| CATEGORII DOCUMENTE |
| Bulgara | Ceha slovaca | Croata | Engleza | Estona | Finlandeza | Franceza |
| Germana | Italiana | Letona | Lituaniana | Maghiara | Olandeza | Poloneza |
| Sarba | Slovena | Spaniola | Suedeza | Turca | Ucraineana |
Neurotransmission and the Central Nervous System
Overview
|
Drugs that act upon the central nervous system (CNS) influence the lives of everyone, every day. These agents are invaluable therapeutically because they can produce specific physiological and psychological effects. Without general anesthetics, modern surgery would be impossible. Drugs that affect the CNS can selectively relieve pain, reduce fever, suppress disordered movement, induce sleep or arousal, reduce the desire to eat, or allay the tendency to vomit. Selectively acting drugs can be used to treat anxiety, mania, depression, or schizophrenia and do so without altering consciousness (see Chapters 19: Drugs and the Treatment of Psychiatric Disorders: Depression and Anxiety Disorders and 20: Drugs and the Treatment of Psychiatric Disorders: Psychosis and Mania). The nonmedical self-administration of CNS-active drugs is a widespread practice. Socially acceptable stimulants and antianxiety agents produce stability, relief, and even pleasure for many. However, the excessive use of these and other drugs also can affect lives adversely when their uncontrolled, compulsive use leads to physical dependence on the drug or to toxic side effects, which may include lethal overdosage (see Chapter 24: Drug Addiction and Drug Abuse). The unique quality of drugs that affect the nervous system and behavior places investigators who study the CNS in the midst of an extraordinary scientific challengethe attempt to understand the cellular and molecular basis for the enormously complex and varied functions of the human brain. In this effort, pharmacologists have two major goals: to use drugs to elucidate the mechanisms that operate in the normal CNS and to develop appropriate drugs to correct pathophysiological events in the abnormal CNS. Approaches to the elucidation of the sites and mechanisms of action of CNS drugs demand an understanding of the cellular and molecular biology of the brain. Although knowledge of the anatomy, physiology, and chemistry of the nervous system is far from complete, the acceleration of interdisciplinary research on the CNS has led to remarkable progress. This chapter introduces guidelines and fundamental principles for the comprehensive analysis of drugs that affect the CNS. Specific therapeutic approaches to neurological and psychiatric disorders are discussed in the chapters that follow in this section (see Chapters 13: History and Principles of Anesthesiology, 14: General Anesthetics, 15: Local Anesthetics, 16: Therapeutic Gases: Oxygen, Carbon Dioxide, Nitric Oxide, and Helium, 17: Hypnotics and Sedatives, 18: Ethanol, 19: Drugs and the Treatment of Psychiatric Disorders: Depression and Anxiety Disorders, 20: Drugs and the Treatment of Psychiatric Disorders: Psychosis and Mania, 21: Drugs Effective in the Therapy of the Epilepsies, 22: Treatment of Central Nervous System Degenerative Disorders, 23: Opioid Analgesics, and 24: Drug Addiction and Drug Abuse). |
Organizational Principles of the Brain
|
The brain is an assembly of interrelated neural systems that regulate their own and each other's activity in a dynamic, complex fashion. Macrofunctions of Brain Regions The large anatomical divisions provide a superficial classification of the distribution of brain functions. Cerebral Cortex The two cerebral hemispheres constitute the largest division of the
brain. Regions of the cortex are classified in several ways: (1) by the modality
of information processed (e.g., sensory, including somatosensory,
visual, auditory, and olfactory, as well as motor and associational); (2) by
anatomical position (frontal, temporal, parietal, and occipital); and (3) by
the geometrical relationship between cell types in the major cortical layers
('cytoarchitectonic' classifications). The cerebral cortex exhibits
a relatively uniform laminar appearance within any given local region.
Columnar sets of approximately 100 vertically connected neurons are thought
to form an elemental processing module. The specialized functions of a
cortical region arise from the interplay upon this basic module of
connections among other regions of the cortex (corticocortical systems) and
noncortical areas of the brain (subcortical systems) (seeMountcastle,
1997). Varying numbers of adjacent columnar modules may be functionally, but
transiently, linked into larger information-processing ensembles. The pathology
of Alzheimer's disease, for example, destroys the integrity of the columnar
modules and the corticocortical connections (seeMorrison and These columnar ensembles serve to interconnect nested distributed systems in which sensory associations are rapidly modifiable as information is processed (seeMountcastle, 1997; Tononi and Edelman, 1998). Cortical areas termed association areas receive and somehow process information from primary cortical sensory regions to produce higher cortical functions such as abstract thought, memory, and consciousness. The cerebral cortices also provide supervisory integration of the autonomic nervous system, and they may integrate somatic and vegetative functions, including those of the cardiovascular and gastrointestinal systems. Limbic System The 'limbic system' is an archaic term for an assembly of brain regions (hippocampal formation, amygdaloid complex, septum, olfactory nuclei, basal ganglia, and selected nuclei of the diencephalon) grouped around the subcortical borders of the underlying brain core to which a variety of complex emotional and motivational functions have been attributed. Modern neuroscience avoids this term, because the components of the limbic system neither function consistently as a system nor are the boundaries of such a system precisely defined. Parts of the limbic system also participate individually in functions that are capable of more precise definition. Thus, the basal ganglia or neostriatum (the caudate nucleus, putamen, globus pallidus, and lentiform nucleus) form an essential regulatory segment of the so-called extrapyramidal motor system. This system complements the function of the pyramidal (or voluntary) motor system. Damage to the extrapyramidal system depresses the ability to initiate voluntary movements and causes disorders characterized by involuntary movements, such as the tremors and rigidity of Parkinson's disease or the uncontrollable limb movements of Huntington's chorea (seeChapter 22: Treatment of Central Nervous System Degenerative Disorders). Similarly, the hippocampus may be crucial to the formation of recent memory, since this function is lost in patients with extensive bilateral damage to the hippocampus. Memory also is disrupted with Alzheimer's disease, which destroys the intrinsic structure of the hippocampus as well as parts of the frontal cortex (see also Squire, 1998). Diencephalon The thalamus lies in the center of the brain, beneath the cortex and basal ganglia and above the hypothalamus. The neurons of the thalamus are arranged into distinct clusters, or nuclei, which are either paired or midline structures. These nuclei act as relays between the incoming sensory pathways and the cortex, between the discrete regions of the thalamus and the hypothalamus, and between the basal ganglia and the association regions of the cerebral cortex. The thalamic nuclei and the basal ganglia also exert regulatory control over visceral functions; aphagia and adipsia, as well as general sensory neglect, follow damage to the corpus striatum or to selected circuits ending there (seeJones, 1998). The hypothalamus is the principal integrating region for the entire autonomic nervous system, and, among other functions, it regulates body temperature, water balance, intermediary metabolism, blood pressure, sexual and circadian cycles, secretion of the adenohypophysis, sleep, and emotion. Recent advances in the cytophysiological and chemical dissection of the hypothalamus have clarified the connections and possible functions of individual hypothalamic nuclei (Swanson, 1999). Midbrain and Brainstem The mesencephalon, pons, and medulla oblongata connect the cerebral hemispheres and thalamus-hypothalamus to the spinal cord. These 'bridge portions' of the CNS contain most of the nuclei of the cranial nerves, as well as the major inflow and outflow tracts from the cortices and spinal cord. These regions contain the reticular activating system, an important but incompletely characterized region of gray matter linking peripheral sensory and motor events with higher levels of nervous integration. The major monoamine-containing neurons of the brain (see below) are found here. Together, these regions represent the points of central integration for coordination of essential reflexive acts, such as swallowing and vomiting, and those that involve the cardiovascular and respiratory systems; these areas also include the primary receptive regions for most visceral afferent sensory information. The reticular activating system is essential for the regulation of sleep, wakefulness, and level of arousal as well as for coordination of eye movements. The fiber systems projecting from the reticular formation have been called 'nonspecific,' because the targets to which they project are relatively more diffuse in distribution than those of many other neurons (e.g., specific thalamocortical projection). However, the chemically homogeneous components of the reticular system innervate targets in a coherent, functional manner despite their broad distribution (seeFoote and Aston-Jones, 1995; Usher et al., 1999). Cerebellum The cerebellum arises from the posterior pons behind the cerebral hemispheres. It is also highly laminated and redundant in its detailed cytological organization. The lobules and folia of the cerebellum project onto specific deep cerebellar nuclei, which in turn make relatively selective projections to the motor cortex (by way of the thalamus) and to the brainstem nuclei concerned with vestibular (position-stabilization) function. In addition to maintaining the proper tone of antigravity musculature and providing continuous feedback during volitional movements of the trunk and extremities, the cerebellum also may regulate visceral function (e.g., heart rate, so as to maintain blood flow despite changes in posture). In addition, the cerebellum has been shown in recent studies to play a significant role in learning and memory (seeMiddleton and Strick, 1998). Spinal Cord The spinal cord extends from the caudal end of the medulla oblongata to the lower lumbar vertebrae. Within this mass of nerve cells and tracts, the sensory information from skin, muscles, joints, and viscera is locally coordinated with motoneurons and with primary sensory relay cells that project to and receive signals from higher levels. The spinal cord is divided into anatomical segments (cervical, thoracic, lumbar, and sacral) that correspond to divisions of the peripheral nerves and spinal column. Ascending and descending tracts of the spinal cord are located within the white matter at the perimeter of the cord, while intersegmental connections and synaptic contacts are concentrated within the H-shaped internal mass of gray matter. Sensory information flows into the dorsal cord, and motor commands exit via the ventral portion. The preganglionic neurons of the autonomic nervous system (seeChapter 6: Neurotransmission: The Autonomic and Somatic Motor Nervous Systems) are found in the intermediolateral columns of the gray matter. Autonomic reflexes (e.g., changes in skin vasculature with alteration of temperature) easily can be elicited within local segments of the spinal cord, as shown by the maintenance of these reflexes after the cord is severed. Microanatomy of the Brain Neurons operate either within layered structures (such as the olfactory bulb, cerebral cortex, hippocampal formation, and cerebellum) or in clustered groupings (the defined collections of central neurons that aggregate into nuclei). The specific connections between neurons within or across the macrodivisions of the brain are essential to the brain's functions. It is through their patterns of neuronal circuitry that individual neurons form functional ensembles to regulate the flow of information within and between the regions of the brain. Cellular Organization of the Brain Present understanding of the cellular organization of the CNS can be viewed simplistically according to three main patterns of neuronal connectivity (seeShepherd, 1998). Long-hierarchical neuronal connections typically are found in the primary sensory and motor pathways. Here the transmission of information is highly sequential, and interconnected neurons are related to each other in a hierarchical fashion. Primary receptors (in the retina, inner ear, olfactory epithelium, tongue, or skin) transmit first to primary relay cells, then to secondary relay cells, and finally to the primary sensory fields of the cerebral cortex. For motor output systems, the reverse sequence exists with impulses descending hierarchically from motor cortex to spinal motoneuron. This hierarchical scheme of organization provides a precise flow of information, but such organization suffers the disadvantage that destruction of any link incapacitates the entire system. Local-circuit neurons establish their connections mainly within their immediate vicinity. Such local-circuit neurons frequently are small and may have very few processes. They are believed to regulate (i.e., expand or constrain) the flow of information through their small spatial domain. Given their short axons, they may function without generating action potentials, which are essential for the long-distance transmission between hierarchically connected neurons. The neurotransmitters for many local-circuit neurons in most brain regions have been inferred through pharmacological tests (see below). Single-source divergent circuitry is utilized by certain neuronal systems of the hypothalamus, pons, and medulla. From their clustered anatomical location, these neurons extend multiple-branched and divergent connections to many target cells, almost all of which lie outside the brain region in which the neurons are located. Neurons with divergent circuitry can be conceived of as special local-circuit neurons whose spatial domains are one to two orders of magnitude larger than those of the classical intraregional interneurons rather than as sequential elements within any known hierarchical system. For example, neurons of the locus ceruleus project from the pons to the cerebellum, spinal cord, thalamus, and several cortical zones, whose function is only subtly disrupted when the adrenergic fibers are destroyed experimentally. Abundant data suggest that these systems could mediate linkages between regions that may require temporary integration (seeFoote and Aston-Jones, 1995; Aston-Jones et al., 1999). The neurotransmitters for some of these connections are well known (see below), while others remain to be identified. Cell Biology of Neurons Neurons are classified in many different ways, according to function ( sensory, motor, or interneuron ), location, or identity of the transmitter they synthesize and release. Microscopic analysis focuses on their general shape and, in particular, the number of extensions from the cell body. Most neurons have one axon to carry signals to functionally interconnected target cells. Other processes, termed dendrites, extend from the nerve cell body to receive synaptic contacts from other neurons; these dendrites may branch in extremely complex patterns. Neurons exhibit the cytological characteristics of highly active secretory cells with large nuclei; large amounts of smooth and rough endoplasmic reticulum; and frequent clusters of specialized smooth endoplasmic reticulum (Golgi apparatus), in which secretory products of the cell are packaged into membrane-bound organelles for transport out of the cell body proper to the axon or dendrites (Figure 121). Neurons and their cellular extensions are rich in microtubuleselongated tubules approximately 24 nm in diameter. Their functions may be to support the elongated axons and dendrites and to assist in the reciprocal transport of essential macromolecules and organelles between the cell body and the distant axon or dendrites.
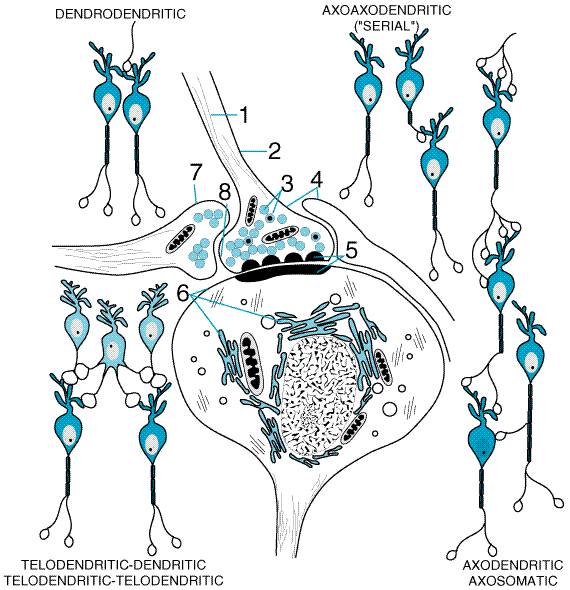
The sites of interneuronal communication in the CNS are termed synapses (see below). Although synapses are functionally analogous to 'junctions' in the somatic motor and autonomic nervous systems, the central junctions are characterized morphologically by various additional forms of paramembranous deposits of specific proteins (essential for transmitter release, response, and catabolism; seeLiu and Edwards, 1997; Geppert and Sdhof, 1998). These specialized sites are presumed to be the active zone for transmitter release and response. The paramembranous proteins constitute a specialized junctional adherence zone, termed the synaptolemma(seeBodian, 1972). Like peripheral 'junctions,' central synapses also are denoted by accumulations of tiny (500 to 1500 ) organelles, termed synaptic vesicles. The proteins of these vesicles have been shown to have specific roles in transmitter storage, vesicle docking onto presynaptic membranes, voltage- and Ca2+-dependent secretion (seeChapter 6: Neurotransmission: The Autonomic and Somatic Motor Nervous Systems), and recycling and restorage of released transmitter (seeAugustine et al., 1999). Synaptic Relationships Synaptic arrangements in the CNS fall into a wide variety of morphological and functional forms that are specific for the cells involved. Many spatial arrangements are possible within these highly individualized synaptic relationships (seeFigure 121). The most common arrangement, typical of the hierarchical pathways, is the axodendritic or axosomatic synapse in which the axons of the cell of origin make their functional contact with the dendrites or cell body of the target. In other cases, functional contacts may occur more rarely between adjacent cell bodies (somasomatic) or overlapping dendrites (dendrodendritic). Some local-circuit neurons can enter into synaptic relationships through modified dendrites, telodendrites, that can be either presynaptic or postsynaptic. Within the spinal cord, serial axoaxonic synapses are relatively frequent. Here, the axon of an interneuron ends on the terminal of a long-distance neuron as that terminal contacts a dendrite in the dorsal horn. Many presynaptic axons contain local collections of typical synaptic vesicles with no opposed specialized synaptolemma (termed boutons en passant). Release of transmitter may not occur at such sites. The bioelectric properties of neurons and junctions in the CNS generally follow the outlines and details already described for the peripheral autonomic nervous system (seeChapter 6: Neurotransmission: The Autonomic and Somatic Motor Nervous Systems). However, in the CNS there is found a much more varied range of intracellular mechanisms (Nicoll et al., 1990; Tzounopoulos et al., 1998). Supportive Cells Neurons are not the only cells in the CNS. According to most estimates, neurons are outnumbered, perhaps by an order of magnitude, by the various nonneuronal supportive cellular elements (seeCherniak, 1990). Nonneuronal cells include the macroglia, microglia, the cells of the vascular elements (including the intracerebral vasculature as well as the cerebrospinal fluid-forming cells of the choroid plexus found within the intracerebral ventricular system), and the meninges, which cover the surface of the brain and comprise the cerebrospinal fluid-containing envelope. Macroglia are the most abundant supportive cells; some are categorized as astrocytes (nonneuronal cells interposed between the vasculature and the neurons, often surrounding individual compartments of synaptic complexes). Astrocytes play a variety of metabolic support roles including furnishing energy intermediates and supplementary removal of excessive extracellular neurotransmitter secretions (seeMagistretti et al., 1995). A second prominent category of macroglia are the myelin-producing cells, the oligodendroglia. Myelin, made up of multiple layers of their compacted membranes, insulates segments of long axons bioelectrically and accelerates action-potential conduction velocity. Microglia are relatively uncharacterized supportive cells believed to be of mesodermal origin and related to the macrophage/ monocyte lineage (seeAloisi, 1999; Gonzlez-Scarano and Baltuch, 1999). Some microglia are resident within the brain, while additional cells of this class may be attracted to the brain during periods of inflammation following either microbial infection or other postinjury inflammatory reactions. The response of the brain to inflammation differs strikingly from that of other tissues (seeAndersson et al., 1992; Raber et al., 1998; Schnell et al., 1999) and may in part explain its unique reactions to trauma (see below). BloodBrain Barrier Apart from the exceptional instances in which drugs are introduced directly into the CNS, the concentration of the agent in the blood after oral or parenteral administration will differ substantially from its concentration in the brain. Although not thoroughly defined anatomically, the bloodbrain barrier is an important boundary between the periphery and the CNS in the form of a permeability barrier to the passive diffusion of substances from the bloodstream into various regions of the CNS (seePark and Cho, 1991; Rubin and Staddon, 1999). Evidence of the barrier is provided by the greatly diminished rate of access of chemicals from plasma to the brain (seeChapter 1: Pharmacokinetics: The Dynamics of Drug Absorption, Distribution, and Elimination). This barrier is much less prominent in the hypothalamus and in several small, specialized organs lining the third and fourth ventricles of the brain: the median eminence, area postrema, pineal gland, subfornical organ, and subcommissural organ. In addition, there is little evidence of a barrier between the circulation and the peripheral nervous system (e.g., sensory and autonomic nerves and ganglia). While severe limitations are imposed on the diffusion of macromolecules, selective barriers to permeation also exist for small charged molecules such as neurotransmitters, their precursors and metabolites, and some drugs. These diffusional barriers are at present best thought of as a combination of the partition of solute across the vasculature (which governs passage by definable properties such as molecular weight, charge, and lipophilicity) and the presence or absence of energy-dependent transport systems. Active transport of certain agents may occur in either direction across the barriers. The diffusional barriers retard the movement of substances from brain to blood as well as from blood to brain. The brain clears metabolites of transmitters into the cerebrospinal fluid by excretion through the acid transport system of the choroid plexus (seeCserr and Bundgaard, 1984; Strange, 1993). Substances that rarely gain access to the brain from the bloodstream often can reach the brain after injection directly into the cerebrospinal fluid. Under certain conditions, it may be possible to open the bloodbrain barrier, at least transiently, to permit the entry of chemotherapeutic agents (seeEmerich et al., 1998; Granholm et al., 1998; LeMay et al., 1998, for discussion). Cerebral ischemia and inflammation also modify the bloodbrain barrier, resulting in increased access to substances that ordinarily would not affect the brain. Response to Damage: Repair and Plasticity in the CNS Because the neurons of the CNS are terminally differentiated cells, they do not undergo proliferative responses to damage, although recent evidence suggests the possibility of neural stem-cell proliferation as a natural means for selected neuronal replacement (seeGage, 2000). As a result, neurons have evolved other adaptive mechanisms to provide for maintenance of function following injury. These adaptive mechanisms endow the brain with considerable capacity for structural and functional modification well into adulthood (seeYang et al., 1994; Jones et al., 2000), and they may represent some of the mechanisms employed in the phenomena of memory and learning (seeKandel and O'Dell, 1992). Recent studies have shown that molecular signaling processes employed during brain development also may be involved in the plasticity seen in the adult brain, relying on specific neurotrophic agents (seeBothwell, 1995; Casaccia-Bonnefil et al., 1998; Chao et al., 1998); see below). |
Integrative Chemical Communication in the Central Nervous System
|
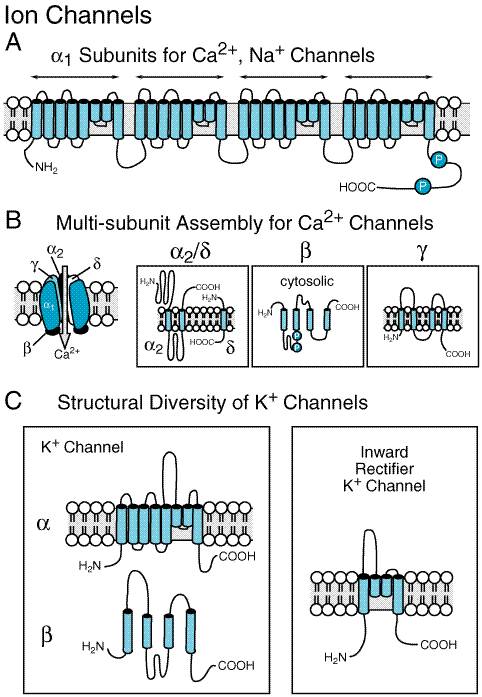
The capacity to integrate information from a variety of external and internal sources epitomizes the cardinal role of the CNS, namely to optimize the needs of the organism within the demands of the individual's environment. These integrative concepts transcend individual transmitter systems and emphasize the means by which neuronal activity is normally coordinated. Only through a detailed understanding of these integrative functions, and their failure in certain pathophysiological conditions, can effective and specific therapeutic approaches be developed for neurological and psychiatric disorders. The identification of molecular and cellular mechanisms of neural integration is productively linked to clinical therapeutics, because untreatable diseases and unexpected nontherapeutic side effects of drugs often reveal ill-defined mechanisms of pathophysiology. Such observations can then drive the search for novel mechanisms of cellular regulation. The capacity to link molecular processes to behavioral operations, both normal and pathological, provides one of the most exciting aspects of modern neuropharmacological research. A central underlying concept of neuropsychopharmacology is that drugs that influence behavior and improve the functional status of patients with neurological or psychiatric diseases act by enhancing or blunting the effectiveness of specific combinations of synaptic transmitter actions. Four research strategies provide the neuroscientific substrates of neuropsychological phenomena: molecular, cellular, multicellular (or systems), and behavioral. The intensively exploited molecular level has been the traditional focus for characterizing drugs that alter behavior. Molecular discoveries provide biochemical probes for identifying the appropriate neuronal sites and their mediative mechanisms. Such mechanisms include: (1) the ion channels, which provide for changes in excitability induced by neurotransmitters; (2) the neurotransmitter receptors (see below); (3) the auxiliary intramembranous and cytoplasmic transductive molecules that couple these receptors to intracellular effectors for short-term changes in excitability and for longer-term regulation e.g., through alterations in gene expression (seeNeyroz et al., 1993; Gudermann et al., 1997); (4) transporters for the conservation of released transmitter molecules by reaccumulation into nerve terminals, and then into synaptic vesicles (Blakely et al., 1994; Amara and Sonders, 1998; Fairman and Amara, 1999). Transport across vesicle membranes utilizes a transport protein distinct from that involved in reuptake into nerve terminals (Liu and Edwards, 1997). Research at the molecular level also provides the pharmacological tools to verify the working hypotheses of other molecular, cellular, and behavioral strategies and allows for a means to pursue their genetic basis. Thus, the most basic cellular phenomena of neurons now can be understood in terms of such discrete molecular entities. It has been known for some time that the basic excitability of neurons is achieved through modifications of the ion channels that all neurons express in abundance in their plasma membranes. However, it is now possible to understand precisely how the three major cations, Na+, K+, and Ca2+, as well as the Clanion are regulated in their flow through highly discriminative ion channels (seeFigures 122 and 123). The voltage-dependent ion channels (Figure 122), which are contrasted with the 'ligand-gated ion channels' (Figure 123), provide for rapid changes in ion permeability. These rapid changes underlie the rapid propagation of signals along axons and dendrites, and for the excitation-secretion coupling that releases neurotransmitters from presynaptic sites (Catterall, 1988, 1993). Cloning, expression, and functional assessment of constrained molecular modifications have defined conceptual chemical similarities among the major cation channels (seeFigure 122A). The intrinsic membrane-embedded domains of the Na+and Ca2+ channels are envisioned as four tandem repeats of a putative six-transmembrane domain, while the K+ channel family contains greater molecular diversity. X-ray crystallography has now confirmed these configurations for the K+ channel (Doyle et al., 1998). One structural form of voltage-regulated K+ channels, shown in Figure 122C, consists of subunits composed of a single putative six-transmembrane domain. The inward rectifier K+ channel structure, in contrast, retains the general configuration corresponding to transmembrane spans 5 and 6 with the interposed 'pore region' that penetrates only the exofacial surface membrane. These two structural categories of K+ channels can form heteroligomers, giving rise to multiple possibilities for regulation by voltage, neurotransmitters, assembly with intracellular auxiliary proteins, or posttranslational modifications (Krapivinsky et al., 1995). The structurally defined channel molecules (see Jan et al., 1997; Doyle et al., 1998) now can be examined to determine how drugs, toxins, and imposed voltages alter the excitability of a neuron, permitting a cell either to become spontaneously active or to die through prolonged opening of such channels (seeAdams and Swanson, 1994). Within the CNS, variants of the K+ channels (the delayed rectifier, the Ca2+-activated K+ channel, and the afterhyperpolarizing K+ channel) regulated by intracellular second messengers repeatedly have been shown to underlie complex forms of synaptic modulation (seeNicoll, et al., 1990; Malenka and Nicoll, 1999).
Research at the cellular level determines which specific neurons and which of their most proximate synaptic connections may mediate a behavior or the behavioral effects of a given drug. For example, research at the cellular level into the basis of emotion exploits both molecular and behavioral leads to determine the most likely brain sites at which behavioral changes pertinent to emotion can be analyzed. Such research provides clues as to the nature of the interactions in terms of interneuronal communication (i.e., excitation, inhibition, or more complex forms of synaptic interaction; seeAston-Jones et al., 1999; Brown et al., 1999). An understanding at the systems level is required to assemble the descriptive structural and functional properties of specific central transmitter systems, linking the neurons that make and release this transmitter to the possible effects of this release at the behavioral level. While many such transmitter-to-behavior linkages have been postulated, it has proven difficult to validate the essential involvement of specific transmitter-defined neurons in the mediation of specific mammalian behavior. Research at the behavioral level often can illuminate the integrative phenomena that link populations of neurons (often through operationally or empirically defined ways) into extended specialized circuits, ensembles, or more pervasively distributed systems that integrate the physiological expression of a learned, reflexive, or spontaneously generated behavioral response. The entire concept of animal models of human psychiatric diseases rests on the assumption that scientists can appropriately infer from observations of behavior and physiology (heart rate, respiration, locomotion, etc.) that the states experienced by animals are equivalent to the emotional states experienced by human beings expressing similar physiological changes (seeKandel, 1998). Identification of Central Transmitters An essential step in understanding the functional properties of neurotransmitters within the context of the circuitry of the brain is to identify which substances are the transmitters for specific interneuronal connections. The criteria for the rigorous identification of central transmitters require the same data used to establish the transmitters of the autonomic nervous system (seeChapter 6: Neurotransmission: The Autonomic and Somatic Motor Nervous Systems).
Other studies, especially those that have implicated peptides as
transmitters in the central and peripheral nervous systems, suggest that many
brain and spinal cord synapses contain more than one transmitter substance (seeHkfelt
et al., 2000). Although rigorous proof is lacking, substances that
coexist in a given synapse are presumed to be released together and to act
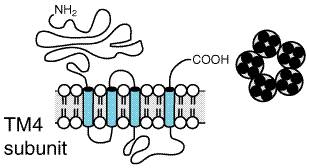
jointly on the postsynaptic membrane (seeDerrick and CNS Transmitter Discovery Strategies The earliest transmitters considered for central roles were acetylcholine and norepinephrine, largely because of their established roles in the somatic motor and autonomic nervous systems. In the 1960s, serotonin, epinephrine, and dopamine also were investigated as potential CNS transmitters. Histochemical as well as biochemical and pharmacological data yielded results consistent with roles as neurotransmitters, but complete satisfaction of all criteria was not achieved. In the early 1970s, the availability of selective and potent antagonists of gamma-aminobutyric acid (GABA), glycine, and glutamate, all known to be enriched in brain, led to their acceptance as transmitter substances in general. Also at this time, a search for hypothalamic-hypophyseal factors led to an improvement in the technology to isolate, purify, sequence, and synthetically replicate a growing family of neuropeptides (seeHkfelt, et al., 2000, for an overview). This advance, coupled with the widespread application of immunohistochemistry, strongly supported the view that neuropeptides may act as transmitters. Adaptation of bioassay technology from studies of pituitary secretions to other effectors (such as smooth-muscle contractility and, later, ligand-displacement assays) gave rise to the discovery of endogenous peptide ligands for drugs acting at opiate receptors (seeChapter 23: Opioid Analgesics). The search for endogenous factors whose receptors constituted the drug-binding sites was extended later to the benzodiazepine receptors (Costa and Guidotti, 1991). A more recent extension of this strategy has identified a series of endogenous lipid amides as the natural ligands for the tetrahydrocannabinoid receptors (seePiomelli et al., 1998). Assessment of Receptor Properties Until quite recently, central synaptic receptors were characterized either by examination of their ability to bind radiolabeled agonists or antagonists (and on the ability of other unlabeled compounds to compete for such binding sites) or by electrophysiological or biochemical consequences of receptor activation of neurons in vivo or in vitro. Radioligand-binding assays can quantify binding sites within a region, track their appearance throughout the phylogenetic scale and during brain development, and evaluate how physiological or pharmacological manipulation regulates receptor number or affinity (seeDumont et al., 1998; Redrobe et al., 1999, for examples). The properties of the cellular response to the transmitter can be studied electrophysiologically by the use of microiontophoresis (involving recording from single cells and highly localized drug administration). The patch-clamp technique can be used to study the electrical properties of single ionic channels and their regulation by neurotransmitters. These direct electrophysiological tests of neuronal responsiveness can provide qualitative and quantitative information on the effects of a putative transmitter substance (seeJardemark et al., 1998, for recent examples). Receptor properties also can be studied biochemically when the activated receptor is coupled to an enzymatic reaction, such as the synthesis of a second messenger and the ensuing biochemical changes measured. In the current era, molecular biological techniques have led to identification of mRNAs (or cDNAs) for the receptors for virtually every natural ligand considered as a neurotransmitter. A common practice is to introduce these coding sequences into test cells (frog oocytes or mammalian cells) and to assess the relative effects of ligands and of second-messenger production in such cells. Molecular cloning studies have revealed two major (seeFigures 123 and 124) and one minor molecular motif of transmitter receptors. Oligomeric ion-channel receptors composed of multiple subunits usually have four putative 'transmembrane domains' consisting of 20 to 25 generally hydrophobic amino acids (seeFigure 123). The ion channel receptors (called ionotropic receptors) for neurotransmitters contain sites for reversible phosphorylation by protein kinases and phosphoprotein phosphatases and for voltage-gating. Receptors with this structure include nicotinic cholinergic (or nicotinic acetylcholine) receptors (seeChapters 2: Pharmacodynamics: Mechanisms of Drug Action and the Relationship between Drug Concentration and Effect and 7: Muscarinic Receptor Agonists and Antagonists); the receptors for the amino acids GABA, glycine, glutamate, and aspartate, and for the 5-HT3 receptor (seeChapter 11: 5-Hydroxytryptamine (Serotonin): Receptor Agonists and Antagonists).
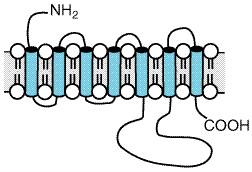
The second major structural motif for transmitter receptors is manifest by G proteincoupled receptors (GPCR), in which a monomeric receptor has seven putative transmembrane domains, with varying intra- and extracytoplasmic loop lengths (seeFigure 124). Multiple mutagenesis strategies have defined how the activated receptors (themselves subject to reversible phosphorylation at one or more functionally distinct sites) can interact with the heterotrimeric GTP-binding protein complex to ultimately activate, inhibit, or otherwise regulate enzymatic effector systems, e.g., adenylyl cyclase or phospholipase C, or ion channels, such as voltage-gated Ca2+ channels or receptor-operated K+ channels (seeFigure 21 and related text in Chapter 2: Pharmacodynamics: Mechanisms of Drug Action and the Relationship Between Drug Concentration and Effect). The GPCR family includes muscarinic cholinergic receptors, GABABand metabotropic glutamate receptors, and all other aminergic and peptidergic receptors. By transfecting 'null cells' with uncharacterized GPCR mRNAs, novel neuropeptides have been identified (seeReinscheid et al., 1995). A third receptor motif is represented by cell-surface receptors whose cytoplasmic domains possess catalytic activities, in particular, guanylyl cyclase (seeChapter 2: Pharmacodynamics: Mechanisms of Drug Action and the Relationship Between Drug Concentration and Effect). An additional molecular motif expressed within the CNS involves the transporters that remove transmitters after secretion by an ion-dependent reuptake process (Figure 125). Transporters exhibit a molecular motif with 12 hypothetical transmembrane domains similar to glucose transporters and to mammalian adenylyl cyclase (seeTang and Gilman, 1992).
Postsynaptic receptivity of CNS neurons is regulated continuously in terms of the number of receptor sites and the threshold required for generation of a response. Receptor number often is dependent on the concentration of agonist to which the target cell is exposed. Thus, chronic excess of agonist can lead to a reduced number of receptors (desensitization or down-regulation) and consequently to subsensitivity or tolerance to the transmitter. A deficit of transmitter can lead to increased numbers of receptors and supersensitivity of the system. These adaptive processes become especially important when drugs are used to treat chronic illness of the CNS. With prolonged periods of exposure to drug, the actual mechanisms underlying the therapeutic effect may differ strikingly from those that operate when the agent is first introduced into the system. Similar adaptive modifications of neuronal systems also can occur at presynaptic sites, such as those concerned with transmitter synthesis, storage, reuptake, and release. |
Neurotransmitters, Neurohormones, and Neuromodulators: Contrasting Principles of Neuronal Regulation
|
Neurotransmitters Satisfaction of the experimental criteria for identification of synaptic transmitters can lead to the conclusion that a substance contained in a neuron is secreted by that neuron to transmit information to its postsynaptic target. Given a definite effect of neuron A on target cell B, a substance found in or secreted by neuron A and producing the effect of A on B operationally would be the transmitter from A to B. In some cases, transmitters may produce minimal effects on bioelectric properties yet activate or inactivate biochemical mechanisms necessary for responses to other circuits. Alternatively, the action of a transmitter may vary with the context of ongoing synaptic eventsenhancing excitation or inhibition, rather than operating to impose direct excitation or inhibition (seeBourne and Nicoll, 1993). Each chemical substance that fits within the broad definition of a transmitter may, therefore, require operational definition within the spatial and temporal domains in which a specific cell-cell circuit is defined. Those same properties may or may not be generalized to other cells that are contacted by the same presynaptic neurons, with the differences in operation related to differences in postsynaptic receptors and the mechanisms by which the activated receptor produces its effect. Classically, electrophysiological signs of the action of a bona fide transmitter fall into two major categories: (1) excitation, in which ion channels are opened to permit net influx of positively charged ions, leading to depolarization with a reduction in the electrical resistance of the membrane; and (2) inhibition, in which selective ion movements lead to hyperpolarization, also with decreased membrane resistance. More recent work suggests there may be many 'nonclassical' transmitter mechanisms operating in the CNS. In some cases, either depolarization or hyperpolarization is accompanied by a decreased ionic conductance (increased membrane resistance) as actions of the transmitter lead to the closure of ion channels (so-called leak channels) that normally are open in some resting neurons (Shepherd, 1998). For some transmitters, such as monoamines and certain peptides, a 'conditional' action may be involved. That is, a transmitter substance may enhance or suppress the response of the target neuron to classical excitatory or inhibitory transmitters while producing little or no change in membrane potential or ionic conductance when applied alone. Such conditional responses have been termed modulatory, and specific categories of modulation have been hypothesized (seeNicoll et al., 1990; Foote and Aston-Jones, 1995). Regardless of the mechanisms that underlie such synaptic operations, their temporal and biophysical characteristics differ substantially from the rapid onset-offset type of effect previously thought to describe all synaptic events. These differences have thus raised the issue of whether or not substances that produce slow synaptic effects should be described with the same termneurotransmitter. Some of the alternative terms and the molecules they describe deserve brief mention with regard to mechanisms of drug action. Neurohormones Peptide-secreting cells of the hypothalamicohypophyseal circuits originally were described as neurosecretory cells, a form of neuron that was both fish and fowl, receiving synaptic information from other central neurons yet secreting transmitters in a hormone-like fashion into the circulation. The transmitter released from such neurons was termed a neurohormonei.e., a substance secreted into the blood by a neuron. However, this term has lost most of its original meaning, because these hypothalamic neurons also may form traditional synapses with central neurons (Hkfelt et al., 1995, 2000). Cytochemical evidence indicates that the same substance that is secreted as a hormone from the posterior pituitary (oxytocin, antidiuretic hormone), mediates transmission at these sites. Thus, the designation hormone relates to the site of release at the pituitary and does not necessarily describe all of the actions of the peptide. Neuromodulators Florey (1967) employed the term modulator to describe substances that can influence neuronal activity in a manner different from that of neurotransmitters. In the context of this definition, the distinctive feature of a modulator is that it originates from cellular and nonsynaptic sites, yet influences the excitability of nerve cells. Florey specifically designated substances such as CO2and ammonia, arising from active neurons or glia, as potential modulators through nonsynaptic actions. Similarly, circulating steroid hormones, steroids produced in the nervous system (Baulieu, 1998), locally released adenosine and other purines, prostaglandins and other arachidonic acid metabolites, and nitric oxide (NO) (Gally et al., 1990) might all now be regarded as modulators. Neuromediators Substances that participate in the elicitation of the postsynaptic response to a transmitter fall under this heading. The clearest examples of such effects are provided by the involvement of adenosine 3',5'-monophosphate (cyclic AMP), guanosine 3',5'-monophosphate (cyclic GMP), and inositol phosphates as second messengers at specific sites of synaptic transmission (seeChapters 6: Neurotransmission: the Autonomic and Somatic Motor Nervous Systems, 7: Muscarinic Receptor Agonists and Antagonists, 10: Catecholamines, Sympathomimetic Drugs, and Adrenergic Receptor Antagonists, and 11: 5-Hydroxytryptamine (Serotonin): Receptor Agonists and Antagonists). However, it is technically difficult to demonstrate in brain that a change in the concentration of cyclic nucleotides occurs prior to the generation of the synaptic potential and that this change in concentration is both necessary and sufficient for its generation. It is possible that changes in the concentration of second messengers can occur and enhance the generation of synaptic potentials. Activation of second messenger-dependent protein phosphorylation reactions can initiate a complex cascade of precise molecular events that regulate the properties of membrane and cytoplasmic proteins that are central to neuronal excitability (Greengard et al., 1999). These possibilities are particularly pertinent to the action of drugs that augment or reduce transmitter effects (see below). Neurotrophic Factors Neurotrophic factors are substances produced within the CNS by
neurons, astrocytes, microglia, or transiently invading peripheral
inflammatory or immune cells that assist neurons in their attempts to repair
damage. Seven categories of peptide factors have been recognized to operate
in this fashion (seeBlack, 1999; McKay et al., 1999, for recent
reviews): (1) the classic neurotrophins (nerve growth factor, brain-derived
neurotrophic factor, and the related neurotrophins); (2) the neuropoietic
factors, which have effects both in brain and in myeloid cells [e.g.,
cholinergic differentiation factor (also called leukemia inhibitory factor),
ciliary neurotrophic factor, and some interleukins]; (3) growth factor
peptides, such as epidermal growth factor, transforming growth factors |
Central Neurotransmitters
|
In examining the effects of drugs on the CNS with reference to the neurotransmitters for specific circuits, attention should be devoted to the general organizational principles of neurons. The view that synapses represent drug-modifiable control points within neuronal networks thus requires explicit delineation of the sites at which given neurotransmitters may operate and the degree of specificity with which such sites may be affected. One principle that underlies the following summaries of individual transmitter substances is the chemical-specificity hypothesis of Dale (1935), which holds that a given neuron releases the same transmitter substance at each of its synaptic terminals. In the face of growing indications that some neurons may contain more than one transmitter substance (Hkfelt, et al., 1995, 2000), Dale's hypothesis has been modified to indicate that a given neuron will secrete the same set of transmitters from all of its terminals. However, even this theory may require revision. For example, it is not clear whether or not a neuron that secretes a given peptide will process the precursor peptide to the same end product at all of its synaptic terminals. Table 121 provides an overview of the pharmacological properties of the transmitters in the CNS that have been studied extensively. Neurotransmitters are discussed below in terms of the groups of substances within given chemical categories: amino acids, amines, and neuropeptides. Other substances that may participate in central synaptic transmission include purines (such as adenosine and ATP (seeEdwards and Robertson, 1999; Moreau and Huber, 1999; Baraldi et al., 2000), nitric oxide (seeCork et al., 1998), and arachidonic acid derivatives (seeMechoulam et al., 1996; Piomelli, et al., 1998). Amino Acids The CNS contains uniquely high concentrations of certain amino acids,
notably glutamate and GABA; these amino acids are extremely potent in their
ability to alter neuronal discharge. Initially, physiologists were reluctant
to accept these simple substances as central neurotransmitters. Their
ubiquitous distribution within the brain and the consistent observation that
they produced prompt, powerful, and readily reversible but redundant effects
on every neuron tested seemed out of keeping with the extreme heterogeneity
of distribution and responsivity seen for other putative transmitters. The
dicarboxylic amino acids produced near-universal excitation, and the
monocarboxylic GABA GABA was identified as a unique chemical constituent of brain in 1950, but its potency as a CNS depressant was not immediately recognized. At the crustacean stretch receptor, GABA was identified as the only inhibitory amino acid found exclusively in crustacean inhibitory nerves and the inhibitory potency of extracts of these nerves was accounted for by their content of GABA. Release of GABA correlated with the frequency of nerve stimulation, and application of GABA and inhibitory nerve stimulation produced identical increases of Cl conductance in the muscle, fully satisfying the criteria for identification of GABA as the transmitter for this nerve (for further historical references, seeBloom, 1996). These same physiological and pharmacological properties later were found to be useful models in tests of a role for GABA in the mammalian CNS. Substantial data support the idea that GABA mediates the inhibitory actions of local interneurons in the brain and that GABA also may mediate presynaptic inhibition within the spinal cord. Presumptive GABA-ergic inhibitory synapses have been demonstrated most clearly between cerebellar Purkinje neurons and their targets in Deiter's nucleus; between small interneurons and the major output cells of the cerebellar cortex, olfactory bulb, cuneate nucleus, hippocampus, and lateral septal nucleus; and between the vestibular nucleus and the trochlear motoneurons. GABA also mediates inhibition within the cerebral cortex and between the caudate nucleus and the substantia nigra. GABA-ergic neurons and nerve terminals have been localized with immunocytochemical methods that visualize glutamic acid decarboxylase, the enzyme that catalyzes the synthesis of GABA from glutamic acid, or by in situ hybridization of the mRNAs for this protein. GABA-containing neurons frequently have been found to coexpress one or more neuropeptides. The most useful drugs for confirmation of GABA-ergic mediation have been bicuculline and picrotoxin; however, many convulsants whose actions previously were unexplained (including penicillin and pentylenetetrazol) also may act as relatively selective antagonists of the action of GABA (Macdonald et al., 1992; Macdonald and Olsen, 1994). Useful therapeutic effects have not yet been obtained through the use of agents that mimic GABA (such as muscimol), inhibit its active reuptake (such as 2,4-diaminobutyrate, nipecotic acid, and guvacine), or alter its turnover (such as aminooxyacetic acid). GABA is the major inhibitory neurotransmitter in the mammalian CNS. Its receptors have been divided into two main types. The more prominent GABA receptor subtype, the GABAA receptor, is a ligand-gated Cl ion channel, an 'ionotropic receptor' that is opened after release of GABA from presynaptic neurons. A second receptor, the GABAB receptor, is a member of the GPCR family, as noted above, and is coupled both to biochemical pathways and to regulation of ion channels, a class of receptor generally referred to as 'metabotropic' (Grifa et al., 1998; Billinton et al., 1999; Brauner-Osborne and Krogsgaard-Larsen, 1999). The GABAA receptor subunit proteins have been well characterized due to their high abundance and the receptor's role in almost every neuronal circuit. The receptor also has been extensively characterized in its role as the site of action of many neuroactive drugs (seeChapter 17: Hypnotics and Sedatives). Notable among these are benzodiazepines and barbiturates. It has been suggested recently that direct interactions occur between GABAA receptors and anesthetic steroids, volatile anesthetics, and alcohol (Macdonald, Twyman et al., 1992). Based on sequence homology to the first GABAA subunit cDNAs, more than 15 other subunits have been cloned. In addition to these subunits, which are products of separate genes, mRNA splice variants for several subunits have been described. The GABAA receptor, by analogy with the classical ionotropic nicotinic cholinergic receptor, may be either a pentameric or tetrameric protein in which the subunits assemble together around a central ion pore, a structural format typical for all ionotropic receptors. Abundant evidence has shown that there are multiple subtypes of GABAA receptors in the brain. The existence of subtypes was first suggested by pharmacological differences. It is now known that receptors composed of particular subunits have distinct pharmacological properties (Barnard et al., 1988; Olsen et al., 1990; Seeburg et al., 1990), but the true heterogeneity of GABAA-receptor subtypes has yet to be fully defined. Differences in anatomical distribution of subunits and differences in the time course of development of genes expressing each subunit suggest that there are important functional differences among the subtypes. The subunit composition of the major form of the GABAA
receptor contains at least three different subunits Glycine Many of the features described for the GABAA receptor family also are features of the inhibitory glycine receptor that is prominent in the brainstem and spinal cord. Multiple subunits have been cloned, and they can assemble into a variety of glycine-receptor subtypes (Grenningloh et al., 1987; Malosio et al., 1991). These pharmacological subtypes are detected in brain tissue with particular neuroanatomical and neurodevelopmental profiles. However, as with the GABAA receptor, the complete functional significance of the glycine receptor subtypes is not known. Glutamate and Aspartate Glutamate and aspartate are found in very high concentrations in brain, and both amino acids have extremely powerful excitatory effects on neurons in virtually every region of the CNS. Their widespread distribution tended to obscure their roles as transmitters, but there is now broad acceptance of the view that glutamate and possibly aspartate function as the principal fast ('classical') excitatory transmitters throughout the CNS (seeSeeburg, 1993; Cotman et al., 1995; Herrling, 1997). Furthermore, over the past decade, multiple subtypes of receptors for excitatory amino acids have been characterized pharmacologically, based on the relative potencies of synthetic agonists and the discovery of potent and selective antagonists (seeHerrling, 1997). Glutamate receptors, like those for GABA, are classified functionally either as ligand-gated ion channels ('ionotropic' receptors) or as 'metabotropic' (G proteincoupled) receptors. Neither the precise number of subunits that assemble to generate a functional glutamate receptor ion channel in vivo nor the topography of each subunit has been established unequivocally (Borges and Dingledine, 1998; Dingledine et al., 1999). The ligand-gated ion channels are further classified according to the
identity of agonists that selectively activate each receptor subtype. These
receptors include Multiple cDNAs encoding metabotropic receptors and subunits of NMDA, AMPA, and kainate receptors have been cloned in recent years (Borges and Dingledine, 1998; Dingledine et al., 1999). The diversity of gene expression and, consequently, of the protein structure of glutamate receptors also arises by alternative splicing and in some cases by single-base editing of mRNAs encoding the receptors or receptor subunits. Alternative splicing has been described for metabotropic receptors and for subunits of NMDA, AMPA, and kainate receptors (Hollmann and Heinemann, 1994). A remarkable form of endogenous molecular engineering occurs with some subunits of AMPA and kainate receptors in which the RNA sequence differs from the genomic sequence in a single codon of the receptor subunit and determines the extent of Ca2+ permeability of the receptor channel (Traynelis et al., 1995). This RNA-editing process alters the identity of a single amino acid (out of about 900 amino acids) that dictates whether or not the receptor channel gates Ca2+. The glutamate receptor genes seem to be unique families with only limited similarity to other ligand-gated channels such as the nicotinic acetylcholine receptor or, in the case of metabotropic receptors, to members of the GPCR superfamily. AMPA and kainate receptors mediate fast depolarization at most glutamatergic synapses in the brain and spinal cord. NMDA receptors also are involved in normal synaptic transmission, but activation of NMDA receptors is more closely associated with the induction of various forms of synaptic plasticity rather than with fast point-to-point signaling in the brain. AMPA or kainate receptors and NMDA receptors may be colocalized at many glutamatergic synapses. A well-characterized phenomenon that involves NMDA receptors is the induction of long-term potentiation (LTP). LTP refers to a prolonged (hours to days) increase in the size of a postsynaptic response to a presynaptic stimulus of given strength. Activation of NMDA receptors is obligatory for the induction of one type of LTP that occurs in the hippocampus (Bliss and Collingridge, 1993). NMDA receptors normally are blocked by Mg2+ at resting membrane potentials. Thus, activation of NMDA receptors requires not only binding of synaptically released glutamate but simultaneous depolarization of the postsynaptic membrane. This is achieved by activation of AMPA/kainate receptors at nearby synapses from inputs from different neurons. Thus, NMDA receptors may function as coincidence detectors, being activated only when there is simultaneous firing of two or more neurons. Interestingly, NMDA receptors also can induce long-term depression (LTD; the flip side of LTP) at CNS synapses (Malenka and Nicoll, 1998). It seems that the frequency and pattern of synaptic stimulation is what dictates whether a synapse undergoes LTP or LTD (seeMalenka and Nicoll, 1999). Glutamate Excitotoxicity The ability of high concentrations of glutamate to produce neuronal cell death has been known for more than three decades (Olney, 1969), but the mechanisms by which glutamate and selective, rigid agonists of its receptors produce this effect only recently have begun to be clarified. The cascade of events leading to neuronal death initially was thought to be triggered exclusively by excessive activation of NMDA or AMPA/kainate receptors, which allow significant influx of Ca2+ into the neurons. Such glutamate neurotoxicity was thought to underlie the damage that occurs after ischemia or hypoglycemia in the brain, during which a massive release and impaired reuptake of glutamate in the synapse would lead to excess stimulation of glutamate receptors and subsequent cell death. Although NMDA receptor antagonists can attenuate or block neuronal cell death induced by activation of these receptors (seeHerrling, 1997), even the most potent antagonists could not prevent all such damage. More recent studies (seeChoi and Koh, 1998; Lee et al., 1999; Zipfel et al., 1999) implicate both local depletion of Na+ and K+, as well as small but significant elevations of extracellular Zn2+ as factors that can activate both necrotic and proapoptotic cascades (Merry and Korsmeyer, 1997) leading to neuronal death. NMDA receptors also may be involved in the development of susceptibility to epileptic seizures and in the occurrence of seizure activity (Blumcke et al., 1995). Cases of Rasmussen's encephalitis, a childhood disease leading to intractable seizures and dementia, were found to correlate with levels of serum antibodies to a glutamate receptor subunit (Rogers et al., 1994). Because of the widespread distribution of glutamate receptors in the CNS, it is likely that these receptors ultimately will become the targets for diverse therapeutic interventions. For example, a role for disordered glutamatergic transmission in the etiology of chronic neurodegenerative diseases and in schizophrenia has been postulated (Farber et al., 1998; Olney et al., 1999). Acetylcholine After acetylcholine (ACh) was identified as the transmitter at neuromuscular and parasympathetic neuroeffector junctions, as well as at the major synapse of autonomic ganglia (seeChapter 6: Neurotransmission: The Autonomic and Somatic Motor Nervous Systems), the amine began to receive considerable attention as a potential central neurotransmitter. Based on its irregular distribution within the CNS and the observation that peripheral cholinergic drugs could produce marked behavioral effects after central administration, many investigators were willing to consider the possibility that ACh also might be a central neurotransmitter. In the late 1950s, Eccles and colleagues demonstrated that the recurrent excitation of spinal Renshaw neurons was sensitive to nicotinic cholinergic antagonists; these cells also were found to be cholinoceptive. Such observations were consistent with the chemical and functional specificity of Dale's hypothesis that all branches of a neuron released the same transmitter substance and, in this case, produced similar types of postsynaptic action (seeEccles, 1964). Although the ability of ACh to elicit neuronal discharge subsequently has been replicated on scores of CNS cells (seeShepherd, 1998), the spinal Renshaw cell remains the prototype for central nicotinic cholinergic synapses. Nevertheless, the search for selectively acting, central nicotinic drugs continues (Decker et al., 1997; Bannon et al., 1998). In most regions of the CNS, the effects of ACh, assessed either by iontophoresis or by radioligand receptordisplacement assays, appear to be generated by interaction with a mixture of nicotinic and muscarinic receptors. Several sets of presumptive cholinergic pathways have been proposed in addition to that of the motoneuron-Renshaw cell. By combination of immunocytochemistry of choline acetyltransferase (ChAT; the enzyme that synthesizes ACh) and ligand binding or in situ hybridization studies for the detection of neurons expressing subunits of nicotinic and muscarinic receptors, eight major clusters of ACh neurons and their pathways have been characterized (Mesulam, 1995). Four separate groups of cell bodies located in the basal forebrain, between the septum and the nucleus basalis of Meynert, send largely autonomous projections to the neocortex, hippocampal formation, and olfactory bulb. While rodent brains exhibit cholinergic neurons that are intrinsic to the neocortex, these neurons are not found in primate brain. Two collections of cholinergic neurons in the upper pons provide the major cholinergic innervation of thalamus and striatum, while medullary cholinergic neurons provide the cholinergic innervation of midbrain and brainstem regions. The intense cholinergic projections to neocortex and hippocampal formation will atrophy if these neurons are deprived of the trophic growth factors provided to them by retrograde axonal transport from their target neurons (Sofroniew et al., 1993). This occurs in Alzheimer's disease when these target neurons are diseased (seeChapter 22: Treatment of Central Nervous System Degenerative Disorders) and has driven therapeutic efforts to restore residual cholinergic signaling. Catecholamines The brain contains separate neuronal systems that utilize three different catecholaminesdopamine, norepinephrine, and epinephrine. Each system is anatomically distinct and serves separate, but similar, functional roles within their fields of innervation. Much of the original mapping was performed in rodent brains (Hkfelt et al., 1976, 1977), but recent studies have extended these maps into primates (Foote, 1997; Lewis, 1997). Dopamine Although dopamine originally was regarded only as a precursor of norepinephrine, assays of distinct regions of the CNS eventually revealed that the distributions of dopamine and norepinephrine are markedly different. In fact, more than half the CNS content of catecholamine is dopamine, and extremely large amounts are found in the basal ganglia (especially the caudate nucleus), the nucleus accumbens, the olfactory tubercle, the central nucleus of the amygdala, the median eminence, and restricted fields of the frontal cortex. The anatomical connections of the dopamine-containing neurons are known with some precision. Dopaminergic neurons fall into three major morphological classes: (1) ultrashort neurons within the amacrine cells of the retina and periglomerular cells of the olfactory bulb; (2) intermediate-length neurons within the tuberobasal ventral hypothalamus that innervate the median eminence and intermediate lobe of the pituitary, connect the dorsal and posterior hypothalamus with the lateral septal nuclei, and extend caudally to the dorsal motor nucleus of the vagus, the nucleus of the solitary tract, and the periaqueductal gray matter; and (3) long projections between the major dopamine-containing nuclei in the substantia nigra and ventral tegmentum and their targets in the striatum, in the limbic zones of the cerebral cortex, and in other major regions of the limbic system except the hippocampus (seeHkfelt, et al., 1976, 1977). At the cellular level, the actions of dopamine depend on receptor subtype expression and the contingent convergent actions of other transmitters to the same target neurons. Although initial pharmacological studies discriminated between two subtypes of dopamine receptors, D1 (by which dopamine activates adenylyl cyclase) and D2 (by which dopamine inhibits adenylyl cyclase), subsequent cloning studies identified at least five genes encoding subtypes of dopamine receptors. Nevertheless, the two major categories, D1-like or D2-like, persist. The D1-like receptors include the D1 and the D5 receptors, whereas the D2-like receptors include the two isoforms of the D2 receptor, differing in the length of their predicted third cytoplasmic loop, dubbed D2short (D2S) and D2long (D2L), the D3, and the D4 receptors (seeGrandy and Civelli, 1992; Gingrich and Caron, 1993; Civelli, 1994). The D1 and D5 receptors activate adenylyl cyclase. The D2 receptors couple to multiple effector systems, including the inhibition of adenylyl cyclase activity, suppression of Ca2+ currents, and activation of K+ currents. The effector systems to which the D3 and D4 receptors couple have not been unequivocally defined (Sokoloff and Schwartz, 1995; Schwartz et al., 1998). D2 dopamine receptors have been implicated in the pathophysiology of schizophrenia and Parkinson's disease (seeChapters 20: Drugs and the Treatment of Psychiatric Disorders: Psychosis and Mania and 22: Treatment of Central Nervous System Degenerative Disorders). Norepinephrine There are relatively large amounts of norepinephrine within the hypothalamus and in certain zones of the limbic system, such as the central nucleus of the amygdala and the dentate gyrus of the hippocampus. However, this catecholamine also is present in significant, although lower, amounts, in most brain regions. Detailed mapping studies indicate that most noradrenergic neurons arise either in the locus ceruleus of the pons or in neurons of the lateral tegmental portion of the reticular formation. From these neurons, multiple branched axons innervate specific target cells in a large number of cortical, subcortical, and spinomedullary fields (Hkfelt, et al., 1976, 1977; Foote and Aston-Jones, 1995; Foote, 1997). Although norepinephrine has been firmly established as the transmitter
at synapses between presumptive noradrenergic pathways and a wide variety of
target neurons, a number of features of the mode of action of this biogenic
amine have complicated the acquisition of convincing evidence. In large part,
these problems reflect its 'nonclassical' electrophysiological
synaptic actions, which result in 'state-dependent' or
'enabling' effects. In some instances, the pharmacological
properties of such synapses have been complex, with evidence for mediation by
both As in the periphery, three families of adrenergic receptors have been
described in the CNS (i.e., The Epinephrine Neurons in the CNS that contain epinephrine were recognized only after the development of sensitive enzymatic assays for phenylethanolamine-N-methyltransferase and immunocytochemical staining techniques for the enzyme (seeHkfelt et al., 1976 and references therein). Epinephrine-containing neurons are found in the medullary reticular formation and make restricted connections to a few pontine and diencephalic nuclei, eventually coursing as far rostrally as the paraventricular nucleus of the dorsal midline thalamus. Their physiological properties have not been identified. 5-Hydroxytryptamine Following the chemical determination that a biogenic substance found both in serum ('serotonin') and in gut ('enteramine') was 5-hydroxytryptamine (5-HT), assays for this substance revealed its presence in brain (seeChapter 11: 5-Hydroxytryptamine (Serotonin): Receptor Agonists and Antagonists). Since that time, studies of 5-HT have had a pivotal role in advancing our understanding of the neuropharmacology of the CNS. Various cytochemical methods have been used to trace the central anatomy of 5-HT-containing neurons in several species. Tryptaminergic neurons are found in nine nuclei lying in or adjacent to the midline (raphe) regions of the pons and upper brainstem, corresponding to well-defined nuclear ensembles (Steinbusch and Mulder, 1984). The rostral raphe nuclei innervate forebrain regions, while the caudal raphe nuclei project within the brainstem and spinal cord with some overlaps. The median raphe nucleus makes a major contribution to the innervation of the limbic system, and the dorsal raphe nucleus makes a similar contribution to cortical regions and the neostriatum. In the mammalian CNS, cells receiving cytochemically demonstrable tryptaminergic input, such as the suprachiasmatic nucleus, ventrolateral geniculate body, amygdala, and hippocampus, exhibit a uniform and dense investment of reactive terminals. Molecular biological approaches have led to identification of 14
distinct mammalian 5-HT-receptor subtypes. These subtypes exhibit
characteristic ligand-binding profiles, couple to different intracellular
signaling systems, exhibit subtype-specific distributions within the CNS, and
mediate distinct behavioral effects of 5-HT. Present terminology has grouped
the known 5-HT receptor subtypes into multiple classes: the 5-HT1and
5-HT2classes of receptor are both G proteincoupled receptors with
a seven-transmembrane-spanning-domain motif and include multiple isoforms
within each class, while the 5-HT3 receptor is a ligand-gated ion
channel with structural similarity to the The 5-HT1 receptor subset is composed of at least five intronless receptor subtypes (5-HT1A, 5-HT1B, 5-HT1D, 5-HT1E, 5-HT1F) that are linked to inhibition of adenylyl cyclase activity or to regulation of K+ or Ca2+ channels. The 5-HT1A receptors are abundantly expressed on 5-HT neurons of the dorsal raphe nucleus, where they are thought to be involved in temperature regulation. They also are found in regions of the CNS associated with mood and anxiety such as the hippocampus and amygdala. Activation of 5-HT1A receptors leads to opening of an inwardly rectifying K+ conductance, which leads to hyperpolarization and neuronal inhibition. These receptors can be activated by the drugs buspirone and ipsapirone, which are used to treat anxiety and panic disorders (seeAghajanian, 1995). 5-HT1D receptors are potently activated by the drug sumatriptan, which is currently prescribed for acute management of migraine headaches. Three receptor subtypes constitute the 5-HT2 receptor class: 5-HT2A, 5-HT2B, and 5-HT2C. In contrast to 5-HT1 receptors, these 5-HT2 receptors contain introns and all are linked to activation of phospholipase C. Based on ligand binding and mRNA in situ hybridization patterns, 5-HT2A receptors are enriched in forebrain regions such as neocortex and olfactory tubercle, as well as in several nuclei arising from the brainstem. On facial motoneurons, 5-HT enhances excitability by two mechanisms: (1) slow closure of resting K+ conductances, increasing membrane resistance; and (2) a more potent, ritanserin-antagonizable opening of a voltage-sensitive K+ conductance that is activated by hyperpolarization (Aghajanian, 1995). In piriform cortex, Aghajanian and colleagues have observed an indirect inhibition of pyramidal neurons through activation of local-circuit, GABA-mediated inhibitory interneurons, an effect that is blocked by ritanserin. In the cerebral cortex, 5-HT2A-receptor agonists (but not 5-HT itself) also produce neuronal inhibition, but other effects reported may be the result of coexpression of multiple 5-HT-receptor subtypes on the same neuron. The 5-HT2C receptor, which is very similar in sequence and pharmacology to the 5-HT2A receptor, is expressed abundantly in the choroid plexus where it regulates transferrin and cerebrospinal fluid production (seeChapter 11: 5-Hydroxytryptamine (Serotonin): Receptor Agonists and Antagonists). Receptors of the 5-HT3class first were recognized in the peripheral autonomic system. They also are expressed in brain within the area postrema and nucleus tractus solitarius, where they couple to potent depolarizing responses that show rapid desensitization to continued 5-HT exposure. The 5-HT3 receptor leads to enhanced Na+and K+ currents but does not seem to affect Ca2+ permeability. At the behavioral level, actions of 5-HT at central 5-HT3 receptors can lead to emesis and antinociceptive actions; 5-HT3-receptor antagonists such as ondansetron are beneficial in the management of chemotherapy-induced emesis (seeChapter 38: Prokinetic Agents, Antiemetics, and Agents Used in Irritable Bowel Syndrome). The hallucinogen LSD is among the most interesting of the compounds that interact with 5-HT, primarily through 5-HT2 receptors. In iontophoretic tests, LSD and 5-HT are both potent inhibitors of the firing of raphe (5-HT) neurons, whereas LSD and other hallucinogens are far more potent excitants on facial motoneurons that receive innervation from the raphe. The inhibitory effect of LSD on raphe neurons offers a plausible explanation for the drug's hallucinogenic effects, namely, that these effects result from depression of activity in a system that tonically inhibits visual and other sensory inputs. However, typical LSD-induced behavior is still seen in animals with destroyed raphe nuclei or after blockade of the synthesis of 5-HT by p-chlorophenylalanine. Other evidence against this explanation of LSD-induced hallucinations is potentiation of the effects of LSD by administration of the precursor of 5-HT, 5-hydroxytryptophan. More precise definition of the various functional roles of tryptaminergic pathways in the CNS awaits the results of studies using more specific agents, whose number is steadily increasing (seeAghajanian and Marek, 1999). Histamine For many years, histamine and antihistamines that are active in the periphery have been known to produce significant effects on animal behavior. Relatively recently, however, evidence has accumulated to suggest that histamine also might be a central neurotransmitter. Biochemical detection of histamine synthesis by neurons, as well as direct cytochemical localization of these neurons, has established the existence of a histaminergic system in the CNS. Most of these neurons are located in the ventral posterior hypothalamus; they give rise to long ascending and descending tracts to the entire CNS that are typical of the patterns characteristic of other aminergic systems. Based on the presumptive central effects of histamine antagonists, the histaminergic system is thought to function in the regulation of arousal, body temperature, and vascular dynamics. Three subtypes of histamine receptors have been described. H1 receptors, the most prominent, may be located on glia and vessels as well as on neurons and may act to mobilize Ca2+ in receptive cells. H2 receptors are linked to the activation of adenylyl cyclase, perhaps in concert with H1 receptors in certain circumstances. H3 receptors, which have the greatest sensitivity to histamine, are localized much more selectively in basal ganglia and olfactory regions in the rat, but consequences of their activation remain unresolved. Unlike the monoamines and amino acid transmitters, there does not appear to be an active reuptake process for histamine after its release. In addition, no direct evidence has been obtained for release of histamine from neurons either in vivo or in vitro(seeSchwartz et al., 1995, for additional recent references). Peptides The discovery during the 1980s of numerous novel peptides in the CNS, each capable of regulating one or another aspect of neural function, produced considerable excitement and an imposing catalog of entities (seeHkfelt et al., 1995; Darlison and Richter, 1999). In addition, certain peptides previously thought to be restricted to the gut or to endocrine glands also have been found in the CNS. Relatively detailed neuronal maps are available that show immunoreactivity to peptide-specific antisera. While some CNS peptides may function on their own, most are now thought to act mainly in concert with coexisting transmitters, both amines and amino acids. Some neurons may contain more than two possible transmitters (seeHkfelt et al., 1995), and they can be independently regulated. At this time at least three approaches appear to have utility in attempting to grasp the continuously enlarging peptidergic systems of neurons. Organization by Peptide Families Because of significant homology in amino acid sequences, families of related molecules can be defined as either ancestral or concurrent. The ancestral relationship is illustrated by peptides such as the tachykinin/ substance P or the vasotocin (vasopressin/oxytocin) family, in which species differences can be correlated with modest variations in peptide structure. The concurrent relationship is best exemplified by the endorphins and by the glucagon-secretin family. In the endorphin 'superfamily,' three major systems of endorphin peptides (proopiomelanocortin, proenkephalin, and prodynorphin) exist in independent neuronal circuits (seeAkil et al., 1998 for recent review). These natural opioid peptides arise from independent, but homologous, genes. The peptides all share some actions at receptors once classed generally as 'opioid' but now are undergoing progressive refinement (seeChapter 23: Opioid Analgesics). In the glucagon family, multiple and somewhat homologous peptides are found simultaneously in different cells of the same organism but in separate organ systems: glucagon and vasoactive intestinal polypeptide (VIP) in pancreatic islets; secretin in duodenal mucosa; VIP and related peptides in enteric, autonomic, and central neurons (seeMagistretti et al., 1998); and growth hormone-releasing factor in central neurons only (Guillemin et al., 1984). The general metabolic effects produced by this family can be viewed as leading to increased blood glucose. To some degree, ancestral and concurrent relationships are not mutually exclusive. For example, multiple members of the tachykinin/substance P family within mammalian brains and intestines may account for the apparent existence of subsets of receptors for these peptides (Vanden Broeck et al., 1999). The mammalian terminus of the vasotocin family shows two concurrent products as well, vasopressin and oxytocin, each having evolved to perform separate functions once executed by single vasotocin-related peptides in lower phyla. Organization by Anatomic Pattern Some peptide systems follow rather consistent anatomical organizations. Thus, the hypothalamic peptides oxytocin, vasopressin, proopiomelanocortin, gonadotropin-releasing hormone, and growth hormonereleasing hormone all tend to be synthesized by single large clusters of neurons that give off multibranched axons to several distant targets. Others, such as systems that contain somatostatin, cholecystokinin, and enkephalin, can have many forms, with patterns varying from moderately long, hierarchical connections to short-axon, local-circuit neurons that are widely distributed throughout the brain (seeHkfelt et al., 2000). Organization by Function Since almost all peptides initially were identified on the basis of bioassays, their names reflect these biologically assayed functions (e.g., thyrotropin-releasing hormone, vasoactive intestinal polypeptide). These names become trivial if more ubiquitous distributions and additional functions are discovered. Some general integrative role might be hypothesized for widely separated neurons (and other cells) that make the same peptide. However, a more parsimonious view would be that each peptide has unique messenger roles at the cellular level and that these are used repeatedly in functionally similar pathways within large systems that differ in their overall functions. The cloning of the major members of the opioid-peptide receptors revealed unexpected and as yet unexplained conservation of sequences with receptors for somatostatin, angiotensin, and other peptides (seeUhl et al., 1994). Comparison with Other Transmitters Peptides differ in several important respects from the monoamine and amino acid transmitters considered earlier. Synthesis of a peptide is performed in the rough endoplasmic reticulum, where mRNA for the propeptide can be translated into an amino-acid sequence. The propeptide is cleaved (processed) to the form that is secreted as the secretory vesicles are transported from the perinuclear cytoplasm to the nerve terminals. Further, no active reuptake mechanisms for peptides have been described (but seeHonor et al., 1999, for possible exception). This increases the dependency of peptidergic nerve terminals on distant sites of synthesis. Perhaps most importantly, linear chains of amino acids can assume many conformations at their receptors, making it difficult to define the sequences and their steric relationships that are critical for activity. Until recently, it has been difficult to develop nonpeptidic, synthetic agonists or antagonists that will interact with specific receptors for peptides. However, such drugs are now being developed (for cholecystokinin CCK1and CCK2 receptors, for neurotensin receptors, and for corticotropin-releasing-hormone receptors), and some (against substance P NK-1 receptors) have entered clinical trials (seeRupniak and Kramer, 1999; Hkfelt et al., 2000). Nature also has had limited success in this regard, since only one plant alkaloid, morphine, has been found to act selectively at peptidergic synapses. Fortunately for pharmacologists, morphine was discovered before the endorphins, or rigid molecules capable of acting at peptide receptors might have been deemed impossible to develop. Other Regulatory Substances In addition to these major families of neurotransmitters, other endogenous substances also may participate in the regulated flow of signals between neurons, but in sequences of events that differ somewhat from the conventional concepts of neurotransmitter function. These substances have significant potential importance as regulatory factors and as targets for future drug development. Purines In addition to their roles as essential biochemical anabolites, adenosine monophosphate, adenosine triphosphate, and free adenosine have gained attention as independent, neuronal signaling molecules in their own right (seeMoreau and Huber, 1999; Williams et al., 1999; Baraldi et al., 2000). Two large families of purinergic receptors have been characterized. Those in the P1 class are GPCRs. These have been further divided into four subtypes (A1A4) based on agonist actions of adenosine; A1 and A2 adenosine receptors are antagonized by xanthines, whereas A3 and A4 adenosine receptors are not. A1 adenosine receptors have been associated with inhibition of adenylyl cyclase, activation of K+ currents, activation of phospholipase C in some circumstances, and ion-channel regulation, while A2 receptors activate adenylyl cyclase. The P2 class of purine receptors refers to the receptors for ATP and related triphosphate nucleotides such as UTP. The P2X subtype of receptor is a ligand-gated ion channel, while the P2Y subtype is a GPCR. Adenosine can act presynaptically throughout the cortex and hippocampal formation to inhibit the release of amine and amino acid transmitters. ATP-regulated responses have been linked pharmacologically to a variety of supracellular functions including anxiety, stroke, and epilepsy (Williams, 1995). Diffusible Mediators Certain potent agents shown to be active under pharmacological conditions and inferred to be physiological regulators in systems throughout the body recently have been examined for their roles within the central nervous system. Arachidonic acid, normally stored within the cell membrane as a glycerol ester, can be liberated during phospholipid hydrolysis (by pathways involving phospholipases A2, C, and D). Phospholipases are activated by a variety of receptors. Arachidonic acid can be converted to highly reactive regulators by three major enzymatic pathways (seeChapter 26: Lipid-Derived Autacoids: Eicosanoids and Platelet-Activating Factor): cyclooxygenases (leading to prostaglandins and thromboxanes), lipoxygenases (leading to the leukotrienes and other transient catabolites of eicosatetraenoic acid), and cytochrome P450 (which is inducible although expressed at low levels in brain). These arachidonic acid metabolites have been implicated as diffusible modulators in the CNS, particularly for long-term potentiation and other forms of plasticity (Mechoulam et al., 1996; Piomelli et al., 1998). Nitric oxide has been recognized as an important regulator of vascular and inflammatory mediation for more than a decade, but came into focus with respect to roles in the CNS after successful efforts to characterize brain nitric oxide synthases (NOS; seeSnyder and Dawson, 1995). Molecular cloning studies have now revealed at least four isoforms of this biosynthetic enzyme in the brain, a constitutive form present in some neurons, capillary endothelial cells, and macrophages, as well as inducible forms of the enzyme. The availability of potent activators and inhibitors of NOS has led to reports of the presumptive involvement of nitric oxide in a host of phenomena in the brain including long-term potentiation, guanylyl cyclase activation, neurotransmitter release and reuptake, and enhancement of glutamate (NMDA)-mediated neurotoxicity. Subsequently, rational analysis based on proposed mechanisms of NO action through binding to the iron in the active site of target enzymes led to the idea that carbon monoxide may be a second gaseous, labile, diffusible intercellular regulator, at least in the regulation of guanylyl cyclase in neurons in vitro. Cytokines The term cytokines encompasses a large and diverse family of polypeptide regulators, produced widely throughout the body by cells of diverse embryological origin. In general, these regulators have multiple functions attributed to effects under controlled conditions in vitro. In vivo, the effects of cytokines are known to be further regulated by the conditions imposed by other cytokines, interacting as a network with variable effects leading to synergistic, additive, or opposing actions. Within the cytokines, tissue-produced peptidic factors termed chemokines serve to attract cells of the immune and inflammatory lines into interstitial spaces. These special cytokines have received much attention as potential regulators in nervous system inflammation (as in early stages of dementia, following infection with human immunodeficiency virus; seeAsensio et al., 1999; Mennicken et al., 1999) and during recovery from traumatic injury. The more conventional neuronal and glial-derived growth-enhancing and growth-retardant factors were mentioned above. The fact that, under some pathophysiological conditions, neurons and astrocytes may be induced to express cytokines or other growth factors further blurs the dividing line between neurons and glia. |
Actions of Drugs in the CNS
|
Specificity and Nonspecificity of CNS Drug Actions The effect of a drug is considered to be specific when it affects an identifiable molecular mechanism unique to target cells that bear receptors for that drug. Conversely, a drug is regarded as nonspecific when it produces effects on many different target cells and acts by diverse molecular mechanisms. This distinction is often a property of the dose-response relationship of the drug and the cell or mechanisms under scrutiny (seeChapter 3: Principles of Therapeutics). Even a drug that is highly specific when tested at a low concentration may exhibit nonspecific actions at substantially higher doses. Conversely, even generally acting drugs may not act equally on all levels of the CNS. For example, sedatives, hypnotics, and general anesthetics would have very limited utility if central neurons that control the respiratory and cardiovascular systems were sensitive to their actions. Drugs with specific actions may produce nonspecific effects if the dose and route of administration produce high tissue concentrations. Drugs whose mechanisms currently appear to be primarily general or nonspecific are classed according to whether they produce behavioral depression or stimulation. Specifically acting CNS drugs can be classed more definitively according to their locus of action or specific therapeutic usefulness. It must be remembered that the absence of overt behavioral effects does not rule out the existence of important central actions for a given drug. For example, the impact of muscarinic cholinergic antagonists on the behavior of normal animals may be subtle, but these agents are used extensively in the treatment of movement disorders and motion sickness (seeChapter 7: Muscarinic Receptor Agonists and Antagonists). General (Nonspecific) CNS Depressants This category includes the anesthetic gases and vapors, the aliphatic alcohols, and some hypnotic-sedative drugs. These agents share the ability to depress excitable tissue at all levels of the CNS, leading to a decrease in the amount of transmitter released by the nerve impulse, as well as to general depression of postsynaptic responsiveness and ion movement. At subanesthetic concentrations, these agents (e.g., ethanol) can exert relatively specific effects on certain groups of neurons, which may account for differences in their behavioral effects, especially the propensity to produce dependence (Koob and Le Moal, 1997; Koob et al., 1998; seealsoChapters 14: General Anesthetics, 17: Hypnotics and Sedatives, 18: Ethanol, and 24: Drug Addiction and Drug Abuse). General (Nonspecific) CNS Stimulants The drugs in this category include pentylenetetrazol and related agents that are capable of powerful excitation of the CNS and the methylxanthines, which have a much weaker stimulant action. Stimulation may be accomplished by one of two general mechanisms: (1) by blockade of inhibition or (2) by direct neuronal excitation (which may involve increased transmitter release, more prolonged transmitter action, labilization of the postsynaptic membrane, or decreased synaptic recovery time). Drugs That Selectively Modify CNS Function The agents in this group may cause either depression or excitation. In some instances, a drug may produce both effects simultaneously on different systems. Some agents in this category have little effect on the level of excitability in doses that are used therapeutically. The principal classes of these CNS drugs include the following: anticonvulsants, antiparkinsonism drugs, opioid and nonopioid analgesics, appetite suppressants, antiemetics, analgesic-antipyretics, certain stimulants, antidepressants, antimanic agents, antipsychotic agents, sedatives, and hypnotics. Although selectivity of action may be remarkable, a drug usually affects several CNS functions to varying degrees. When only one constellation of effects is wanted in a therapeutic situation, the remaining effects of the drug are regarded as limitations in selectivity (i.e., unwanted side effects). The specificity of a drug's action frequently is overestimated. This is partly due to the fact that the drug is identified with the effect that is implied by the class name. General Characteristics of CNS Drugs Combinations of centrally acting drugs frequently are administered to therapeutic advantage (e.g., an anticholinergic drug and levodopa for Parkinson's disease). However, other combinations of drugs may be detrimental because of potentially dangerous additive or mutually antagonistic effects. The effect of a CNS drug is additive with the physiological state and with the effects of other depressant and stimulant drugs. For example, anesthetics are less effective in a hyperexcitable subject than in a normal patient; the converse is true with respect to the effects of stimulants. In general, the depressant effects of drugs from all categories are additive (e.g., the fatal combination of barbiturates or benzodiazepines with ethanol), as are the effects of stimulants. Therefore, respiration depressed by morphine is further impaired by depressant drugs, while stimulant drugs can augment the excitatory effects of morphine to produce vomiting and convulsions. Antagonism between depressants and stimulants is variable. Some instances of true pharmacological antagonism among CNS drugs are known; for example, opioid antagonists are very selective in blocking the effects of opioid analgesics. However, the antagonism exhibited between two CNS drugs is usually physiological in nature. Thus, an individual who has received one drug cannot be returned entirely to normal by another. The selective effects of drugs on specific neurotransmitter systems may be additive or competitive. This potential for drug interaction must be considered whenever such drugs are administered concurrently. To minimize such interactions, a drug-free period may be required when modifying therapy. An excitatory effect is commonly observed with low concentrations of certain depressant drugs due either to depression of inhibitory systems or to a transient increase in the release of excitatory transmitters. Examples are the 'stage of excitement' during induction of general anesthesia and the 'stimulant' effects of alcohol. The excitatory phase occurs only with low concentrations of the depressant; uniform depression ensues with increasing drug concentration. The excitatory effects can be minimized, when appropriate, by pretreatment with a depressant drug that is devoid of such effects (e.g., benzodiazepines in preanesthetic medication). Acute, excessive stimulation of the cerebrospinal axis normally is followed by depression, which is in part a consequence of neuronal fatigue and exhaustion of stores of transmitters. Postictal depression is additive with the effects of depressant drugs. Acute, drug-induced depression is not, as a rule, followed by stimulation. However, chronic drug-induced sedation or depression may be followed by prolonged hyperexcitability upon abrupt withdrawal of the medication (barbiturates, alcohol). This type of hyperexcitability can be controlled effectively by the same or another depressant drug (seeChapters 17: Hypnotics and Sedatives and 18: Ethanol). Organization of CNSDrug Interactions The structural and functional properties of neurons provide a means to specify the possible sites at which drugs could interact specifically or generally in the CNS (seeFigure 121). In this scheme, drugs that affect neuronal energy metabolism, membrane integrity, or transmembrane ionic equilibria would be generally acting compounds. Similarly general in action would be drugs that affect the two-way intracellular transport systems (e.g., colchicine). These general effects still can exhibit different dose-response or time-response relationships based, for example, on such neuronal properties as rate of firing, dependence of discharge on external stimuli or internal pacemakers, resting ionic fluxes, or axon length. In contrast, when drug actions can be related to specific aspects of the metabolism, release, or function of a neurotransmitter, the site, specificity, and mechanism of action of a drug can be defined by systematic studies of dose-response and time-response relationships. From such data the most sensitive, rapid, or persistent neuronal event can be identified. Transmitter-dependent actions of drugs can be organized conveniently into presynaptic and postsynaptic categories. The presynaptic category includes all of the events in the perikaryon and nerve terminal that regulate transmitter synthesis (including the acquisition of adequate substrates and cofactors), storage, release, reuptake, and catabolism. Transmitter concentrations can be lowered by blockade of synthesis, storage, or both. The amount of transmitter released per impulse is generally stable but also can be regulated. The effective concentration of transmitter may be increased by inhibition of reuptake or by blockade of catabolic enzymes. The transmitter that is released at a synapse also can exert actions on the terminal from which it was released by interacting with receptors at these sites (termed autoreceptors;see above). Activation of presynaptic autoreceptors can slow the rate of discharge of transmitter and thereby provide a feedback mechanism that controls the concentration of transmitter in the synaptic cleft. The postsynaptic category includes all the events that follow release of the transmitter in the vicinity of the postsynaptic receptor in particular, the molecular mechanisms by which occupancy of the receptor by the transmitter results in changes in the properties of the membrane of the postsynaptic cell (shifts in membrane potential) as well as more enduring biochemical actions (changes in intracellular cyclic nucleotides, protein kinase activity, and related substrate proteins). Direct postsynaptic effects of drugs generally require relatively high affinity for the receptors or resistance to metabolic degradation. Each of these presynaptic or postsynaptic actions is potentially highly specific and can be envisioned as being restricted to a single, chemically defined subset of CNS cells. Convergence, Synergism, and Antagonism Result from Transmitter Interactions A hallmark of modern neuropharmacology is the capacity to clone receptor or receptor-subunit cDNAs and to determine their properties by expression in cells that do not normally express the receptor or subunit being studied. The simplicity of in vitro models of this type may divert one's attention from the fact that, in the intact CNS, a given neurotransmitter may interact simultaneously with all of the various isoforms of its receptor on neurons that are also under the influence of multiple other afferent pathways and their transmitters. Thus, attempts to predict the behavioral or therapeutic consequences of drugs designed to elicit precise and restricted receptor actions may fail due to differences under normal as compared to diseased conditions, and as a consequence of the complexity of the interactions possible. |
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 3255
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved