| CATEGORII DOCUMENTE |
| Alimentatie nutritie | Asistenta sociala | Cosmetica frumusete | Logopedie | Retete culinare | Sport |
Tesutul muscular
Fibra musculara este o celula cu functie contractila care are capacitatea de a dezvolta tensiune si capacitatea de a se scurta.
Muschiul este o structura inalt organizata si specializata, contractia fiind o functie complexa sub controlul motoneuronilor sistemului nervos central voluntar care trebuie sa asigure un lucru mecanic intens cu un cost energetic redus in conditii de rapiditate, repetitivitate, precizie si rezistenta in timp.
Caracterul
striat, tarcat al fibrelor musculare de la nivelul muschiului scheletic si a
muschiului cardiac datorat particularitatilor lor de structura (ce la
microscopul optic apar ca o succesiune regulata de benzi de culoare deschisa si
de culoare inchisa) le-a dat denumirea generica de muschi striat.
Fibrele musculare
scheletice sunt celule mari, cu mai multi nuclei - uneori extrem de multi -
cu un diametru intre 10-100 μm (cele mai multe intre 20-80 μm) si o
lungime foarte mare, de peste 4 cm. In tesutul embrionar nucleii sunt
localizati in centrul fibrei musculare. Ulterior multimea acestor nuclei fuge
spre periferie. Citoplasma este ocupata de catre miofibrile, de catre granule
de glicogen si de catre mitocondrii, extrem de abundente mai ales in tipul
oxidativ de fibre musculare, asa zisele fibre de tip I.
Elementele
contractile cuprinse in sarcoplasma unei fibre musculare sunt fibrilele.
Sarcoplasma unei singure fibre musculare poate contine pana la 10.000 de
microfibrile.
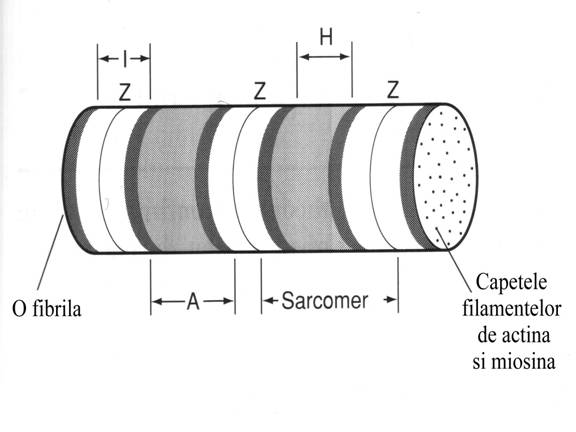
Miofibrilele
reprezinta structuri fibroase subcelulare cu diametrul de 1 μm.
Unitatea
contractila fundamentala a muschiului este sarcomerul.Lungimea
sarcomerului in repaus este de 2,5 μm. Fiecare sarcomer poseda doua
tipuri de filamente (subtiri si groase) de natura proteica, filamente aranjate
in paralel cu axul fibrei si partial suprapuse.
Filamentele
subtiri au o largime de 7 nm si o lungime de 1 μm iar filamentele groase o
largime de 10-14 nm si o lungime de 1,6 μm.
Aranjamentul
filamentelor subtiri si groase produce aspectul striat prin succesiunea
regulata de benzi dense (cu o lungime de 1,6 μm) si benzi de aspect mai
clar la microscop.
Benzile dense contin
filamente groase si sunt denumite benzi A fiind anizotropice la microscopie
(indici de refractie diferiti pentru diferite planuri de polarizare ale luminii
incidente).
Benzile de culoare deschisa,
denumite I (sunt izotrope, au un singur indice de refractie), sunt
alcatuite din filamente subtiri, extinse simetric in raport cu o linie mai
densa ce le centreaza, asa numita linie Z. Filamentele subtiri se extind de o
parte si de alta a liniei Z pe cate 1 μm.
Linia Z este alcatuita de o proteina in retea ce apare in
dezvoltarea embrionara inaintea filamentelor subtiri si impletindu-se cu
acestea, le ancoreaza. Astfel este mentinuta dispunerea regulata a complexului
filamentelor subtiri.
Sarcomerul este alcatuit din structurile cuprinse intre doua
linii Z consecutive. Deci un sarcomer este alcatuit dintr-o banda A avand la
fiecare din capete cate o jumatate de banda I.
In
fibra musculara exista trei tipuri de retele filamentoase. Pe langa
reteaua filamentelor groase (miozina) si reteaua filamentelor subtiri (actina
si alte proteine) exista si o retea filamentoasa ce asigura elasticitatea
musculara.
Elasticitatea
musculara intrinseca este asigurata de catre doua proteine gigante titina si nebulina. Titina sau
conectina este o proteina giganta (3000 KDa) si extrem de abundenta (cantitativ
a treia proteina a muschiului dupa actina si miozina). O portiune a moleculei
bogata in resturi de prolina, acid glutamic, valina si lizina asigura
elsaticitatea titinei si impune o tensiune pasiva sarcomerelor.
Nebulina (600-900 KDa)
reprezinta 3% din proteinele musculare si ancoreaza actina in discurile Z.
Prezenta pe toata lungimea filamentului fin ea regleaza numarul de monomeri de
actina ce constituie acest filament.

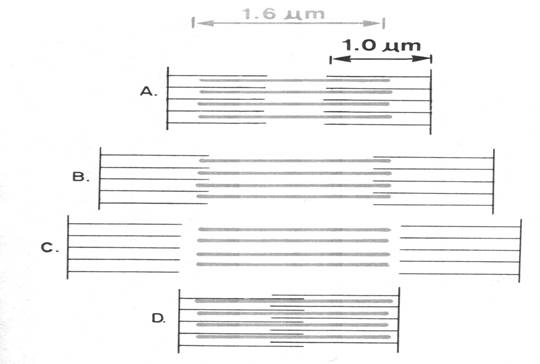
In
conditii de repaus in majoritatea muschilor cele doua tipuri de filamente se
interpatrund pe o lungime destul de mare la fiecare capat a benzii A.
Daca muschiul
este intins, supus tractiunii fibrilele groase si fibrilele scurte se separa,
nu se mai intrepatrund.
La o lungime a
sarcomerului de 3,6 μm miofibrilele subtiri sunt trase afara din banda A
si cele doua tipuri de fibrile scurte dispuse cap la cap.
Daca contractia este extrem de puternica nu
numai ca fibrilele subtiri si groase se suprapun ci fibrilele subtiri dintr-o
parte si alta aluneca unele catre celelalte si se suprapun intre ele.


Membrana fibrei
musculare - sarcolema
Membrana
ei bazala alcatuita din mucopolizaharide si fibre de colagen se contopeste la
extremitatile celulei cu tendoanele.
Membrana confera rezistenta la intinderea
muschiului si impreuna cu structurile tesutului conjunctiv ce inconjoara
fibrele musculare realizeaza aparatul "elementelor elastice paralele" cu rol
mecanic.
Membrana izoland fiecare fibra musculara una
de cealalta permite functionarea muschiului pe baza unitatilor motorii: un
motoneuron inerveaza un numar mai mic sau mai mare de fibre musculare ce se pot
afla la distanta una de alta si care actioneaza ca un ansamblu coerent si
izolat de restul muschiului.
Sarcolema se
invagineaza la nivelul fiecarui sarcomer pentru a forma o retea membranara de
tubusoare transverse deservind fiecare miofibrila.
Sarcotubulele se
invagineaza la nivelul fiecarei linii Z sau la jonctiunea benzilor A-I. Ei au
un diametru de 0,03 μm si permit undei de excitatie sa patrunda rapid in
fibra si sa excite practic simultan miofibrilele.
Reticulul sarcoplasmatic
este un sistem tubular complex ce inconjoara miofibrilele. Diametrul
tubusoarelor este de 0,04 μm, ele mergand paralel cu miofilamentele.
Reticulul sarcoplasmic fixeaza calciu printr-o ATP-aza calciu si magneziu
dependenta. Tubusoarele se pot extinde pe toata lungimea sacromerului si se
termina in structuri denumite cisterne terminale. Functiile
reticulului sarcoplasmic sunt eliberarea de calciu in timpul contractiei
musculare si sechestrarea si depozitarea calciului in timpul relaxarii
musculare.
De o parte si de alta a acestui spatiu
jonctional membranele sunt reunite de niste picioruse, structuri repetitive
denumite "spanning proteins".
Un grup
de sarcotubule si doua cisterne terminale (in total trei elemente) este denumit
triada.
Triadele
sunt regiuni unde sistemul tubular este in stransa contiguitate cu partile
umflate ale reticulului sarcoplasmatic (asa zisele cisterne).
Triadele sunt
elemente cheie ale cuplajului excitatie-contractie. In timpul excitatiei
musculare calciul stocat in cisternele terminale este eliberat pentru a se
putea fixa pe troponina si sa permita interactiunea filamentelor groase cu
filamentele subtiri.
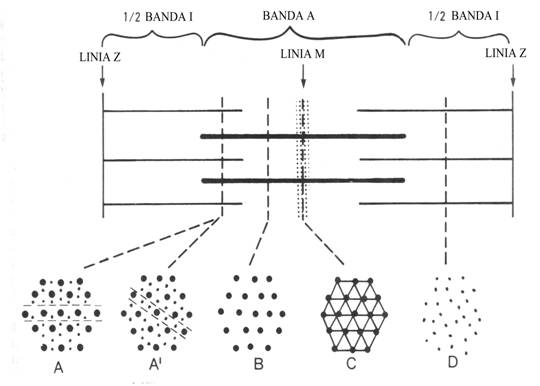
Filamentele subtiri sunt compuse
din trei tipuri de proteina-actina, tropomiozina si troponina aflate in
proportie de 7/1/1.
Unitatea
functionala necesara pentru relaxare cuprinde 7 monomeri de actina, 1 complex
de tropomiozina si 1 complex de troponina.
Filamentele
groase sunt alcatuite din miozina si in mult mai mica masura
din alte proteine.
Numai actina si
miozina sunt direct implicate in generarea de tensiune, complexul tropomiozinic
si troponinic sunt proteine de reglare intrucat regleaza interactiunea
actina-miozina.
Filamentele
subtiri sunt formate din doua lanturi de actina F formate din polimerizarea actinei G (globulare).
Actina
G este o proteina
monomerica globulara cu o greutate de 42.000 daltoni. Un glob de actina G are
un diametru de 4-5 nm.
Actina
F este un polimer alcatuit din peste 300 monomeri de actina G.
Tropomiozina previne interactiunea miozinei cu actina,
efect modulat de troponina.
Tropomiozina este o
proteina alungita, alcatuita din doua lanturi in elice. Are o greutate de
66.000 daltoni si este dispusa in spatiul dintre doi polimeri de actina.
Fiecare molecula de tropomiozina se extinde peste sapte monomeri de actina.
Troponina este o
proteina globulara, de fapt un complex alcatuit din trei proteine. Are o
greutate de 70.000 daltoni.
Din cele 3 proteine ce
alcatuiesc troponina, troponina T se leaga de tropomiozina, troponina-I inhiba
ATP-aza actomiozinei iar troponina-C leaga calciul.
In prezenta calciului
configuratia complexului troponina-tropomiozina se modifica si miozina poate interactiona
cu actina.
Biochimic
actina si miozina puse impreuna in conditiile disocierii ATP se produce
supraprecipitare cu formare de "dopuri" si rezulta contractie musculara.
Actina si miozina puse impreuna
in conditiile in care nu se mai produce ruptura moleculei ATP (bunaoara absenta
Ca++) se produce disocierea actomiozinei, scaderea vascozitatii si relaxare
musculara.
ATP este
singura sursa de energie pe care o pot
folosi proteinele contractile dar fibra musculara are rezerve numai pentru opt
contractii. In timpul contractiei musculare ATP este regenerat din ADP folosind
energia din depozitele mai generoase de creatinfosfat.
Contractia musculara implica ca filamentele groase
si filamentele subtiri sa alunece unele de-a lungul celorlalte. Aceasta alunecare
este rezultatul fortei ce o exercita puntile create la nivelul capetelor de
miozina ce trag de fibrele de actina spre centrul sarcomerului.
Care este procesul fizic intim legat de crearea
acestor punti nu este inca bine cunoscut. Fiecare punte se fixeaza, trage si se
detaseaza de actina. Energia este furnizata de catre ATP. Procesul este reglat
de catre calciu, troponina si tropomiozina.
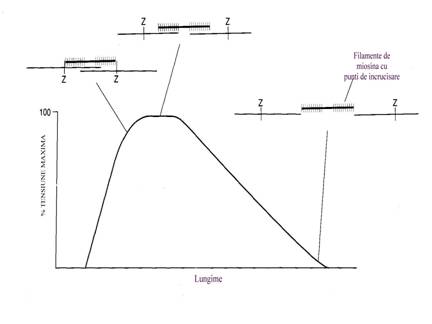

Forta de alunecare produsa depinde de cantitatea de calciu eliberata de catre reticulul sarcoplasmic. Hidroliza ATP realizata de ATP-aza miozinei permite rotatia capetelor de miozina in jurul situsului de legare a actinei. Forta de alunecare depinde de numarul de punti active si de viteza de fixare si detasare a puntilor. Exista si o diferenta intre fortele sarcomerice individuale.
Detasarea
puntilor este de asemeni un proces activ, consumator de ATP. Sarcomerele
actioneaza asemeni unor cremaliere minuscule (Lassagne F.).
Reticulul sarcoplasmic este
actorul privilegiat al acestei piese. El are rol esential atat in contractie
(eliberarea calciului) cat si in relaxarea musculara (captarea calciului
intracitoplasmatic). Contractia depinde de activarea canalului calcic cuplat cu
receptorul sensibil la rianodina
Relaxarea depinde de capacitatea de stocare a calciului
prin actiunea unei ATPaze calciu-magneziu dependente ce reconstituie depozitele
calcice reticulare impotriva gradientului de concentratie.
Transportul a
doi ioni de calciu in interiorul reticulului sarcoplasmic "costa" o molecula de
ATP ce urmeaza a fi hidrolizata.
Calciu este
pompat in timpul relaxarii musculare de catre reticulul longitudinal (membrana
cu foarte multe pompe) iar stocarea si eliberarea se face de catre reticulul
jonctional care are putine pompe calcice dar multe canale calcice si care
contine proteina ce leaga calciul, calsequestrina.
Cand potentialul de actiune prin sistemul de tuburi T
depolarizeaza membrana se induce eliberarea calciului din reticulul
sarcoplasmic in citoplasma.
Concentratia
calciului liber in citoplasma creste de o suta de ori (de la 0,1 μM la 10
μM).
Calciul interactioneaza cu troponina C,
tropomiosina trage de actina expunand-o miozinei, se formeaza punti, este
suprimata inhibarea ATP-azei actomiozinice de catre troponina-I. Rezulta contractia
musculara.
Cand reticulul sarcoplasmatic
sechestreaza calciu si se restabileste inhibitia prin troponina - tropomiozina
se produce relaxare musculara.
Cum se
transmite impulsul de la nerv la muschiul striat?
Cea mai mica
parte dintr-un muschi ce se poate contracta independent se numeste unitate
motorie.
Unitatea
motorie este alcatuita dintr-un singur neuron motor si de grupul
de fibre musculare pe care acesta le inerveaza.
Frecventa
impulsurilor ce vin prin neuronul motor variaza mult, limita inferioara a
impulsurilor fiind de 16/secunda iar limita maxima ce duce la o contractie
rapida, brutala a fibrei este de 50/secunda (maximum cat poate propaga un axon
de mamifer).
Numarul
fibrelor musculare dintr-o unitate motorie a unui muschi variaza in functie de
precizia cu care tensiunea dezvoltata de catre respectivul muschi este
controlata.
Cel mai mic
numar de fibre musculare per unitate motorie au muschii ce dezvolta miscari de
mare finete: muschii striati oculomotori au mai putin de 10 fibre musculare per
unitate motorie, muschii ce misca osisoarele urechii mijlocii au 10-125 fibre,
muschii faringieni au 2-6 fibre musculare iar muschii laringieni au 2-3 fibre
musculare per unitate motorie. Muschii ce executa miscari de mica precizie, cum
ar fi muschiul gastrocnemian au un raport mare de 1/2000.
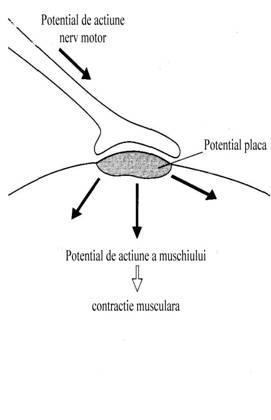
Contactul dintre axonul neuronului motor si fibra
musculara se face la nivelul unei formatiuni specializate denumite placa
neuro-motorie.
-De altfel,
intre cele doua nu exista un contact direct. La nivelul placii motorii membrana
modificata a fibrei musculare formeaza o serie de pliuri. -Terminatiile demielinizate ale
neuronului motor se afla in santurile menajate de aceste pliuri intre cele doua
structuri fiind o fanta sinaptica foarte ingusta de 500 A. -Terminatiile
nervoase poseda multiple vezicule sinaptice ce contin acetilcolina.
-Pe partea
opusa, in crestele placii motorii se afla receptori nicotinici.
-In fanta
jonctionala se afla enzima acetilcolinesteraza. - Acetilcolina
are un rol central in procesul de transmisie neuromusculara.
-Transmisia neuromusculara este un proces de amplificare intrucat potentialul de actiune a nervului produce un potential de actiune a muschiului cu mult mai mare.
Acetilcolina este sintetizata in axoplasma neuronului din
colina si acetilcoenzima A.
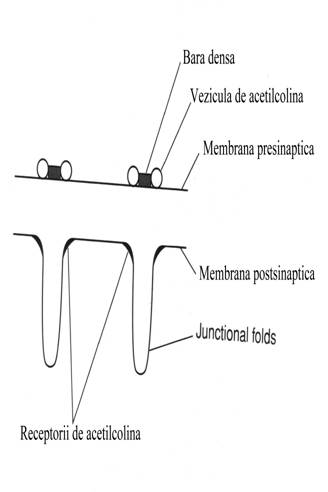
In fata crestelor de pe
faldurile postsinaptice de pe membrana muschiului sunt dispuse pe capatul
nervos presinaptic asa numitele bare dense avand la capat vezicule de
acetilcolina imediat mobilizabile. Stimularea
nervului creste sinteza de acetilcolina prin cresterea concentratiei
intracelulare de sodiu
Acetilcolina
este depozitata in nervul terminal in vezicule. O cantitate de 80% din
acetilcolina depozitata in nerv poate fi eliberata de catre potentialul de
actiune.

Ce anume varsa
o vezicula in fanta sinaptica? Pe langa
acetilcolina se elimina ATP, veziculina, colesterol, fosfolipide si calciu.
Toti componentii sunt reciclati si reintrodusi in circuit.
Un potential de
actiune al nervului implica exocitoza a 200 de vezicule ce contin 200x1500
300.000 de molecule de acetilcolina ce declanseaza un potential la nivelul
placii muschiului.
Este un
mecanism cu o marja de securitate de 500%.
Ca impulsul sa
se propage prin nerv sarind peste nodurile lui Ranvier este necesara prezenta
calciului extracelular. Exocitoza veziculelor este de asemenea conditionata de
catre calciu.
Calciul se leaga de proteina
numita calmodulina, complexul calciu-calmodulina activeaza diverse enzime si
afecteaza proteinele structurale ale veziculelor inclusiv sinapsina.
Vezicula fuzioneaza cu membrana
prejonctionala a nervului si isi varsa continutul in fanta.
Pentru fiecare molecula de
acetilcolina eliberata exista 10 receptori ce asteapta de cealalta parte a
fantei. Dupa excitatia receptorului porul nicotinic a acestuia se deschide,
membrana devenind permeabila pentru sodiu, potasiu, calciu, magneziu si amoniu.

Fiecare molecula de acetilcolina poate stimula un singur receptor de nicotina si sa deschida canalul ionic doar pentru o milisecunda.Dupa acest interval acetilcolina este hidrolizata de catre acetilcolinesteraza.
Acetilcolinesteraza
este localizata in pliurile jonctionale si in lamina bazala a structurilor
fantei sinaptice. Fiecare molecula de acetilcolinesteraza are sase situsuri
enzimatice ce atrage si hidrolizeaza acetilcolina.
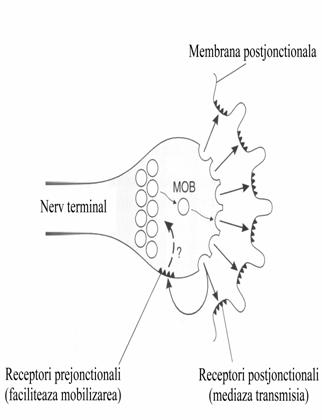
Receptori nicotinici se afla si
pe terminatiile nervului ei avand rolul sa creasca mobilizarea acetilcolinei si
eliberarea acesteia din vezicule printr-un feed-back pozitiv.
Printr-un mecanism inca incomplet inteles receptorii de
acetilcolina dispusi prejonctional mobilizeaza veziculele de acetilcolina sa se
deplaseze in zona activa gata de eliberare in cazul in care soseste un alt
impuls nervos. MOB=mobilizare.
Potentialul
transmembranar la nivelul placii este de - 90mV. Interiorul fibrei musculare
este electronegativ. In conditii normale de functionare se produce o
depolarizare de 40 mV, potentialul transmembranar scazand la -50 mV.
Cand
potentialul placii neuromotorii atinge acest nivel critic se propaga un
potential de actiune ce se supune legii tot sau nimic si care trece via
sarcolema sa elibereze Ca++ din reticulul sarcoplasmatic si sa
activeze contractia fibrei musculare.

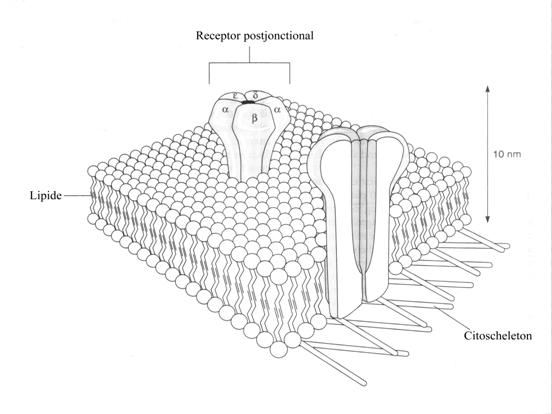
Receptorii nicotinici
Receptorii nicotinici pentru
acetilcolina a membranei postsinaptice sunt dispusi in ciorchini distincti pe
"umerii" faldurilor membranei jonctionale. Fiecare ciorchine are un diametru de
0,1M si contine cateva sute de receptori. Fiecare receptor are cinci
subunitati din care doua, asa zisele alfa (cu o greutate moleculara de 40.000
daltoni) sunt identice.
Receptorii
alcatuiesc structuri cilindrice ce traverseaza membrana celulara si care
alcatuiesc un canal central, asa numitul ionofor, care in mod obisnuit este
inchis.
Fiecare unitate alfa are pe suprafata sa
extracelulara o unica zona de legare pentru acetilcolina, zona in care se pot
fixa si pahicurarele.
Pentru ca un
receptor sa fie activat trebuie ca ambele situsuri alfa sa fie ocupate.
Consecutiv legarii celor doua situsuri din subunitatile alfa se produce o
modificare structurala a complexului receptor care deschide pentru o clipa
canalul ionofor(aici clipa este egala cu o milisecunda) .

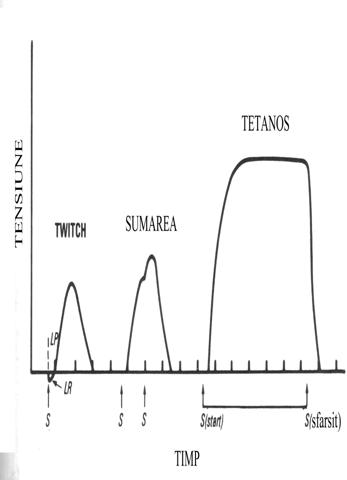
O
stimulare unica produce un ciclu contractie-relaxare a muschiului numit secusa.
Secusa este precedata de o
perioada de latenta de 2-4 msecunde iar uneori in aceasta perioada muschiul ce
trebuie sa execute contractia se lungeste. Este asa numita relaxare de
latenta a carei substrat subcelular nu se cunoaste dar care creeaza
conditii optime pentru dinamica puntilor dintre fibrele contractile ale
muschiului.
O stimulare
repetata, efectuata inaintea relaxarii complete a muschiului, duce la o
contractie care este sumata cu prima. Este asa numitul fenomen de sumare.
Efectul de
sumare este rezultatul aditionarii calciului liber din
sarcoplasma, eliberat din reticul de cel doua stimulari.
Repetarea la
rata mare a stimulului va face ca muschiul sa ramana constant contractat. Este
asa numitul tetanus (in greaca veche tetanos=rigid).
Platoul contractiei tetanice este la un nivel mai ridicat
de tensiune decat varful secusei.
Tetanosul nu
poate fi mentinut foarte mult timp din cauza oboselii a carei mecanism intim
ramane necunoscut
O diviziune de timp este de 5 ms.
S=stimulus, LP=perioada de latenta, LR=relaxare de latenta

Muschiul scheletic este un ansamblu de fibre cu caracter divers, avand proprietati metabolice si contractile proprii. -muschii posturali ce actionand antigravitational mentin postura. Sunt alcatuiti prioritar din fibre de tip I, foarte rezistente la oboseala, fibre "rosii" cu contractie lenta si metabolism oxidativ; -muschii fazici (de actiune), cei ce permit diverse miscari. Contin fibre cu contractie rapida, IIa de tip metabolic oxidativ si glicolitic foarte rezistent la oboseala si fibre de tip IIb, de tip metabolic glicolitic, foarte sensibile la oboseala.
Fibrele
musculare rosii
Fibrele ce isi
asigura ATP prin fosforilare oxidativa au un echipament pe masura. Ele au un
important echipament mitocondrial si sunt inconjurate de numeroase vase
sanghine. Au de asemeni un fixator proteic reversibil de oxigen, mioglobina.
Mioglobina
creste rata de difuziune a oxigenului in fibra musculara si asigura un mic
depozit de oxigen. Mioglobina da si culoarea caracteristica a fibrei oxidative,
o culoare rosie inchisa. Din aceasta
cauza fibrele oxidative sunt deseori denumite si fibre musculare rosii.
Fibrele musculare albe
Fibrele ce asi asigura
ATP-ul prin glicoliza sunt denumite si fibre albe. Ele au putine
mitocondrii,dar multe enzime glicolitice.
Cele doua
caracteristici generale amintite (capacitatea ATP-azica a miozinei si tipul
metabolic de generare a ATP-ului) determina clasificarea fibrelor musculare
striate scheletice in trei tipuri:
-Fibrele de tip I
(lent-oxidative). Sunt fibre cu activitate ATP-azica a miozinei scazuta si cu
mare capacitate oxidativa.
-Fibrele de tip II a (rapid-oxidative).
Sunt fibre cu activitate ATP-azica a miozinei ridicata si cu mare capacitate
oxidativa.
-Fibrele
de tip II b (rapid-glicolitice). Sunt fibre cu activitate ATP-azica
a miozinei ridicata si cu capacitate glicolitica mare.
MUSCHII NETEZI
Muschii netezi, involuntari sunt
dispusi in peretele organelor cu lumen (tub digestiv, cai urinare, artere) si
in jurul retinei regland accesul luminii la aceasta.
Fibra musculara neteda are
diametrul de 20 ori mai redus ca fibra striata si o lungime de mii de ori mai
redusa. Au o mare elasticitate si rezistenta la efort. Aceste fibre se pot
intinde la dublul lungimii din starea de repaus si suporta timpi de contractie
de 30 ori mai lungi ca muschii voluntari.
Pe plan metabolic au o deosebita eficacitate
energetica
Oboseala musculara este scaderea
tensiunii pe care o dezvolta muschiul ca efect al activitatii contractile
prealabile. Se produc de asemeni scaderea vitezei de scurtare si scaderea
vitezei de relaxare.
Este legata oboseala de scaderea cantitatii de ATP
din muschi? S-a
constatat ca in muschiul obosit cantitatea de ATP este doar usor mai redusa
decat in muschiul aflat in stare de repaus si nu suficient de scazuta pentru a
afecta ciclurile de cuplare a puntilor dintre proteinele contractile.
Daca nu ar
aparea oboseala, ATP-ul ar scadea pana cand puntile dintre proteinele
contractile ar fi blocate intr-o maniera de tip rigor. Este deci posibil ca
oboseala a aparut ca un mecanism biologic ce evita fenomenele de rigor.
In timpul
eforturilor scurte de mare intensitate se produce o insuficienta a cuplarii intre
excitatie si contractie.Recuperarea dupa eforturi rapide de scurta durata este
rapida (de ordinul minutelor) pe cand recuperarea dupa eforturi de lunga durata dureaza ore.
FRISONUL
Muschiul
intervine in termoreglare asigurand termogeneza prin cresterea tonusului
muscular si prin frison.
Frisonul
termic este o succesiune de secuse clonice ale musculaturii
striate. Frecventa contractiilor este de 5-10 pe secunda. Comanda motorie este
extrapiramidala.
Productia de
caldura in timpul frisonului generalizat este de cinci ori cat cea din repaus.
Contractia izometrica maximala produce 1 C pe minut (50
w per kg de muschi).
Termogeneza
prin frison nu poate fi mentinuta mai mult de 3-5 ore.
Cum este
alcatuita masina chimica ce furnizeaza energie muschiului? Sursa de
energie a contractiei musculare este ruptura enzimatica a unei molecule
macroergice si anume molecula de ATP. Legaturile chimice intre cele trei
resturi de fosfat sunt macroergice, fiecare dintre ele stocand 7300 calorii/mol
de ATP. Aceasta cantitate este suficienta pentru cel mult 3 secunde de
activitate musculara maxima.
Din aceasta cauza exista mecanisme, multiple,
ce refac stocul de ATP. Exista in celula musculara un depozit a unei alte
substante ce contine o legatura macroergica si anume creatinfosfatul.
Legatura macroergica contine o cantitate mult
mai mare de energie decat cea a ATP-ului (10300 calorii/mol in raport cu 7300)
si in multi muschi cantitatea de creatinfosfat este de 2-4 ori mai mare decat
cea a ATP.
Putem vorbi de
un sistem unic ingloband ATP si creatinfosfatul
O alta sursa de refacere al ATP-ului muscular este sistemul
glicogen-acid lactic.
Adrenalina,
hormonul stresului, activeaza glicogenoliza atat la nivel hepatic cat si la
nivel muscular, glucagonul activeaza numai glicogenoliza hepatica.
Degradarea
glicogenului si a glucozei se produce anaerob si rapid, producerea energiei
este de 2,5 ori mai rapida decat pe calea oxidativa. Deci exista un
sistem autonom, independent de oxigen, si rapid. Este un mecanism de "a doua
linie" cu o viteza pe jumatate cat a sistemului fosfagen dar asigura la o
performanta scazuta in raport cu fosfagenul o durata mai mare de actiune de 10
ori mai lunga.
Este sistemul
ce asigura energia contractiei musculare viguroase pe o perioada scurta sau
moderata.
Molecula de
glucoza este scindata in doua molecule de acid piruvic si din fiecare molecula
de glucoza libera rezulta 4 molecule de ATP. Acidul piruvic poate fi convertit
in acid lactic si sa nu necesite pe aceasta cale metabolism oxidativ. Acidul
lactic produce o stare de extrema oboseala, febra musculara si pe aceasta cale
limiteaza efortul. Este o supapa necesara de protectie caci efortul excesiv
poate ucide.
Ce inseamna 2000 kcal in termeni de efort fizic?
S-a demonstrat ca pentru eforturi prelungite cheltuiala energetica este
constanta de 1 kcal/kgc/km parcurs. Aceasta inseamna ca intr-o cursa de maraton
un individ de 70 kg cheltuieste 3000 kcal. Glucoza este extrem de importanta
atat pentru muschi cat si pentru celula nervoasa.
Celula nervoasa nu are rezerve de energie depuse in
celula, nu are depozite de glicogen (acestea sunt nesemnificative, de ordinul a
doua minute de functionare).
Dupa epuizarea rezervelor se produce un proces de
generare a glucidelor din celelalte principii chimice alimentare, asa zisa
neoglucogeneza.
Masina metabolica numita om are un randament ce este in functie de
carburantii sai biochimici. Eficienta energetica a metabolismului este de 25%.
Restul energiei chimice se pierde sub forma de caldura. Antrenamentul creste
aptitudinea muschilor de a utiliza acizi grasi deci si randamentul.
Desi proteinele reprezinta
componenta structurala cea mai importanta a organismului uman dupa apa, 15% din
greutatea corporala, si desi ele sunt dispuse mai ales in muschiul striat ele
sunt pentru muschi un element structural si nu un carburant.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 3868
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2026 . All rights reserved