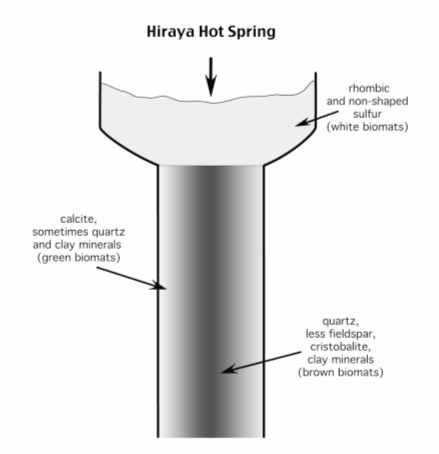
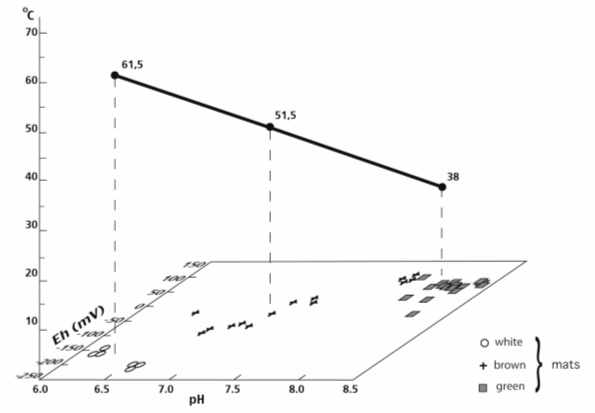
Fig. 5 a, b. Distribution of different types of mats and minerals in Hiraya Hot Spring, Japan (according of data by K. Tazaki et al., 1997).
As I have already noted mentioning Andrusovs lectures, the bacterial role in the accumulation of Fe, S, Si, P, Mn was undoubtful. Later we realized, that there are few elements in the Mendeleevs Periodical System (except for artificially derived and inert gases) which are not connected in some way with the bacterial activity. The most striking example of the latest studies is the Tomtor rare metal deposit (Siberian Platform) [48]. Here cyanobacteria act as a specific filter increasing the concentration of elements up to the tremendous values. For instance, the content of Nb is 12%!
It is clear today, that element-specific bacteria are not numerous, and their majority extracts a wide specter of elements from water. Even twenty years ago we knew somewhat twenty minerals that could originate with the microbial activity [19]. Recently the number of such minerals grew up significantly, and nowadays it is more than a hundred.
Quartz, cristobalite, pyrolusite, silicates (including layer ones), and feldspars are of special interest [4, 9, 21, 35].
Biomineralization process in modern hot springs and some terrestrial water basins convincingly demonstrates a rapid (minutes, hours) mineral formation with the participation of bacteria. Doubtless examples of authigenic bacterial mineralization were described by Tazaki et al. [35]. Quartz, crystobalite, barite, feldspar, sulfur, buserite, ferrohydrite, hematite, pyrite, jarosite, smectite, kaolinite, calcite and some other minerals were discovered in different environments of hot springs, lakes, mines, waterfalls. In contrast to previous fragmentary data on these minerals, the Japanese finds are very impressive. Therefore a title of their book is New world in the science of mineralization
Let us consider the example derived from the Hiraya hot spring. The spring is coptated. Three groups of minerals were formed in a box and a tray where three bacterial communities (white, brown and green mats) lived (Fig. 5a). Changes in temperature, pH and Eh values were followed (Fig. 5b). In the box with sausage-shaped bacteria (the white mat) located immediately at the hot spring, sulfur minerals of rhomboidal and uncertain form and fewer clay minerals (7 Ǻ) were formed. It has been long known that rhombic sulfur is generated with the participation of microorganisms.
Two different mats occurred in the chute. The brown mat of filamentous bacteria and bacilli was in the axial part, where quartz (3.3 Ǻ), feldspar (2.8 Ǻ), cristobalite (4.0 Ǻ), and clay minerals (7.0 Ǻ) were formed. The green mat of filamentous bacteria functioned at the peripheral parts, where calcite (3.0 Ǻ), quartz and clay minerals appeared.
Thus, in this case we can confidently state the bacterial participation in authigenic formation of the minerals listed; some values, e.g., those of pH, being registered.
It should be stressed that diffractograms of mineral structure most frequently show a distinct gallo reflecting the amorphous phase of these minerals. Tazaki and his colleagues unambiguously noted the amorphous phase of the minerals. This makes the successive stages of mineral formation more clear.
Among many results regularly published now [1, 4], those obtained by German researchers [14] are most impressive. The researchers revealed that forsterite was formed with the participation of microorganisms, such as cyanobacteria, actinomycetes, and partly lichens.
A very high (minutes, hours) rate of mineral formation was confirmed by numerous laboratory tests [10, 21]. This can explain the ideal preservation degree of fossil bacteria. In addition, the high rate of fossilization put forward a problem of dating the fossilization events, which could repeat over and over again in the geological history.
As it was already mentioned, the analyze of the roentgen-structural data of the mineral phase on bacteria or inside bacterial mats revealed the presence of amorphous mineral stage in most cases, except for carbonates. It is typical for minerals of quartz group, iron and manganese oxides, layer silicates, feldspars, etc. The amorphous stage is almost absent or unrecognizable by the roentgen-structural method in calcite and aragonite.
Probably, the preservation of fossilized bacteria is better when the amorphous stage is longer and more abundant. The worst preservation corresponds carbonates, since the cellular shells and membranes are destroyed by rapid crystallization almost without amorphous stage. Carbonate micrites are usually believed to be products of microbial activity.
In present time the most interesting examples of bacterial formation of minerals are obviously referred to those minerals that are widely known as composing of sedimentary rocks: clay minerals, feldspars, etc. Laboratory experiments and recent natural examples are already known [35]. However, the verification by data from the geological past is forthcoming.

Fig. 6 a. Coccoids bacteria in clay (Carboniferous, Moscow basin).

Fig. 6 b. Coccoids bacteria and biofilm in clay (Carboniferous, Moscow basin).
The first step to prove the idea on the authigenic formation of thin clay layers was the investigation by P.B. Kabanov, a specialist in the lithology and stratigraphy of Carboniferous deposits of the Moscow region. He studied using SEM the samples from thin layers from the Peski section (Fig. 6). The first illustration shows rounded bodied immersed within the unstructured matrix. It is noteworthy, that all bodied are 2-5 *m sized and have rounded shape. If it is terrigenous debris, it shapes would be obligatory angular under such sizes. So we have suppose, that it is coccoid bacteria.
The second case is more convincing. It was found out, that the rock is composed from spherical or granular bodies of 3-5 *m in size connected by thin threads. This situation is principally similar with the structure of the biofilms. I suppose, that further study of the clayish rocks will present us new evidences favoring the authigenic origin of most of them, especially thin layers with inconstant spreading. The main problem is how to recognize and calculate ratio between terrigenous (probably more abundant) and authigenic-bacterial compounds of the clay.
In connection with the studies of clays, the problems of high carbon rocks, including so called black shale, becomes very important. The above mentioned works on the Cambrian bituminous rocks of Siberia suggest that the earlier interpretation of such rocks as deep water formations is dubious.
The presence of benthic cyanobacterial remains in the Sinyaya Formation [3, 23] makes us to reinterpret the paleogeography of the Sinsk paleobasin. Instead of rather deep water basin opened towards the ocean, we can draw a shallow water partly closed sea with bottom anoxia (probably within the sediment) (Fig. 7). In addition to the biological evidences, the geological data also support this reconstruction. Firstly, the transition between transitional zone and eastern part of the basin is clearly traceable within the section of the middle reaches of Lena River. One has no chance to suggest 400 m depth for the basin without ignorance of the structural geology and stratigraphy. Secondly, the sections of the Kolyma Masiff (although fragmentary ones) correspond the transitional zone of the Siberian Platform in faunal and thickness characteristic.
All these facts make me to speak about the general features of the ancient epicontinental basin. At first, such type of the basin is absent in recent environment. These basins covered the significant part of modern continents, mostly shallow water, and probably with intensive hydrodynamics that formed numerous short-living and migrating islands. Owing to shallow
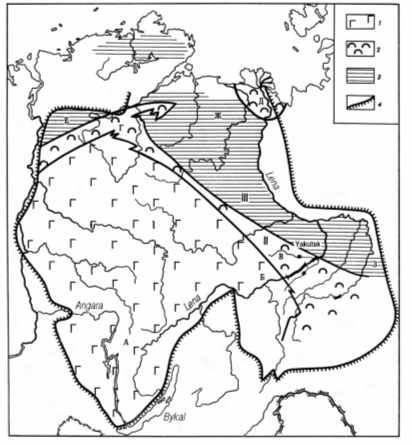
Fig. 7 a. Lower Cambrian paleogeography of the Siberian platform (1 evaporite basins, 2 biogerms, 3 deep water basins, 4 boundary of Siberian platform) (after V. Savitsky and V. Astashkin).
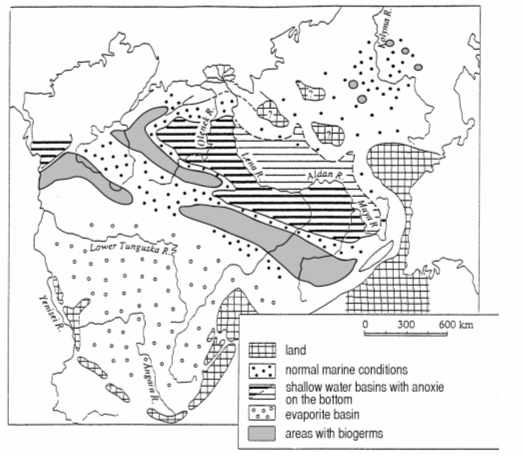
Fig. 7 b. Lower Cambrian (botomian) paleogeography of the Siberian platform (Rozanov, Zavarzin, 1997).
water conditions and fact, that a whole water column was placed within the photic zone, the water masses were rich in bacteria.
A study of paleogeographic reconstructions (Fig. 8) shows, that such gigantic pools were very typical for the Proterozoic and Paleozoic. They had no in common with the recent seas and oceans, so the character of sedimentation was different and to the significant extent was determined by the bacterial and especially by cyanobacterial activity.
All these data make a serious basis for study of the biomorphic structures in meteorites. It was the bacterial paleontology, that gave a possibility to reconsider already obtained and to get new data on carbon chondrites. A review of these data are present in several recent publications [11, 15, 21, 47]. It has especial importance, that all these works deal with objects of micrometric but non nannometric size. In addition, the problem of recognition of contamination commonly present in meteorites was also discussed.
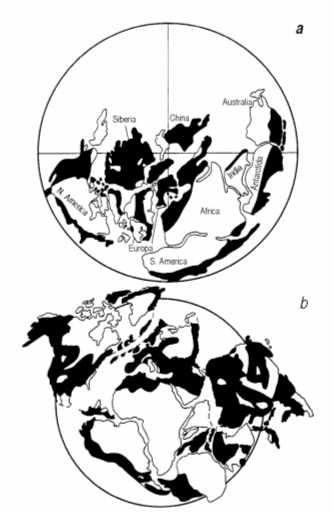
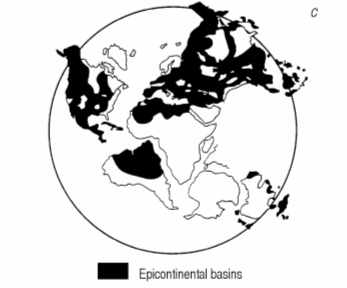
Fig. 8. Distribution of epicontinental basins (black) in Early Cambrian (a), Early Ordovician (b) and late Devonian (c).
Bacterial paleontology is rather young branch of paleontological studies, but the first steps have been made long ago. But during the last years the special bacterial-paleontological investigations made possible to formulate a series of statements which significantly changed our conceptions in different areas of knowledge.
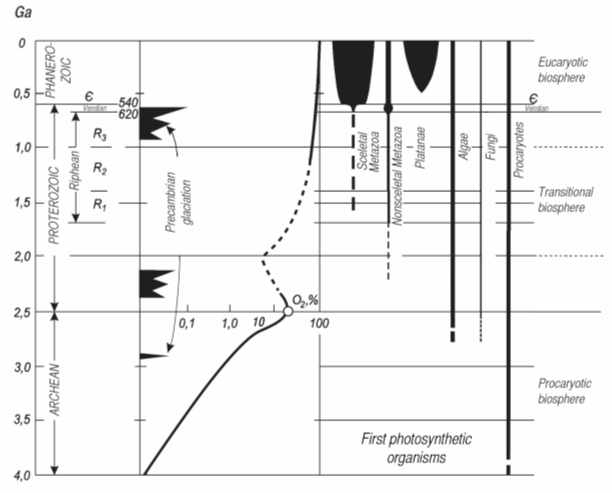
Fig. 9. Principal events in the evolution of the Preacmbrian biosphere (after Rozanov, 1992, supplemented).
At first, it has become clear that bacteria (and microbes in general) could be perfectly preserved as fossils, since the period of their fossilization is minutes our hours.
Secondly, we have realized, that the major part of the sedimentary rocks formed in the photic zone of epicontinental basins of the past originated under influence of microorganisms, in many cases with conclusive role of cyanobacterial communities.
Thirdly, corresponding to the previous statement, sedimentary mineral resources (ores and nonmetallics as well) usually originate with the significant assist of microorganisms.
At fourth, we have realized, that after special studies the number of minerals formed with the participation of microorganisms increase and already is above a hundred. Such minerals are the following: cristobalite, quartz, feldspar, layer silicates, oxides of Fe, Mn, etc.
At fifth, it is clear today, that the bacterial-paleontological data accompanied by the data on the first origin of eukariots, metazoa, etc. significantly enrich our knowledge on biosphere evolution (Fig. 9), and reveal the long period of transitional biosphere prior to the appearance of typical eukariotic biosphere of modern type.
At sixth, we understand, that the bacterial-paleontological data on carbon chondrites make foundation for the possibility of presence of extraterrestrial bacterial life.
References
[1] American Mineralogist, (1998), 83, 11-12, 2, 1387-1607.
[2] Amosov R.A., Vasin S.L., Kolesnikova E.B., (1995) The role of the biological factors in accumulation of gold in the sedimentss. Ecosystem restructures and the evolution of biosphere, Moscow, PIN RAN, 2, 147-153.
[3] Astafieva M.M., (in press) Biomorphic microstructures in the Sinsk formation (Lower Cambrian of the Siberian platform), Proc. SPIE.
[4] Banfield J.E, Nealson K.H., (eds), (1997) Geomicribiology: Interactions between Microbes and Minerals, Reviews in Mineralogy, 35, 448 p.
[5] Barghoorn T.S., Tyier S.A., (1965) Microorganisms from the Guntflint cherts. Science, 147, 3658, 563-577.
[6] Brasier M.D., Green O.K., Jephcoat A.P., Kleppe A.K., Van Kranendonk M.J., Lindsay J.E, Steele A., Grassineau N.V., (2002) Questioning the evidence for Earth's oldest fossils. Nature, 416, 76-81.
[7] Devouagard B., Posfai M., Xin Hua, Bazylinski D.A., Frankel R.B., Busek P.R., (1998) Magnetite from magnetotactic bacteria: Size distributions and twinning, American Mineralogist, 83, 1387-1398.
[8] Fortin D., Ferris F.G., Scott S.D., (1998) Formation of Fe-silicates and Fe-oxides on bacterial surfaces in samples collected near hydrothermal vents on the Southern Explorer Ridge in the northeast Pacific Ocean, American Mineralogist, 83, 11-12, 2, 1399-1408.
[9] Geptner A.R., Petrova V.V., Sokolova A.L., Gorkova N.V., (1997) Biochemical formation of layered silicates under hydrothermal changing of basalts, Iseland, Litologia i poleznye iskopaemye, 2, 249-259.
[10] Gerasimenko L.M., Goncharova I.V, Zhegallo E.A., Zavarzin G.A., Zajtseva L.V., Orleanskiy V.K., Rozanov A.Yu., Tichomirova N.S., Ushatinskaya G.T., (1996) The process of mineralization (phosphatization) of filamental cyanobacteria, Litologia i poleznye iskopaemye, 2, 208-214.
[11] Gerasimenko L.M., Hoover R.B., Rozanov A.Yu., Zhegallo E.A., Zhmur S.I., (1999) Bacterial Paleontology and Studies of Carbonaceous Chondrites, Paleontol. Journ., 33, 4, 439-459 (translated from Paleontol. Zhurnal, 4, 109-125).
[12] Gerasimenko L.M., Zavarzin G.A., (1993) The relict cyanobacterial communities in The problems of preanthropogenic evolution of biosphere, Moscow, Nauka, 222-254.
[13] Gerasimenko L.M., Goncharova I.V., Zavarzin G.A., Pochtareva I.V., Rozanov A.Yu., Ushatinskaya G.T., (1994) The Dinamic of release and absorbtion of phosphorus by cyanobacteria, in Ecosystem changing and evolution of biosphere, Moscow, Nedra, 1, 348-458.
[14] Gorbushina A., Boettcher M., Brumsack H.-J., Krumbein W.E., Vendrell-Saz M., (2001) Biogenic Forsterite and Opal as a Product of Biodeterioration and Lichen Stromatolite Formation in Table Mountain Systems (Tepuis) of Venezuela, Geomicrobiol. Journ., 18, 117-132.
[15] Hoover R.B., Rozanov A.Yu., (2002) Chemical Biomarkers and Microfossils in Carbonaceous Meteorites, Proc. SPIE, 4495, 1-18.
[16] Kholl A.H., (1985) Exeptional preservation of photosyntetic organisms in silicified carbonates and peats, Phil. Trans. R. Soc. London, Ser. B311, 11-22.
[17] Kirschvink J.L., Jones D.S., MacFadden B.J., (eds), (1985) Magnetite Biomineralization and Magnetoreception in Organisms, New York, London: Plenum, 2.
[18] Krivolutsky D.A., Druk, Eitminaviciute I.S., Laskova L.M., Karppinen E., (1990) Fossil oribatid mites, Mokslas Publishers Vilnius, 110 p.
[19] Lowenstam H.A., Weiner S., (1989) 0n biomineralization, Oxford Univ. Press, 324p.
[20] Rozanov A.Yu., (1999) Fossil bacteria and new view on the processes of sedimentation, Soros Educational Journ., 10, 63-68.
[21] Rozanov A.Yu. (ed), (2002) Bacterial Paleontology, Moscow, PIN RAN, 188 p.
[22] Rozanov A.Yu., Zhegallo E.A., (1989) 0n the problem of genesis of ancient phosphorites of Asia, Litologia i poleznye iskopaemye, 3, 67-82.
[23] Rozanov A.Yu., Zavarzin G.A., (1997) Bacterial Paleontology, Vestnic RAN, 67, 3, 241-245.
[24] Rozanov A.Yu., Barskov I.S., (1998) Diversity and Phylum Distribution of Biominerals, in Advanced Mineralogy (Marfunin A.S., ed), v.3. Mineral Matter in Space, Mantle, Ocean Floor, Biosphere, Environmental Management and Jewelry, Printed in Germany, 247-255.
[25] Rozanov A.Yu., Barskov I.S., (1998) Magnetite Biomineralization, Magnetofossils, and Magnetoreception in Organisms, in Advanced Mineralogy (Marfunin A.S., ed.), v.3. Mineral Matter in Space, Mantle, Ocean Floor, Biosphere, Environmental Management, and Jewelry, Printed in Germany, 255-257.
[26] Rozanov A.Yu., Zhegallo E.A., Hoover R., (1999) Microbiota of the Botogol graphites, Proc. SPIE, 3755, 38-46.
[27] Schopf J.W., (ed), (1983) Earth's earliest biosphere: its origin and evolution, Princeton Univ. Press, 544 p.
[28] Schopf J.W., (1993) Microfossils of the Early Archean Apex Chert: new evidence of the antiquity oflife, Science, 260, 640-646.
[29] Schopf J.W., Kudryavtsev A.B., Agresti D.G., Wdowlak Th.J., Czaja A.D., (2002) Laser-Raman imagery of Earth's earliest fossils, Nature, 416, 73-76.
[30] Schopf J.W., Klein C. (eds.), (1992) The Proterozoic biosphaere: a multydisciplinary study, Cambridge: Cambridge Univ. Press, 1348 p.
[31] Sergeev V.N., Knoll A.H., Grotzinger J.R., (1995) Paleobiology of the Mezoproterozoic Billiakh Group, Anabar Uplift, Northeastern Siberia, Journal of Paleontology, 69, Memoir 39, 37 p.
[32] Sergeev V.N., (1992) Silicificated microfossils of Precambrian and Cambrian of the Urals and Middle Asia, Moscow, Nauka, 134 p.
[33] Sergeev V.N., Kholl A.H., Zavarzin G.A., (1996) The first three billion years of life: from procariots to eucariots, Priroda, 6, 54-67.
[34] Skinner H.C., Fitzpatrick R.W., (eds), Biomineralization Processes, Iron, Manganese, Cremlingen, Germany: Catena Verlag, Catena Suppl., 21.
[35] Tazaki K., Aoki A., Asada R., etc. (1997) A new world in the science ofbiomineralization - Enviromental Biomineralization in Microbial Mats in Japan, Science Reports of Kanazawa Univ., 42, 1-2, 64 p.
[36] Visscher P.T., Reid R.P, Bebout B.M., Hoeft S.E., Macintyre I.G., Thompson Jr. J.A., (1998) Formation of lithified micritic laminae in modern marine stromatolites (Bahamas): The role of sulfur cycling, American Mineralogist, 83, 11-12, 2, American Mineralogist, 83, 11-12, 2, 1482-1493.
[37] Vologdin A.G., (1947) Geological activity of microorganisms, Izvestiya AN USSR, Ser. Geol., 3, 19-36.
[38] Walsh M.M., (1992) Microfossils and possible microfossils from the Early Archean Onver-wacht Group, Barbeton Mountain Land, South Africa, Precambrian Res., 54, 271-293.
[39] Weslall F., Rince Y., (1994) Biofilms, microbial mats and microbe-particle interactions: electron microscope observations from diatomaceous sediments, Sedimentology, 41, 147-162.
[40] Westall F, Boni L., Guerzoni M.E., (1995) The experimental silicification of microorganisms, Paleontol., 38, 495-528.
[41] Westall F, Walsh M.M., (2002), Early Archean fossil bacteria, in Bacterial Paleontology, Moscow, PIN RAN, 84-90.
[42] Yates K.K., Robbins L.L., (1998) Production of carbonate sediments by a unicellular green algae, American Mineralogist, 83, 11-12, 2, 1503-1509.
[43] Zavarzin G.A., (1984) Bacteria and the composition of biospere, Moscow, Nauka, 199 p.
[44] Zhang Chuanlun, Vali H., Romanek Ch.H., Phelps T.J., Liu Sh.V, (1998) Formation of single-domain magnetite by a thermophilic bacterium, American Mineralogist, 83, 1409-1418.
[45] Zhegallo E.A. et al., (2000) Atlas of Microorganisms from ancient phosphorites of Khubsugul (Mongolia), Huntsville, Alabama, USA, NASA, 167 p.
[46] Zhmur S.I., Rozanov A.Yu., Zhegallo E.A., Lobzova R.V, (1993) Cyanobacterial benthic system - the producent of carbonaceous material of shungites of the Lower Protherozoic of Karelia, Litologia i poleznye iskopaemye, 2, 122-124.
[47] Zhmur S.I., Rozanov A.Yu., Gorlenko V.M., (1993) Lithified remaines of microorganisms in carbonaceous chondrites, Geochimia, 1, 66-68.
[48] Zhmur S.I., Rozanov A.Yu., Kravchenko S.M., Zhegallo E.A., (1994) About genesis of rich rare-earth ores of Tomtor (the north of the Siberian platform). Reports RAN, Ser. geol., 336, 3, 372-375.
[49] Zhmur S.I., Rozanov A.Yu., Sokolov B.S., Bazhenova O.K., Gorlenko V.M., (1994) Bacterial mats as a resource of the maternal mater of oil, Moscow, DAN RAN, 6, 742-744.
[50] Zhmur S.I., Rozanov A.Yu., Lobzova R.V., Zhegallo E.A., (1996) About resource of carbon of graphite ores of Botogol sienite massif (East Sayan), Moscow, DAN RAN, Ser. Geol., 348, 3, 360-362.
[51] Zhmur S.I., Duda V.I., Skryabin G.K., Roizeman F.M., (2002) Microfossils in Early Archean graphites of Aldan Shield and some aspects of panspermia [4495-04], Proc. SPIE, v. 4495, 19-26.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 4260
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved