| CATEGORII DOCUMENTE |
| Alimentatie nutritie | Asistenta sociala | Cosmetica frumusete | Logopedie | Retete culinare | Sport |
| Fitness |
Bazele biologice si psihologice ale vitezei
1 Cooperarea neuro-musculara si contractibilitate musculara
Frecventa rapida a muschilor nu se poate obtine fara alternarea ultrarapida a excitatiei si inhibitiei, cuplate cu reactia corespondenta a sistemului neuromuscular si in relatie cu efortul optim (Harre, 1976).
Numai o coordonare intra si intermusculara optimala permite cooperarea muschilor agonisti si antagonisti, marimea activarii unitatilor motrice. Acestea permit cresterea fortei musculaturii implicate in miscare.
Viteza excitatie-contractie, adica viteza de reactie a musculaturii declansate de sistemul nervos central, este in general denumita viteza de baza (Weineck, 1993). Dezvoltarea ei variaza de la un individ la altul, in functie de deosebirile ce afecteaza sistemul nervos al fiecaruia si depinde de marimea si descarcarea electrica a celulelor ganglionare.
Formarea de inalte frecvente de impulsie este o problema de exercitii precoce care permit spatiul genetic limitat sa se largeasca inainte ca dezvoltarea completa a sistemului nervos central sa se termine (Blaser, 1978), aceasta pentru ca se pare ca el este izolat intr-un cadru redus (Israel, 1977). Cu toate acestea, mai tarziu exista totusi posibilitatea de a influenta proprietatea de contractie a muschilor (Bhrle si Schmidtbleicher, 1981). Absaliamov, Zorin si Koz (1976) demonstreaza ca sprinterii si saritorii de performanta au contractii musculare foarte rapide, dar remarca faptul ca in cazul probelor aciclice (garduri), proprietatile de contractie ale aceluiasi muschi sunt diferite de la un picior la altul, acesta, desigur, datorandu-se schimbarilor produse prin antrenamentul probei respective.
2. Elasticitatea musculara si tipul de fibre musculare
Daca elasticitatea, limita de intindere si limita de destindere a muschiului, nu sunt suficiente, se va produce o reducere in amplitudinea gestului (Harre, 1976) si o deteriorare a cooperarii neuromusculare si a coordonarii, pentru ca muschii care lucreaza (agonisti) trebuie sa invinga, in timpul unei miscari, o rezistenta ridicata din partea antagonistilor. Miscarile sunt incetinite din cauza frictiunii interne si a unui tonus muscular ridicat, acesta necesitand nu numai o crestere a cheltuielilor energetice (diminuarea eficacitatii), dar provoaca intr-un timp foarte scurt si reducerea vitezei.
Viteza de contractie a muschiului depinde intr-o mare masura de tipul de fibre musculare care il compun. Cercetarile efectuate de Karlsson si colab. (1975), de Coyle si colab. (1974) si Inbar, Kaiser si Tesh (1981) au demonstrat, folosind biopsia, ca exista o corelatie stransa intre procentajul de fibre rapide ce compun un muschi si viteza de miscare. "Sprinterul" innascut poseda un procentaj mai ridicat de fibre rapide decat un alergator de fond, spre exemplu. Urmand un antrenament specific pentru dezvoltarea vitezei si a fortei, sectiunea transversala a fibrelor musculare poate sa creasca peste 25% din forta izometrica maxima (Karlsson si colab., 1975). Astfel, se produce o solicitare selectiva a fibrelor rapide, deci o marire a volumului lor.
FIBRELE RAPIDE, in abreviere FT (Fast Twitch = secusa rapida), denumite, de asemenea, fibre albe sau pale, sunt groase in diametru si se contracta rapid.
FIBRELE LENTE sau rosii, in aceeasi abreviere ST (Slow Twitch = secusa lenta), sunt mult mai fine si mult mai lente. Acest tip de fibre sunt implicate in special in actiunile musculare lente, de lunga durata si cu o intensitate relativ scazuta.
Aceste doua tipuri de fibre sunt diferite intre ele datorita solicitarilor functionale. Fibrele rapide sunt bogate in fosfati, in energie si in rezerve de glicogen, ele continand o mare concentratie de enzime din metabolismul energetic anaerob. La randul lor, fibrele lente, cu toate ca au o rezerva sensibil egala de glicogen, se disting printr-o mare concentratie mult mai mare de enzime din metabolismul aerob.
In fibrele lente, raportul citoplasmomitocondrial este superior mitocondrianelor si, pentru aceasta, se observa o activitate marita a enzimelor din ciclul de acid citric si o degradare in cazul acidului gras liber, deci activitatea enzimelor din glicoliza anaeroba este mai slaba (Keul, Doll si Keppler, 1969).
Ceea ce pare determinant in diferentele ce existau intre fibrele rapide si cele lente, este modul de interventie a sistemului nervos central. Inervatia, in cazul fibrelor lente, este realizata prin mici motoneuroni alfa cu transmitere relativ lenta, care transmit impulsii nervoase continue. Fibrele rapide sunt controlate prin motoneuroni alfa cu transmitere rapida, caracterizati, printr-un influx nervos intrerupt, tipic pentru actiunile motrice de mare intensitate, dar de scurta durata (Burke si Edgerton, 1975; Wittekopf, Marhold si Pieper, 1981).
Dispunerea si proportia fibrelor musculare pare a fi determinata genetic. La mare majoritate a populatiei procentajele sunt relativ aceleasi, cu toate ca repartitia lor poate sa fie 90/10 sau 10/90, in raportul fibre rapide/fibre lente (Weineck, 1993).
La un sprinter "innascut" fibrele rapide sun predominante, pe cand la un individ "innascut" maratonist fibrele lente sunt cele ce domina. Putem, deci, presupune ca tendinta inconstienta de a se dirija sportivii spre anumite probe in care predomina "explozivitatea" sau rezistenta poate fi pusa in raport cu repartitia ereditara a acestor fibre (Saltin 1973). Antrenamentul sportiv nu pare a modifica repartitia fibrelor rapide si lente, ci numai gradul lor de dezvoltare si volumul lor (Slatin, 1973; Karlsson si colab., 1975).
Consideram edificatoare o scurta sinteza privind unele aspecte referitoare la tipurile de fibre musculare si caracteristicile lor, din lucrarea lui Saltin si Henricksson (1980), citati de T. Ardelean (1991).
Denumirile utilizate pentru desemnarea diferitelor tipuri de fibre musculare sunt destul de variate. Noi am vorbit de existenta fibrelor lente, tonice si a fibrelor rapide, fazice, care, dupa coloratie, sunt rosii si respectiv albe, in functie de continutul lor de mioglobina (T. Ardelean, 1991).
Activitatea enzimelor mitocondriale este cu atat mai mare cu cat continutul muschiului in mioglobina este mai crescut, iar continutul in enzime sarcoplasmice, glicolitice, este invers proportional cu continutul in enzime oxidative si in mioglobina (Saltin, 1973; Henricksson, 1980 citati de T. Ardelean, 1991). Asadar vom denumi fibrele de contractie lenta fibre de tip I, iar pe cele de contractie rapida - fibre de tip II. Dupa cum mentioneaza autorii mai sus citati, la nivelul diferitelor specii exista posibilitatea de a efectua o subdiviziune in cadrul fibrelor de tip II (dupa T. Ardelean, 1991):
cu potential relativ slab.
La om, aceasta subdiviziune se datoreaza diferentei de sensibilitate fata de pH-ul acid, in sensul ca primele sunt inactive si isi pierd, deci, coloratia la un pH de 4.6 - 4.8 (tip IIA), pe cand celelalte isi pastreaza coloratia pana la un pH de 4.5 (tip IIB). Fibrele de tip IIA sunt considerate ca intermediare.
Caracteristicile acestor tipuri de fibre sunt prezentate in tabelul urmator:
|
NOMENCLATURA |
BAZA DE IDENTIFICARE |
||||
|
mioglobina |
enzime mitocondriale |
enzime glicolitice |
lipide |
ATP-aza miofibriliara |
|
|
tip I |
rosu |
puternic oxidativ |
slab |
concentratie puternica |
contractie lenta |
|
tip IIA |
intermediar |
oxidativ |
glicolitic |
concentratie intermediara |
contractie rapida |
|
tip IIB |
alb |
oxidativ |
puternic glicolitic |
concentratie slaba |
contractie rapida |
Tabel
nr. 2 (dupa T. Ardelean, 1991)
In cazul subiectului sedentar este posibil sa se puna in evidenta diferentele caracteristice privind metabolismul la nivelul metabolismul la nivelul celor doua tipuri de fibre.
Cercetarile efectuate pe grupe de barbati (70) si femei (45) neselectionati au putut stabili o proportie medie de:
fibre de tip I - 52%;
In schimb, repartitia diferitelor fibre, proportia lor, prezinta diferente individuale destul de semnificative, ceea ce face posibila afirmatia ca factorul genetic, care poate fi interpretat ca talent pentru sporturile de viteza, consta in cantitatea mare, existenta initial, de fibre de tip IIB.
Studiile efectuate pe muschii sportivilor de performanta au prilejuit constatarea conform careia alergatorii de viteza prezinta o proportie crescuta de fibre de tip II la nivelul membrelor inferioare. Asadar, structura muschiului, corespunzatoare celor doua categorii de fibre, este programata genetic (Harre si Hauptman, 1987, citati de T. Ardeleanu, 1991).
Harre si Hauptman (1987), citati de T. Ardelean (1991) considera cert faptul ca procentajul de fibre rapide poate sa creasca, in anumite limite, pana la pubertate daca se aplica stimuli de antrenament oportun.
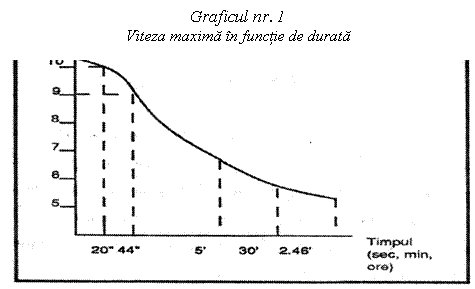
Viteza maxima a
individului depinde in primul rand de rezervele de energie a muschilor
implicati in efort si de viteza lor de mobilizare.
De altfel, viteza (a nu se intelege viteza maxima) se gaseste in stransa corelatie cu tipul de energie mobilizat. Ea atinge cota maxima spre 20 secunde din cursa (aproximativ 200m), ceea ce corespunde cu metabolizarea componentelor de fosfati bogate in energie, respectiv ATP-ul si CP-ul. Ea poate, dupa aceea, sa se mentina la un inalt nivel, in timpul efortului maximal, pana la aproximativ 50 sec. (ceea ce corespunde cu proba de 400m) si unde glicoza anaeroba se gaseste la punctul sau maxim. In final, ea se reduce sensibil odata cu marirea duratei si instalarea progresiva a mecanismelor aerobe de aprovizionare cu energie.
Rezervele componentelor de fosfati bogate in energie, in special creatin fosfatul - CP (Pansold, 1973), asa cum si rezervele de glicogen muscular si glicoza anaeroba pot fi imbunatatite printr-un antrenament specific. In paralel, creste activitatea enzimelor implicate in mobilizarea acestor substante energetice (Weineck, 1993).
3. ATP-ul sursa de energie pentru contractia musculara
Ori de cate ori se contracta un muschi, invingand o rezistenta, se produce un lucru mecanic, deci este necesara o anumita cantitate de energie. S-a constatat ca in timpul contractiei musculare sunt descompuse mari cantitati de ATP. Secventa evenimentelor considerate a fi declansatoare pentru contractia musculara se presupune, dupa Guyton (1988), a fi urmatoarele:
inaintea initierii contractiei capetele puntilor de miozina se leaga cu ATP-ul. Activitatea ATP-azica a capului miozinic desface ATP-ul in ADP si P care raman legate la cap. In acest stadiu, capul puntii se extinde perpendicular spre filamentul de actina, fara a fi atasat acestuia.
cand efectul inhibitor al troponinei si tropomiozinei este anulat de catre ionii de calciu, partile active ale actinei sunt descoperite si capetele puntilor de miozina se leaga cu actina.
aceste legaturi determina o schimbare de conformatie in sensul inclinarii capului la alt unghi cu ajutorul energiei stocate in capul puntii.
odata ce capul puntii este inclinat, se produce o eliberare de ADP si P si se creeaza un loc pentru ca un nou stoc de ATP sa se fixeze. Aceasta legare detaseaza capul puntii miozinice de pe filamentul de actina.
dupa detasarea capului de pe filamentul de actina noua molecula de ATP este hidrolizata, iar energia eliberata reduce capul in pozitie perpendiculara, gata sa inceapa un nou ciclu de acrosare.
aceste faze se produc continuu pana cand filamentele de actina trag membrana Z catre extremitatile filamentelor de miozina sau pana cand ancarcatura asupra muschiului devine prea mare pentru o scurtare suplimentara.
Cele doua tipuri de fibre existente in structura musculaturii striate sunt diferentiate intre ele si prin criterii metabolice. Astfel, fibrele ST se caracterizeaza printr-un metabolism predominant aerob, in acest sens pledand si continutul mitocondrial bogat si continutul bogat in enzime respiratorii. A doua categorie de fibre, cele FT, au metabolismul predominant anaerob, in special glicolitic, cu acumulare de acid lactic.
Eforturile sportive presupun contractii musculare care rezulta din transformarea energiei chimice in energie mecanica, la nivelul musculaturii active.
Definim capacitate de efort fizic posibilitatile sistemului muscular activ de a elibera prin glicoza anaeroba sau fosforilare oxidativa energia necesara pentru producerea unui lucru mecanic cat mai inalt posibil si mentinerea acestuia un timp cat mai indelungat posibil.
In compozitia biochimica a muschiului se gasesc substante de tip glucidic, protidic si lipidic, toate avand rol energetic pentru contractia musculara.
Sursa directa folosita de fibra musculara pentru furnizarea de energie este ATP-ul; rezervele de ATP pot sa asigure o energie pentru 4-6 contractii musculare care dureaza circa 6 secunde. Muschiul contine aproximativ 2 milimoli ATP/gram si poate genera prin hidroliza 11,5 kcal/mol.
Pentru refacerea ATP este folosita energia rezultata din scindarea CP la nivel muscular, care, poate astfel asigura energie pentru alte 15-20 contractii musculare, adica pentru aproximativ alte 20 secunde.
Fosfocreatina, in cantitate de 20 milimoli/gram, poseda si ea legaturi macroergice. Este descompusa instantaneu, iar energia eliberata produce legarea ionului fosfat la molecula de ADP si resinteza ATP-ului. Creatinfosfatul poate fosforila ADP-ul pentru a forma ATP-ul intr-o reactie reversibila, catalizata de enzima creatinchinaza.
CP - ADP (creatinchinaza) = creatina + ATP
Daca efortul continua, energia necesara este eliberata de glicogenul muscular (acesta ajuta regenerarea fosfatilor macroergici) care poate acoperi efortul din punct de vedere energetic pentru circa 45-60 sec., in lipsa de oxigien - glicoza anaeroba. In consecinta, rezulta cantitati relativ mari de acid lactic a carui crestere exagerata induce o scadere a pH-ului sanguin si consecutiv o inhibitie a glicozei.
Cea mai mare parte a energiei, circa 95%, este eliberata in timpul oxidarii finale a glucozei, proces care are loc in mitocondrii. In acest proces, piruvatul rezultat din glicoza este convertit in acetylcoenzima A si apoi oxidat pana la CO2 si H2O in ciclul acizilor tricarboxilici (ciclul lui Krebs). Pe aceasta cale oxidativa se genereaza energie inmagazinata in 38 moli ATP.
O mica parte din energie este eliberata prin degradarea glicogenului si glucozei, in anaerobioza, prin procesul de glicoza (calea Embden-Mayerhoff). Pe aceasta cale se formeaza 2 moli Atp/mmol de glucoza metabolizata.
In prezenta O2 acidul lactic este metabolizat si folosit chiar ca sursa energetica la nivelul miocardului, ficatul rinichilor. Rezulta deci, ca in stransa corelatie cu consumul (glicoza aeroba) si, astfel, efortul fizic poate continua.
4. Forta musculara si parametrii antropometrici
Diferitele nivele de performanta obtinute in domeniul vitezei depind de nivelul initial diferit a capacitatii de coordonare si de forta. Cresterea fortei duce la imbunatatirea vitezei, fapt explicat prin marirea sectiunii transversale a muschiului care permite o proliferare a punctelor de agatare in unitatea de timp, intre actina si miozina ce compun fibrele musculare, si din aceasta cauza, maresc viteza de interpenetratie (alunecarea uneia in cealalta) a filamentelor de actina si miozina, deci maresc viteza de contractie a muschiului. Marirea diametrului fibrelor musculare care compun unitatile motrice activate sincronic, duce in paralel la o reducere a cantitatii de lucru pe unitate, deci o contractie mult mai rapida (Paerish, 1974). Marirea fortei de propulsie influenteaza foarte mult lungimea si frecventa fuleului de alergare. Daca forta de impulsie este mai mare in faza de sprijin pe sol, lungimea fuleului creste, iar timpul de sprijin se scurteaza, ducand deci spre o marire a frecventei. Forta de impingere dinamica, legata de capacitatea de coordonare, determina, nemijlocit, viteza in alergare.
Variatia care exista intre lungimea si frecventa fuleului este asa de mare incat raportul intre talie si muschii ridicatori ai coapsei nu poate fi decisiv.
Letzeler si colab. (1979) ajung, in urma unor cercetari, la urmatoarele concluzii:
barbatii sunt in mod semnificativ superiori femeilor in ceea ce priveste lungimea fuleului, aceasta datorita taliei superioare;
proportional cu talia, barbatii si femeile au fuleuri aproximativ egale (atat pentru alergatorii lenti, cat si pentru cei rapizi);
femeile obtin, in medie, frecvente asemanatoare cu cele ale barbatilor.
Comparatia intre cei mai buni sprinteri si sprintere din lume permite sa constatam ca frecventa relativa a fuleului este factorul determinant al performantei, inaintea lungimii fuleului. Frecventa este deci, obiectivul prioritar in antrenamentul de viteza: din pacate acesta este confirmat de statistica doar in cazul barbatilor (Weineck, 1993).
5. Implicarea planului psihic
Calitatile motrice sunt mediate de procese, capacitati si stari psihice. Dintre factorii planului psihic al calitatii motrice viteza mentionam: capacitatea de mobilizare psihica in general si capacitatea de concentrare a atentiei; praguri absolute ridicate, cu mare putere de discriminare a analizatorilor implicati in senzatia de inaintare si, implicit, perceperea reala a vitezei miscarilor, respectiv aprecierea corecta a vitezei de deplasare (analizatorul tactil, chinestezic); echilibrul afectiv manifestat prin capacitatea de conservare a comportamentului motor specific in prezenta unor factori de stres (T. Ardelean, 1991).
Prestatia sprinterului este in mare masura influentata de coordonare, respectiv de reglarea nuro-musculara. Daca aceasta aptitudine nu este dezvoltata suficient, anumite circumstante pot perturba reglarea provocata de sistemul nervos central. Aceasta explica de ce atunci cand un sprinter, dupa ce este depasit de adversari, nu ramane numai in urma, dar in acelasi timp pierde si din viteza. Aptitudinea sa de coordonare se darama in momentul in care este depasit (J. Weineck, 1993).
O separare a miscarilor se produce in timpul alergarii (Koitzsch, 1972). Ter Oranesian (1971) explica acest fenomen universal cunoscut in practica prin faptul ca incercarea de a controla voluntar miscarile automatizate antreneaza in mod inevitabil deteriorarea coordonarii. Este momentul in care se instaureaza o dezautomatizare, ea fiind mult mai rapida in cazul celor ce nu au fixate miscarile.
Pe de alta parte, efortul maxim de vointa este decisiv in obtinerea unei viteze maxime (Harre, 1976). In aceeasi ordine de idei, Grosser vorbeste de asa-numitul punch voluntar. Acest efort de vointa nu influenteaza desfasurarea miscarii, insa influenteaza mult capacitatea interna de mobilizare (J. Weineck, 1993).
Mai definim ca factori ai planului psihic:
atitudinea fata de dificultatile antrenamentului si concursului;
calitatea proceselor volitive.
Capacitatea de automotivare are un rol central in obtinerea prestatiilor de rapiditate, pentru ca individul decide in maniera determinanta gradul de exploatare a potentialului biologic (T. Ardelean 1991).
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 2708
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved